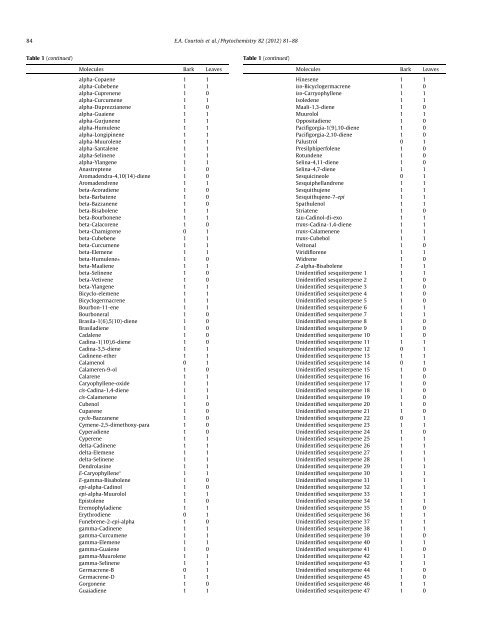
84 E.A. Courtois et al. / Phytochemistry 82 (2012) 81–88Table 1 (cont<strong>in</strong>ued)Table 1 (cont<strong>in</strong>ued)Molecules Bark Leavesalpha-Copaene 1 1alpha-Cubebene 1 1alpha-Cuprenene 1 0alpha-Curcumene 1 1alpha-Duprezzianene 1 0alpha-Guaiene 1 1alpha-Gurjunene 1 1alpha-Humulene 1 1alpha-Longip<strong>in</strong>ene 1 1alpha-Muurolene 1 1alpha-Santalene 1 1alpha-Sel<strong>in</strong>ene 1 1alpha-Ylangene 1 1Anastreptene 1 0Aromadendra-4,10(14)-diene 1 0Aromadendrene 1 1beta-Acoradiene 1 0beta-Barbatene 1 0beta-Bazzanene 1 0beta-Bisabolene 1 1beta-Bourbonene 1 1beta-Calacorene 1 0beta-Chamigrene 0 1beta-Cubebene 1 1beta-Curcumene 1 1beta-Elemene 1 1beta-Humulene⁄ 1 0beta-Maaliene 1 1beta-Sel<strong>in</strong>ene 1 0beta-Vetivene 1 0beta-Ylangene 1 1Bicyclo-elemene 1 1Bicyclogermacrene 1 1Bourbon-11-ene 1 1Bourboneral 1 0Brasila-1(6),5(10)-diene 1 0Brasiladiene 1 0Cadalene 1 0Cad<strong>in</strong>a-1(10),6-diene 1 0Cad<strong>in</strong>a-3,5-diene 1 1Cad<strong>in</strong>ene-ether 1 1Calamenol 0 1Calameren-9-ol 1 0Calarene 1 1Caryophyllene-oxide 1 1cis-Cad<strong>in</strong>a-1,4-diene 1 1cis-Calamenene 1 1Cubenol 1 0Cuparene 1 0cyclo-Bazzanene 1 0Cymene-2,5-dimethoxy-para 1 0Cyperadiene 1 0Cyperene 1 1delta-Cad<strong>in</strong>ene 1 1delta-Elemene 1 1delta-Sel<strong>in</strong>ene 1 1Dendrolas<strong>in</strong>e 1 1E-Caryophyllene ⁄ 1 1E-gamma-Bisabolene 1 0epi-alpha-Cad<strong>in</strong>ol 1 0epi-alpha-Muurolol 1 1Epistolene 1 0Eremophyladiene 1 1Erythrodiene 0 1Funebrene-2-epi-alpha 1 0gamma-Cad<strong>in</strong>ene 1 1gamma-Curcumene 1 1gamma-Elemene 1 1gamma-Guaiene 1 0gamma-Muurolene 1 1gamma-Sel<strong>in</strong>ene 1 1Germacrene-B 0 1Germacrene-D 1 1Gorgonene 1 0Guaiadiene 1 1Molecules Bark LeavesH<strong>in</strong>esene 1 1iso-Bicyclogermacrene 1 0iso-Carryophyllene 1 1Isoledene 1 1Maali-1,3-diene 1 0Muurolol 1 1Oppositadiene 1 0Pacifigorgia-1(9),10-diene 1 0Pacifigorgia-2,10-diene 1 0Palustrol 0 1Presilphiperfolene 1 0Rotundene 1 0Sel<strong>in</strong>a-4,11-diene 1 0Sel<strong>in</strong>a-4,7-diene 1 1Sesquic<strong>in</strong>eole 0 1Sesquiphellandrene 1 1Sesquithujene 1 1Sesquithujene-7-epi 1 1Spathulenol 1 1Striatene 1 0tau-Cad<strong>in</strong>ol-di-exo 1 1trans-Cad<strong>in</strong>a-1,4-diene 1 1trans-Calamenene 1 1trans-Cubebol 1 1Veltonal 1 0Viridiflorene 1 1Widrene 1 0Z-alpha-Bisabolene 1 1Unidentified sesqui<strong>terpene</strong> 1 1 1Unidentified sesqui<strong>terpene</strong> 2 1 0Unidentified sesqui<strong>terpene</strong> 3 1 0Unidentified sesqui<strong>terpene</strong> 4 1 0Unidentified sesqui<strong>terpene</strong> 5 1 0Unidentified sesqui<strong>terpene</strong> 6 1 1Unidentified sesqui<strong>terpene</strong> 7 1 1Unidentified sesqui<strong>terpene</strong> 8 1 0Unidentified sesqui<strong>terpene</strong> 9 1 0Unidentified sesqui<strong>terpene</strong> 10 1 0Unidentified sesqui<strong>terpene</strong> 11 1 1Unidentified sesqui<strong>terpene</strong> 12 0 1Unidentified sesqui<strong>terpene</strong> 13 1 1Unidentified sesqui<strong>terpene</strong> 14 0 1Unidentified sesqui<strong>terpene</strong> 15 1 0Unidentified sesqui<strong>terpene</strong> 16 1 0Unidentified sesqui<strong>terpene</strong> 17 1 0Unidentified sesqui<strong>terpene</strong> 18 1 0Unidentified sesqui<strong>terpene</strong> 19 1 0Unidentified sesqui<strong>terpene</strong> 20 1 0Unidentified sesqui<strong>terpene</strong> 21 1 0Unidentified sesqui<strong>terpene</strong> 22 0 1Unidentified sesqui<strong>terpene</strong> 23 1 1Unidentified sesqui<strong>terpene</strong> 24 1 0Unidentified sesqui<strong>terpene</strong> 25 1 1Unidentified sesqui<strong>terpene</strong> 26 1 1Unidentified sesqui<strong>terpene</strong> 27 1 1Unidentified sesqui<strong>terpene</strong> 28 1 1Unidentified sesqui<strong>terpene</strong> 29 1 1Unidentified sesqui<strong>terpene</strong> 30 1 1Unidentified sesqui<strong>terpene</strong> 31 1 