138 SANTOS, MAYORAL, MUÑIZ, BAJO y ADRIAENSENS
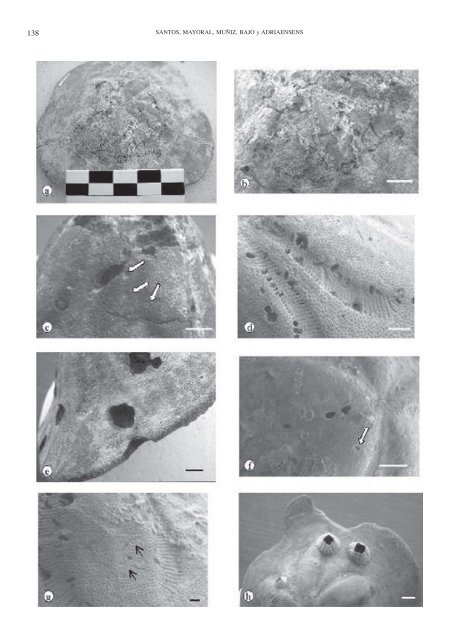
En líneas gener<strong>al</strong>es, y a la vista <strong>de</strong> la distribución <strong>de</strong> todas las evi<strong>de</strong>ncias relacionadas con la actividad <strong>de</strong>predadora (Figs. 3, 4, 6), se <strong>de</strong>duce que éstas <strong>de</strong>bieron producirse en dos estados diferentes <strong>de</strong>l sustrato, una cuando los erizos todavía estaban vivos y otra en una fase inmediatamente post-mortem. Al primer caso pertenecerían todas las roturas y mordiscos en el ambitus, puesto que presentan señ<strong>al</strong>es inequívocas <strong>de</strong> haber sido reparados, mientras que en el segundo caso habría que incluir todas las perforaciones relacionadas con Oichnus. Éstos se loc<strong>al</strong>izan preferentemente en las zonas poríferas <strong>de</strong> los ambulacros, lo que indicaría la búsqueda por parte <strong>de</strong>l <strong>de</strong>predador <strong>de</strong> las partes blandas más accesibles (los podia). Este comportamiento más que <strong>de</strong>predador sería necrófago, ya que los erizos no muestran señ<strong>al</strong>es <strong>de</strong> reparación asociadas a las perforaciones (como es habitu<strong>al</strong> en estos casos; Ab<strong>de</strong>lhamid, 1999), ni tan siquiera removilización <strong>de</strong> materi<strong>al</strong> secundario en las áreas adyacentes (Nebelsick, 1999a), por lo que las presas <strong>de</strong>berían estar ya muertas; eso sí, en una fase muy reciente. El resto <strong>de</strong> las estructuras bioerosivas <strong>de</strong>notan sólo la búsqueda y/o construcción <strong>de</strong> un domicilio permanente y están representadas por Gastrochaenolites, relacionadas con la actividad perforante <strong>de</strong> biv<strong>al</strong>vos litófagos, así como por Maeandropolydora, en relación con la actividad mayoritaria (y presumiblemente) <strong>de</strong> anélidos. Ambas estructuras se han producido en fases claramente post-mortem <strong>de</strong>l sustrato. En el caso <strong>de</strong> Gastrochaenolites, éstos se ubican <strong>de</strong> forma preferente en la zona p<strong>et</strong><strong>al</strong>oi<strong>de</strong>a <strong>de</strong> la cara abor<strong>al</strong>, que es la región más prominente y elevada <strong>de</strong>l caparazón, una vez que éste queda abandonado <strong>al</strong> morir el anim<strong>al</strong>. De este modo, los erizos se constituyen en pequeñas “islas” repartidas por BIOEROSIÓN EN ERIZOS IRREGULARES DEL MIOCENO SUPERIOR 139 el fondo, que son ocupadas masivamente por los biv<strong>al</strong>vos litófagos y esporádicamente por los anélidos, amén <strong>de</strong> otro tipo <strong>de</strong> organismos incrustantes (princip<strong>al</strong>mente b<strong>al</strong>anomorfos). El hecho <strong>de</strong> que la gran mayoría <strong>de</strong> las evi<strong>de</strong>ncias (el 97 %) se presente en la cara abor<strong>al</strong> implica que la energía <strong>de</strong>l medio no <strong>de</strong>bió <strong>de</strong> ser muy elevada, por lo que los caparazones no llegaron a dar la vuelta (<strong>al</strong> menos la mayor parte <strong>de</strong> ellos). A<strong>de</strong>más las evi<strong>de</strong>ncias <strong>de</strong> impactos tampoco son especi<strong>al</strong>mente importantes (1%), lo cu<strong>al</strong> apoyaría esta hipótesis. Es muy factible que se llegara a conseguir un equilibrio tempor<strong>al</strong> entre la tasa <strong>de</strong> sedimentación y la energía, <strong>de</strong> t<strong>al</strong> forma que a una primera <strong>et</strong>apa <strong>de</strong> enterramiento relativamente rápido, que <strong>de</strong>jara <strong>al</strong> <strong>de</strong>scubierto la cúpula p<strong>et</strong><strong>al</strong>oi<strong>de</strong>a <strong>de</strong> los caparazones, siguiera una fase <strong>de</strong> d<strong>et</strong>ención o r<strong>al</strong>entización <strong>de</strong> la sedimentación, que permitiera la acción <strong>de</strong>predadora-carroñera <strong>de</strong> los gasterópodos y la ocupación <strong>de</strong>l sustrato por la mayor parte <strong>de</strong> los organismos incrustantes (b<strong>al</strong>anomorfos, anélidos) y perforantes (biv<strong>al</strong>vos litófagos). La fase <strong>de</strong>predadora no <strong>de</strong>bió <strong>de</strong> ser muy larga, ya