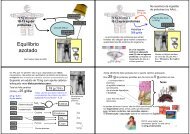
Ciclo de Krebs (ou do citrato)
Ciclo de Krebs (ou do citrato)
Ciclo de Krebs (ou do citrato)

Create successful ePaper yourself
Turn your PDF publications into a flip-book with our unique Google optimized e-Paper software.
<strong>Ciclo</strong> <strong>de</strong> <strong>Krebs</strong> (<strong>ou</strong> <strong>do</strong> <strong>citrato</strong>)<br />
No ciclo <strong>de</strong> <strong>Krebs</strong> (<strong>ou</strong> <strong>do</strong><br />
áci<strong>do</strong> cítrico) o grupo<br />
acetilo da acetil-CoA é<br />
oxida<strong>do</strong> a CO 2<br />
pelo NAD +<br />
e pelo FAD.<br />
Tal como<br />
a <strong>de</strong>sidrogénase <strong>do</strong> piruvato<br />
também todas as enzimas <strong>do</strong><br />
ciclo <strong>de</strong> <strong>Krebs</strong> estão <strong>de</strong>ntro da<br />
mitocôndria.<br />
Ao contrário <strong>do</strong> que acontece<br />
no citoplasma, não há <strong>de</strong>ntro<br />
da mitocôndria uma<br />
<strong>de</strong>sidrogénase láctica:<br />
a re-oxidação <strong>do</strong> NADH e<br />
FADH 2<br />
forma<strong>do</strong>s <strong>de</strong>pen<strong>de</strong> da<br />
activida<strong>de</strong> da ca<strong>de</strong>ia<br />
respiratória e <strong>do</strong> O 2<br />
.<br />
Rui Fontes<br />
1<br />
2<br />
Não existe ciclo <strong>de</strong> <strong>Krebs</strong> anaeróbio...<br />
1- O NADH e o FADH 2<br />
são o combustível <strong>do</strong>s<br />
complexos enzímicos da ca<strong>de</strong>ia respiratória que usam<br />
a energia libertada no processo oxidativo para bombear<br />
protões (contra gradiente) para fora da mitocôndria.<br />
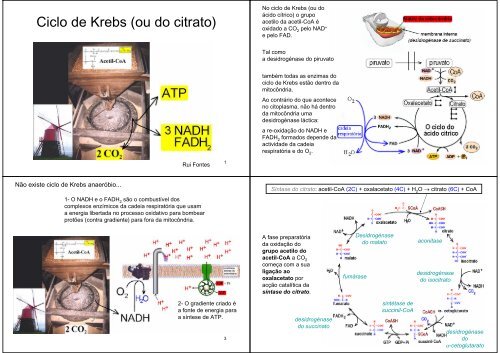
Síntase <strong>do</strong> <strong>citrato</strong>: acetil-CoA (2C) + oxalacetato (4C) + H 2<br />
O → <strong>citrato</strong> (6C) + CoA<br />
A fase preparatória<br />
da oxidação <strong>do</strong><br />
grupo acetilo <strong>do</strong><br />
acetil-CoA a CO 2<br />
começa com a sua<br />
ligação ao<br />
oxalacetato por<br />
acção catalítica da<br />
síntase <strong>do</strong> <strong>citrato</strong>.<br />
fumárase<br />
Desidrogénase<br />
<strong>do</strong> malato<br />
aconitase<br />
<strong>de</strong>sidrogénase<br />
<strong>do</strong> iso<strong>citrato</strong><br />
2- O gradiente cria<strong>do</strong> é<br />
a fonte <strong>de</strong> energia para<br />
a síntese <strong>de</strong> ATP.<br />
3<br />
<strong>de</strong>sidrogénase<br />
<strong>do</strong> succinato<br />
sintétase <strong>de</strong><br />
succinil-CoA<br />
<strong>de</strong>sidrogénase<br />
<strong>do</strong> 4<br />
α-cetoglutarato
Os passos oxidativos em<br />
que ocorre a redução <strong>do</strong><br />
NAD + e <strong>do</strong> FAD são os<br />
catalisa<strong>do</strong>s por 3<br />
<strong>de</strong>sidrogénases<br />
<strong>de</strong>pen<strong>de</strong>ntes <strong>do</strong> NAD +<br />
No passo catalisa<strong>do</strong> pela sintétase <strong>de</strong> succinil-CoA<br />
(succinil-CoA + GDP + Pi → succinato + GTP)<br />
forma-se GTP (“fosforilação ao nível <strong>do</strong> substrato”)<br />
