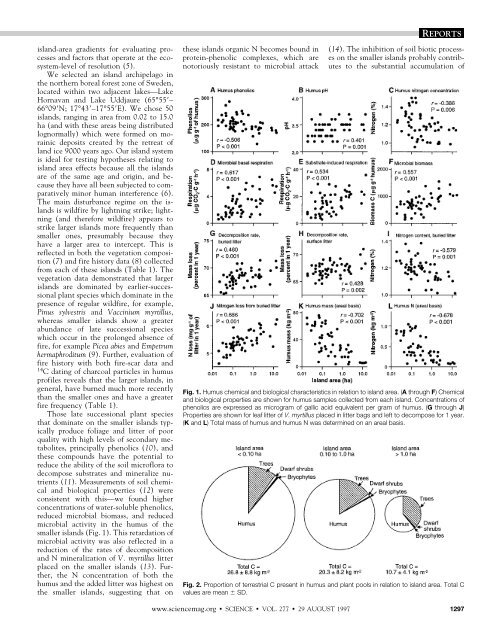
REPORTSisl<strong>and</strong>-area gradients for evaluating processes<strong>and</strong> factors that operate at the ecosystem-level<str<strong>on</strong>g>of</str<strong>on</strong>g> resoluti<strong>on</strong> (5).We selected an isl<strong>and</strong> archipelago inthe northern boreal forest z<strong>on</strong>e <str<strong>on</strong>g>of</str<strong>on</strong>g> Sweden,located within two adjacent lakes—LakeHornavan <strong>and</strong> Lake Uddjaure (65°55–66°09N; 17°43–17°55E). We chose 50isl<strong>and</strong>s, ranging in area from 0.02 to 15.0ha (<strong>and</strong> with these areas being distributedlognormally) which were formed <strong>on</strong> morainicdeposits created by the retreat <str<strong>on</strong>g>of</str<strong>on</strong>g>l<strong>and</strong> ice 9000 years ago. Our isl<strong>and</strong> systemis ideal for testing hypotheses relating toisl<strong>and</strong> area effects because all the isl<strong>and</strong>sare <str<strong>on</strong>g>of</str<strong>on</strong>g> the same age <strong>and</strong> origin, <strong>and</strong> becausethey have all been subjected to comparativelyminor human interference (6).<str<strong>on</strong>g>The</str<strong>on</strong>g> main disturbance regime <strong>on</strong> the isl<strong>and</strong>sis wildfire by lightning strike; lightning(<strong>and</strong> therefore wildfire) appears tostrike larger isl<strong>and</strong>s more frequently thansmaller <strong>on</strong>es, presumably because theyhave a larger area to intercept. This isreflected in both the vegetati<strong>on</strong> compositi<strong>on</strong>(7) <strong>and</strong> fire history data (8) collectedfrom each <str<strong>on</strong>g>of</str<strong>on</strong>g> these isl<strong>and</strong>s (Table 1). <str<strong>on</strong>g>The</str<strong>on</strong>g>vegetati<strong>on</strong> data dem<strong>on</strong>strated that largerisl<strong>and</strong>s are dominated by earlier-successi<strong>on</strong>alplant species which dominate in thepresence <str<strong>on</strong>g>of</str<strong>on</strong>g> regular wildfire, for example,Pinus sylvestris <strong>and</strong> Vaccinium myrtillus,whereas smaller isl<strong>and</strong>s show a greaterabundance <str<strong>on</strong>g>of</str<strong>on</strong>g> late successi<strong>on</strong>al specieswhich occur in the prol<strong>on</strong>ged absence <str<strong>on</strong>g>of</str<strong>on</strong>g>fire, for example Picea abies <strong>and</strong> Empetrumhermaphroditum (9). Further, evaluati<strong>on</strong> <str<strong>on</strong>g>of</str<strong>on</strong>g>fire history with both fire-scar data <strong>and</strong>14 C dating <str<strong>on</strong>g>of</str<strong>on</strong>g> charcoal particles in humuspr<str<strong>on</strong>g>of</str<strong>on</strong>g>iles reveals that the larger isl<strong>and</strong>s, ingeneral, have burned much more recentlythan the smaller <strong>on</strong>es <strong>and</strong> have a greaterfire frequency (Table 1).Those late successi<strong>on</strong>al plant speciesthat dominate <strong>on</strong> the smaller isl<strong>and</strong>s typicallyproduce foliage <strong>and</strong> litter <str<strong>on</strong>g>of</str<strong>on</strong>g> poorquality with high levels <str<strong>on</strong>g>of</str<strong>on</strong>g> sec<strong>on</strong>dary metabolites,principally phenolics (10), <strong>and</strong>these compounds have the potential toreduce the ability <str<strong>on</strong>g>of</str<strong>on</strong>g> the soil micr<str<strong>on</strong>g>of</str<strong>on</strong>g>lora todecompose substrates <strong>and</strong> mineralize nutrients(11). Measurements <str<strong>on</strong>g>of</str<strong>on</strong>g> soil chemical<strong>and</strong> biological properties (12) werec<strong>on</strong>sistent with this—we found higherc<strong>on</strong>centrati<strong>on</strong>s <str<strong>on</strong>g>of</str<strong>on</strong>g> water-soluble phenolics,reduced microbial biomass, <strong>and</strong> reducedmicrobial activity in the humus <str<strong>on</strong>g>of</str<strong>on</strong>g> thesmaller isl<strong>and</strong>s (Fig. 1). This retardati<strong>on</strong> <str<strong>on</strong>g>of</str<strong>on</strong>g>microbial activity was also reflected in areducti<strong>on</strong> <str<strong>on</strong>g>of</str<strong>on</strong>g> the rates <str<strong>on</strong>g>of</str<strong>on</strong>g> decompositi<strong>on</strong><strong>and</strong> N mineralizati<strong>on</strong> <str<strong>on</strong>g>of</str<strong>on</strong>g> V. myrtillus litterplaced <strong>on</strong> the smaller isl<strong>and</strong>s (13). Further,the N c<strong>on</strong>centrati<strong>on</strong> <str<strong>on</strong>g>of</str<strong>on</strong>g> both thehumus <strong>and</strong> the added litter was highest <strong>on</strong>the smaller isl<strong>and</strong>s, suggesting that <strong>on</strong>these isl<strong>and</strong>s organic N becomes bound inprotein-phenolic complexes, which arenotoriously resistant to microbial attack(14). <str<strong>on</strong>g>The</str<strong>on</strong>g> inhibiti<strong>on</strong> <str<strong>on</strong>g>of</str<strong>on</strong>g> soil biotic processes<strong>on</strong> the smaller isl<strong>and</strong>s probably c<strong>on</strong>tributesto the substantial accumulati<strong>on</strong> <str<strong>on</strong>g>of</str<strong>on</strong>g>Fig. 1. Humus chemical <strong>and</strong> biological characteristics in relati<strong>on</strong> to isl<strong>and</strong> area. (A through F) Chemical<strong>and</strong> biological properties are shown for humus samples collected from each isl<strong>and</strong>. C<strong>on</strong>centrati<strong>on</strong>s <str<strong>on</strong>g>of</str<strong>on</strong>g>phenolics are expressed as microgram <str<strong>on</strong>g>of</str<strong>on</strong>g> gallic acid equivalent per gram <str<strong>on</strong>g>of</str<strong>on</strong>g> humus. (G through J)<strong>Properties</strong> are shown for leaf litter <str<strong>on</strong>g>of</str<strong>on</strong>g> V. myrtillus placed in litter bags <strong>and</strong> left to decompose for 1 year.(K <strong>and</strong> L) Total mass <str<strong>on</strong>g>of</str<strong>on</strong>g> humus <strong>and</strong> humus N was determined <strong>on</strong> an areal basis.Fig. 2. Proporti<strong>on</strong> <str<strong>on</strong>g>of</str<strong>on</strong>g> terrestrial C present in humus <strong>and</strong> plant pools in relati<strong>on</strong> to isl<strong>and</strong> area. Total Cvalues are mean SD.www.sciencemag.org SCIENCE VOL. 277 29 AUGUST 1997 1297
humus that occurs <strong>on</strong> them (Fig. 1); thesmallest isl<strong>and</strong>s c<strong>on</strong>tain up to 10 timesmore humus per unit area than do thelargest <strong>on</strong>es. <str<strong>on</strong>g>The</str<strong>on</strong>g>refore, our results dem<strong>on</strong>stratethat isl<strong>and</strong> area is critical in regulatingkey ecological processes (<strong>and</strong> thereforeecosystem functi<strong>on</strong>) <strong>and</strong> that this ismost likely attributable to plant specieseffects.Carb<strong>on</strong> partiti<strong>on</strong>ing is clearly regulatedby isl<strong>and</strong> area. With increasing isl<strong>and</strong> size,there is a distinct trend <str<strong>on</strong>g>of</str<strong>on</strong>g> an increasingTable 1. Vegetati<strong>on</strong> <strong>and</strong> fire history characteristics in relati<strong>on</strong> to isl<strong>and</strong> area. Values presented aremeans SE.Species/measurement0.1 ha(n 14)<str<strong>on</strong>g>Area</str<strong>on</strong>g> class0.1–1.0 ha(n 24)1.0 ha(n 12)Tree stem density (N o /ha)Pinus sylvestris 206 82 222 48 410 78Betula pubescens 1233 229 1707 186 982 214Picea abies 278 87 211 48 45 22Total 1720 198 2140 169 1438 201Tree biomass (kg/m 2 )Pinus sylvestris 1.697 0.627 2.627 0.563 6.160 0.864Betula pubescens 0.844 0.198 1.287 0.211 0.887 0.199Picea abies 1.329 0.323 1.462 0.289 0.332 0.169Total 3.869 0.624 5.376 0.466 7.385 0.826Dwarf shrub biomass (kg/m 2 )Vaccinium myrtillus 0.069 0.014 0.083 0.011 0.119 0.017Vaccinium vitis-idaea 0.164 0.030 0.215 0.021 0.153 0.025Empetrum hermaphroditum 0.129 0.024 0.075 0.010 0.060 0.018Total 0.365 0.022 0.383 0.021 0.332 0.033Bryophyte biomass (kg/m 2 )Pleurozium schreberi 0.152 0.038 0.143 0.030 0.155 0.030Hylocomium splendens 0.121 0.021 0.061 0.016 0.077 0.021Total 0.283 0.033 0.217 0.027 0.261 0.035Additi<strong>on</strong>al parametersTotal vegetati<strong>on</strong> biomass 4.517 0.616 5.977 0.455 7.979 0.817(kg/m 2 )Shann<strong>on</strong>-Weiner species 1.784 0.131 1.465 0.065 1.178 0.134diversity index (H / )*Number <str<strong>on</strong>g>of</str<strong>on</strong>g> fires in past 250 0.143 0.097 0.208 0.085 0.667 0.256yearsAge <str<strong>on</strong>g>of</str<strong>on</strong>g> charcoal in years( 14 C)†2984 340 2081 428 1106 495*Index calculated by use <str<strong>on</strong>g>of</str<strong>on</strong>g> biomass values for each plant species. †Obtained by 14 C radiocarb<strong>on</strong> dating <str<strong>on</strong>g>of</str<strong>on</strong>g>uppermost charcoal layer in humus pr<str<strong>on</strong>g>of</str<strong>on</strong>g>ile; n 5 isl<strong>and</strong>s used for each area class.Fig. 3. Total N c<strong>on</strong>centrati<strong>on</strong><str<strong>on</strong>g>of</str<strong>on</strong>g> selected organismsin relati<strong>on</strong> to isl<strong>and</strong>area. <str<strong>on</strong>g>The</str<strong>on</strong>g> relati<strong>on</strong>ship<str<strong>on</strong>g>of</str<strong>on</strong>g> humus N c<strong>on</strong>centrati<strong>on</strong>to isl<strong>and</strong> areais included for comparativepurposes. <str<strong>on</strong>g>The</str<strong>on</strong>g> Nc<strong>on</strong>centrati<strong>on</strong> <str<strong>on</strong>g>of</str<strong>on</strong>g> leavesfrom V. myrtillus <strong>and</strong> V.vitis-idaea did not showa statistically significantrelati<strong>on</strong>ship with isl<strong>and</strong>area.proporti<strong>on</strong> <str<strong>on</strong>g>of</str<strong>on</strong>g> organic C that is bound inliving organisms, especially trees (Fig. 2).Because smaller isl<strong>and</strong>s c<strong>on</strong>tain much higherterrestrial C levels <strong>on</strong> an areal basis th<strong>and</strong>o larger <strong>on</strong>es, our data indicates that wildfireis <str<strong>on</strong>g>of</str<strong>on</strong>g> critical importance (either directlyor indirectly) in reversing C lock-up inboreal forest ecosystems. This finding suggeststhat deliberate anthropogenic supressi<strong>on</strong><str<strong>on</strong>g>of</str<strong>on</strong>g> fires in boreal forests over the pastcentury has the potential to lead to retardati<strong>on</strong><str<strong>on</strong>g>of</str<strong>on</strong>g> soil biological processes <strong>and</strong> substantialterrestrial C sequestrati<strong>on</strong> (15),which is likely to be <str<strong>on</strong>g>of</str<strong>on</strong>g> global significancegiven the role <str<strong>on</strong>g>of</str<strong>on</strong>g> boreal forests in the globalC cycle (16).