que el porcentaje <strong>de</strong> ataques es bajo (15%) frente <strong>al</strong> grado <strong>de</strong> ocupación por parte <strong>de</strong> los biv<strong>al</strong>vos y anélidos, que es relativamente mucho más significativo (27%). Estos porcentajes <strong>de</strong> <strong>de</strong>predación son anorm<strong>al</strong>mente bajos y se sitúan más en la esc<strong>al</strong>a <strong>de</strong> los v<strong>al</strong>ores observados para los erizos <strong>de</strong>l Cr<strong>et</strong>ácico que suele ser inferior a un 10% (Ab<strong>de</strong>lhamid, 1999; Nebelsick y Kow<strong>al</strong>ewski, 2001) o <strong>de</strong>l P<strong>al</strong>eógeno, entre un 5 y un 10% (Woodcock y Kelley, 2001). Las razones para esta baja tasa <strong>de</strong>ben estar más en relación con las características propias <strong>de</strong>l medio que con la f<strong>al</strong>ta <strong>de</strong> <strong>de</strong>predadores potenci<strong>al</strong>es, cuyo número en el Mioceno superior ya era relativamente importante. Figura 7. a. Vista gener<strong>al</strong> <strong>de</strong> Clypeaster sp. mostrando el área p<strong>et</strong><strong>al</strong>oi<strong>de</strong>a <strong>de</strong>l caparazón con Oichnus simplex Bromley, 1981. Ejemplar MGUS-2060. Esc<strong>al</strong>a: 10 cm. b. D<strong>et</strong><strong>al</strong>le <strong>de</strong> la zona p<strong>et</strong><strong>al</strong>oi<strong>de</strong>a con Oichnus simplex Bromley, 1981, que han llegado a <strong>de</strong>struir parci<strong>al</strong>mente los ambulacros. Esc<strong>al</strong>a <strong>de</strong> la barra: 1 cm. c. Clypeaster portentosus con Gastrochaenolites cf. digujus Kelley y Bromley, 1984 . Las flechas señ<strong>al</strong>an <strong>al</strong>gunos hundimientos <strong>de</strong>l caparazón. Ejemplar MGUS-2061. Esc<strong>al</strong>a <strong>de</strong> la barra: 1 cm. d. D<strong>et</strong><strong>al</strong>le <strong>de</strong> Clypeaster cermenatii con Gastrochaenolites cf. digujus ubicados en la zona porifera <strong>de</strong> los ambulacros. Ejemplar MGUS-2070. Esc<strong>al</strong>a <strong>de</strong> la barra:1 cm. e. D<strong>et</strong><strong>al</strong>le <strong>de</strong> Gastrochaenolites cf. digujus en el bor<strong>de</strong> <strong>de</strong>l ambitus <strong>de</strong> Clypeaster cermenatii. Ejemplar MGUS-2059. Esc<strong>al</strong>a <strong>de</strong> la barra: 1 cm. f. Gastrochaenolites cf. digujus en el ambulacro <strong>de</strong> Clypeaster cermenatii. La flecha señ<strong>al</strong>a un hundimiento irregular <strong>de</strong>l caparazón. Ejemplar MGUS-2064. Esc<strong>al</strong>a <strong>de</strong> la barra: 1 cm. g. Maeandropolydora sulcans (flechas) en el ambulacro <strong>de</strong> Clypeaster cermenatii. Ejemplar MGUS-2070. Esc<strong>al</strong>a <strong>de</strong> la barra: 1 cm. h. Mordisco en el ambitus <strong>de</strong> Clypeaster sp. Ejemplar MGUS-2063. Esc<strong>al</strong>a <strong>de</strong> la barra: 1 cm. a. Gener<strong>al</strong> view of Clypeaster sp. showing Oichnus simplex Bromley, 1981 on the p<strong>et</strong><strong>al</strong>odium zone of the test. Specimen MGUS-2060. Sc<strong>al</strong>e bar: 10 cm. b. D<strong>et</strong>ail of the p<strong>et</strong><strong>al</strong>odium zone with Oichnus simplex Bromley, 1981 causing parti<strong>al</strong> <strong>de</strong>struction of the ambulacra. Sc<strong>al</strong>e bar: 1 cm. c. Clypeaster portentosus with Gastrochaenolites cf. digujus Kelley y Bromley, 1984. Arrows point to some irregular <strong>de</strong>pressions of the test. Specimen MGUS-2061. Sc<strong>al</strong>e bar: 1 cm. d. D<strong>et</strong>ail of Clypeaster cermenatii with Gastrochaenolites cf. digujus located on the poriferous zone of the ambulacra. Specimen MGUS-2070. Sc<strong>al</strong>e bar: 1 cm. e. D<strong>et</strong>ail of Gastrochaenolites cf. digujus Kelley y Bromley, 1984 in the ambitus of Clypeaster cermenatii. Specimen MGUS-2059. Sc<strong>al</strong>e bar: 1 cm. f. Gastrochaenolites cf. digujus on the ambulacrum of Clypeaster cermenatii. Arrow point to an irregular <strong>de</strong>pression of the test. Specimen MGUS-2064. Sc<strong>al</strong>e bar: 1 cm. g. Maeandropolydora sulcans (arrows) on the ambulacrum of Clypeaster cermenatii. Specimen MGUS-2070. Sc<strong>al</strong>e bar: 1 cm. h. Bite on the ambitus of Clypeaster sp. Specimen MGUS-2063. Sc<strong>al</strong>e bar: 1 cm.