... que por acção catalítica da cínase <strong>de</strong> nucleosí<strong>de</strong>os difosfatos (GTP + ADP → ATP<br />
+ GDP) permite a formação <strong>de</strong> ATP.<br />
(1) <strong>de</strong>sidrogénase <strong>do</strong><br />
iso<strong>citrato</strong>,<br />
(2) <strong>de</strong>sidrogénase <strong>do</strong><br />
α-cetoglutarato,<br />
(3) <strong>de</strong>sidrogénase <strong>do</strong><br />
malato<br />
e uma <strong>ou</strong>tra que tem<br />
como grupo prostético o<br />
FAD<br />
(4) <strong>de</strong>sidrogénase <strong>do</strong><br />
succinato<br />
Oxidações <strong>de</strong>scarboxílativas:<br />
as <strong>de</strong>scarboxilações (e consequente libertação <strong>de</strong> CO 2<br />
) ocorrem durante as acções 5<br />
catalíticas da <strong>de</strong>sidrogénase <strong>do</strong> iso<strong>citrato</strong> e da <strong>de</strong>sidrogénase <strong>do</strong> α-cetoglutarato.<br />
No “último” passo <strong>do</strong> ciclo <strong>de</strong><br />
<strong>Krebs</strong> ocorre a regeneração <strong>do</strong><br />
oxalacetato (acção catalítica da<br />
<strong>de</strong>sidrogénase <strong>do</strong> malato).<br />
ADP<br />
ATP<br />
6<br />
Uma visão global <strong>do</strong> processo permite compreen<strong>de</strong>r que se diga que o oxalacetato<br />
tem, no ciclo <strong>de</strong> <strong>Krebs</strong>, um papel catalítico: tal como as enzimas (que em cada ciclo<br />
catalítico se ligam ao substrato e se regeneram como enzima livre após a formação <strong>do</strong><br />
produto)<br />
o oxalacetato reage com o acetil-CoA na “primeira” reacção <strong>do</strong> ciclo<br />
mas é regenera<strong>do</strong> na “última” quan<strong>do</strong> o malato é oxida<strong>do</strong>.<br />
Será mesmo mais correcto<br />
dizer que to<strong>do</strong>s os<br />
intermediários <strong>do</strong> ciclo têm<br />
um papel catalítico:<br />
quan<strong>do</strong> um molécula <strong>de</strong><br />
acetil-CoA é oxidada a CO 2<br />
os intermediários não se<br />
formam nem se consomem.<br />
O ciclo <strong>de</strong> <strong>Krebs</strong> é um “moinho” em<br />
que<br />
o “grão” (o substrato) é o grupo acetilo<br />
<strong>do</strong> acetil-CoA,<br />
a “farinha” (o produto) é o CO 2<br />
e<br />
a “mó <strong>do</strong> moinho” são as enzimas e os<br />
compostos intermediários.<br />
A análise <strong>do</strong> somatório das reacções que<br />
constituem o ciclo <strong>de</strong> <strong>Krebs</strong><br />
CH 3<br />
CO-CoA + 2 H 2<br />
O + 3 NAD + + FAD<br />
+ ADP + Pi<br />
→<br />
2 CO 2<br />
+ CoA + 3 NADH + FADH 2<br />
+ ATP<br />
mostra que, conceptualmente, este po<strong>de</strong> ser entendi<strong>do</strong> como um somatório <strong>de</strong> três<br />
processos:<br />
(1) a hidrólise <strong>do</strong> acetil-CoA (exergónico)<br />
CH 3<br />
CO-CoA + H 2<br />
O → CoA + CH 3<br />
COOH<br />
(2) a oxidação <strong>do</strong> resíduo <strong>de</strong> acetato a CO 2<br />
pelo NAD + e pelo FAD (exergónico)<br />
CH 3<br />
COOH + 2 H 2<br />
O + 3 NAD + + FAD → 2CO 2<br />