<str<strong>on</strong>g>The</str<strong>on</strong>g>re is evidence that N partiti<strong>on</strong>ing isalso likely to be affected by isl<strong>and</strong> area.Although the humus N c<strong>on</strong>tent was negativelyrelated to isl<strong>and</strong> area, the N c<strong>on</strong>centrati<strong>on</strong>s<str<strong>on</strong>g>of</str<strong>on</strong>g> leaves <str<strong>on</strong>g>of</str<strong>on</strong>g> E. hermaphroditum,shoots <str<strong>on</strong>g>of</str<strong>on</strong>g> the moss Pleurozium schreberi,<strong>and</strong> the soil microbial biomass were allpositively related to isl<strong>and</strong> area (Fig. 3).This is indicative <str<strong>on</strong>g>of</str<strong>on</strong>g> enhanced acquisiti<strong>on</strong><str<strong>on</strong>g>of</str<strong>on</strong>g> N by these organisms, <strong>and</strong> means thatthe N present in the humus <str<strong>on</strong>g>of</str<strong>on</strong>g> the smallerisl<strong>and</strong>s is clearly less available to livingorganisms than that <str<strong>on</strong>g>of</str<strong>on</strong>g> the larger isl<strong>and</strong>s.This again supports the c<strong>on</strong>cept that higherlevels <str<strong>on</strong>g>of</str<strong>on</strong>g> phenolics <strong>on</strong> the smaller isl<strong>and</strong>sc<strong>on</strong>tribute to reduced N availability.<str<strong>on</strong>g>The</str<strong>on</strong>g> lower plant tissue N c<strong>on</strong>tent <str<strong>on</strong>g>of</str<strong>on</strong>g> plants<strong>on</strong> smaller isl<strong>and</strong>s would be expected toresult in subsequently produced plant litterwith a lower C:N ratio, <strong>and</strong> thus, areduced rate <str<strong>on</strong>g>of</str<strong>on</strong>g> decompositi<strong>on</strong> (17), ultimatelyresulting in further organic matteraccumulati<strong>on</strong>.<str<strong>on</strong>g>Isl<strong>and</strong></str<strong>on</strong>g> area studies have potential forinvestigati<strong>on</strong> <str<strong>on</strong>g>of</str<strong>on</strong>g> relati<strong>on</strong>ships between speciesdiversity <strong>and</strong> ecosystem-level properties(5). In our study, plant species diversitywas highest <strong>on</strong> the smallest isl<strong>and</strong>s(Table 1), meaning that ecosystem processrates were lowest <strong>on</strong> those isl<strong>and</strong>s with thegreatest diversity. This finding is in directc<strong>on</strong>trast to other studies which haveshown elevated process rates in more diversecommunities (18). However, wefound that the plant biomass <str<strong>on</strong>g>of</str<strong>on</strong>g> the largerisl<strong>and</strong>s was dominated by a single earlysuccessi<strong>on</strong>al competitive tree species, P.sylvestris, which has comparatively favorablelitter quality but which c<strong>on</strong>tributes toa reduced species diversity. <str<strong>on</strong>g>The</str<strong>on</strong>g> greaterecological stresses <strong>on</strong> the smaller isl<strong>and</strong>s(due to higher phenolics, lower pH, <strong>and</strong>reduced N availability) presumably preventeddominance by a single competitivespecies, resulting in a greater diversity(19), but with a greater abundance <str<strong>on</strong>g>of</str<strong>on</strong>g>those plant species with traits likely toc<strong>on</strong>tribute to retarding ecosystem-levelprocesses. Other possible explanati<strong>on</strong>s for1298SCIENCE VOL. 277 29 AUGUST 1997 www.sciencemag.org