+ 3 NADH + FADH 2<br />
7<br />
(3) e a síntese <strong>de</strong> ATP a partir <strong>de</strong> ADP + Pi (en<strong>de</strong>rgónico).<br />
ADP + Pi → ATP + H2O<br />
8
O ciclo <strong>de</strong> <strong>Krebs</strong> tem um papel central no metabolismo oxidativo (catabolismo)<br />
porque<br />
(1) to<strong>do</strong>s os nutrientes que po<strong>de</strong>m ter um papel “energético” geram no seu<br />
catabolismo acetil-CoA e<br />
(2) a acetil-<br />
CoA é um<br />
intermediário<br />
obrigatório na<br />
oxidação<br />
<strong>de</strong>sses<br />
nutrientes.<br />
A velocida<strong>de</strong> com que o resíduo acetilo da acetil-CoA é oxidada a CO 2 no ciclo<br />
<strong>de</strong> <strong>Krebs</strong> <strong>de</strong>pen<strong>de</strong> em última análise da velocida<strong>de</strong> <strong>de</strong> consumo <strong>de</strong> ATP.<br />
acetil-CoA<br />
NADH<br />
NAD +<br />
NADH<br />
NAD +<br />
Desidrogénase<br />
<strong>do</strong> iso<strong>citrato</strong><br />
Desidrogénase <strong>do</strong><br />
α-cetoglutarato<br />
ADP e NAD +<br />
ATP e NADH<br />
ADP e NAD +<br />
ATP e NADH<br />
e Ca 2+<br />
e Ca 2+<br />
... a oxidação completa<br />
<strong>do</strong>s aminoáci<strong>do</strong>s que<br />
geram intermediários <strong>do</strong><br />
ciclo <strong>de</strong> <strong>Krebs</strong> só po<strong>de</strong><br />
ocorrer nos teci<strong>do</strong>s que<br />
tenham carboxicínase <strong>do</strong><br />
fosfoenolpiruvato<br />
(fíga<strong>do</strong>, rim, tec.<br />
adiposo, mama e<br />
intestino).<br />
9<br />
In vitro, o ADP e o NAD + activam as <strong>de</strong>sidrogénases <strong>do</strong> iso<strong>citrato</strong> e <strong>do</strong> α-cetoglutarato<br />
enquanto o ATP e o NADH são inibi<strong>do</strong>res...<br />
no entanto, é (pelo menos) duvi<strong>do</strong>so que estes compostos tenham um papel relevante<br />
na regulação da activida<strong>de</strong> <strong>de</strong>stas enzimas, in vivo.<br />
Actualmente, admite-se que um factores importantes na regulação da activida<strong>de</strong><br />
<strong>de</strong>stas enzimas, in vivo, seja o ião Ca 2+ . Ao contrário das concentrações <strong>de</strong> ATP,<br />
NADH e NAD + (cujas concentrações não variam) e <strong>do</strong> ADP (cuja concentração sofre<br />
variações ligeiras) o ião Ca 2+ aumenta <strong>de</strong> concentração (100 xs) quan<strong>do</strong> uma fibra 10<br />
muscular é excitada e se contrai.<br />
Diz-se que o ciclo <strong>de</strong> <strong>Krebs</strong> tem caracter anfibólico: as enzimas <strong>do</strong> ciclo <strong>de</strong><br />
<strong>Krebs</strong> para além <strong>de</strong> participarem na oxidação da acetil-CoA (catabolismo)<br />
também participam em processos anabólicos:<br />
1- síntese <strong>de</strong> glicose a partir<br />
<strong>de</strong> aminoáci<strong>do</strong>s<br />
2- síntese <strong>de</strong> áci<strong>do</strong>s<br />
gor<strong>do</strong>s a partir <strong>de</strong><br />
acetil-CoA<br />
A utilização <strong>de</strong> intermediários <strong>do</strong> ciclo <strong>de</strong> <strong>Krebs</strong> em processos <strong>de</strong> anabólicos<br />
ten<strong>de</strong> a “esvaziar” o ciclo <strong>de</strong> <strong>Krebs</strong> mas...<br />
a concentração <strong>do</strong>s intermediários <strong>do</strong> ciclo <strong>de</strong> <strong>Krebs</strong> varia muito<br />
p<strong>ou</strong>co porque por cada molécula que sai há uma que entra.<br />
piruvato<br />
CO 2<br />
+<br />
ATP<br />
Carboxílase <strong>do</strong><br />
piruvato ADP + Pi<br />
fosfoenolpiruvato<br />
A reacção catalisada pela síntase <strong>do</strong> <strong>citrato</strong><br />
não é anaplerótica nem cataplerótica<br />
Além disto porque a<br />
reacção catalisada pela<br />
<strong>de</strong>sidrogénase <strong>do</strong><br />
piruvato é irreversível<br />
a acetil-CoA não po<strong>de</strong><br />
gerar oxalacetato.<br />
3- síntese <strong>de</strong><br />
aminoáci<strong>do</strong>s<br />
11<br />
Reacção<br />
cataplerótica<br />
intermediário <strong>do</strong> ciclo →<br />
não intermediário <strong>do</strong> ciclo<br />
(ten<strong>de</strong>m a diminuir as<br />
“mós <strong>do</strong> moinho”)<br />
α-cetoglutarato<br />
glutamato<br />
Reacção anaplerótica<br />
não intermediário <strong>do</strong> ciclo →<br />
intermediário <strong>do</strong> ciclo<br />
(ten<strong>de</strong>m a aumentar as “mós<br />
<strong>do</strong> moinho”)<br />
12
Os áci<strong>do</strong>s gor<strong>do</strong>s <strong>de</strong> ca<strong>de</strong>ia par (a maioria)<br />
não são glicogénicos porque só geram acetil-<br />
CoA no seu processo catabólico.<br />
Para engordar não é necessário comer<br />
gorduras...<br />
também serve comer glicí<strong>de</strong>os.<br />
Alguns aminoáci<strong>do</strong>s po<strong>de</strong>m originar<br />
intermediários <strong>do</strong> ciclo <strong>de</strong> <strong>Krebs</strong>;<br />
porque os intermediários <strong>do</strong> ciclo <strong>de</strong><br />
<strong>Krebs</strong> po<strong>de</strong>m gerar glicose, esses<br />
aminoáci<strong>do</strong>s são glicogénicos.<br />
O glutamato é,<br />
por exemplo, um<br />
aminoáci<strong>do</strong><br />
As enzimas da glicólise, a <strong>de</strong>sidrogénase <strong>do</strong><br />
piruvato, a síntase <strong>do</strong> <strong>citrato</strong> e a líase <strong>do</strong> ATP<strong>citrato</strong><br />
participam na formação <strong>de</strong> áci<strong>do</strong>s gor<strong>do</strong>s<br />
a partir <strong>de</strong> glicose<br />
glicogénico. 13<br />
14<br />
O acetil-CoA é precursor na formação <strong>do</strong>s<br />
áci<strong>do</strong>s gor<strong>do</strong>s mas...<br />
(1) a sua síntese ocorre <strong>de</strong>ntro da mitocôndria e<br />
(2) a formação <strong>do</strong>s áci<strong>do</strong>s gor<strong>do</strong>s ocorre no<br />
citoplasma.<br />
Na membrana da mitocôndria<br />
não existe nenhum<br />
transporta<strong>do</strong>r para o acetil-CoA<br />
mas existe para o <strong>citrato</strong>.<br />
Via <strong>citrato</strong> o acetil-CoA po<strong>de</strong><br />
ser “transporta<strong>do</strong>” para o<br />
citosol.<br />
15