1Unidentified sesqui<strong>terpene</strong> 32 1 1Unidentified sesqui<strong>terpene</strong> 33 1 1Unidentified sesqui<strong>terpene</strong> 34 1 1Unidentified sesqui<strong>terpene</strong> 35 1 0Unidentified sesqui<strong>terpene</strong> 36 1 1Unidentified sesqui<strong>terpene</strong> 37 1 1Unidentified sesqui<strong>terpene</strong> 38 1 1Unidentified sesqui<strong>terpene</strong> 39 1 0Unidentified sesqui<strong>terpene</strong> 40 1 1Unidentified sesqui<strong>terpene</strong> 41 1 0Unidentified sesqui<strong>terpene</strong> 42 1 1Unidentified sesqui<strong>terpene</strong> 43 1 1Unidentified sesqui<strong>terpene</strong> 44 1 0Unidentified sesqui<strong>terpene</strong> 45 1 0Unidentified sesqui<strong>terpene</strong> 46 1 1Unidentified sesqui<strong>terpene</strong> 47 1 0
E.A. Courtois et al. / Phytochemistry 82 (2012) 81–88 85Table 1 (cont<strong>in</strong>ued)200Molecules Bark LeavesUnidentified sesqui<strong>terpene</strong> 48 1 0Unidentified sesqui<strong>terpene</strong> 49 1 0Unidentified sesqui<strong>terpene</strong> 50 1 1Only barkBoth tissuesOnly leavesoverlap <strong>in</strong> the <strong>terpene</strong> <strong>composition</strong> of the bark and the leaves of agiven <strong>in</strong>dividual but our result show that some compounds mayonly be expressed <strong>in</strong> one tissue. One hypothesis to expla<strong>in</strong> this isthe fact that the herbivore and pathogen communities associatedwith the bark and the leaves are different (Novotny et al., 2003).A dist<strong>in</strong>ct herbivory and pathogen pressure could therefore favor<strong>in</strong> plants a dist<strong>in</strong>ct allocation of defensive compounds.We further found that some compounds seem to be restrictedto one tissue (either bark or leaves) but are shared among species.This is highlighted by the fact that 8% of the variability <strong>in</strong> VOC <strong>composition</strong>is expla<strong>in</strong>ed by tissue type only. There are therefore somecompounds that seem to be more expressed <strong>in</strong> one of the two tissues<strong>in</strong>dependently of the species. This is confirmed by the factthat we found 38% of the compounds were found only <strong>in</strong> bark analysisand 10% only <strong>in</strong> leaf analysis. This f<strong>in</strong>d<strong>in</strong>g may be expla<strong>in</strong>ed bythe low host specificity among herbivores of tropical plants (Novotnyet al., 2003): most tropical herbivores and pathogens are ableto feed on multiple tree species and a shared herbivore pressureacross tree species on the same tree part may result <strong>in</strong> a sharedchemical <strong>composition</strong>. More unique compounds are found <strong>in</strong> thebark (38% vs. 10%) and future studies are needed to l<strong>in</strong>k the presenceof these compounds with the associated herbivore and pathogenspecies.3. ConclusionsNumber of compounds15010050In this study, we found that bark of tropical tree species tendhold a dist<strong>in</strong>ct and more diverse blend of <strong>volatile</strong> <strong>terpene</strong>s thanleaves. Future studies should be conducted to precise the effectof <strong>terpene</strong>s diversity on the diversity and density of herbivores.Moreover, we focused only on <strong>volatile</strong> defensive compounds andnon-<strong>volatile</strong>s can also control the distribution of <strong>in</strong>sect herbivores(Meurer-Grimes and Tavakilian, 1997). Such a negative correlation<strong>between</strong> chemical diversity and the number of herbivore specieshas been reported for Nothofagus trees <strong>in</strong> South America andNew Zealand (Lavandero et al., 2009). In that study, non-<strong>volatile</strong>chemical uniqueness and chemical diversity of leaves limited thenumber of species of specialist leaf-feed<strong>in</strong>g <strong>in</strong>sects. A logical nextstep <strong>in</strong> studies of <strong>volatile</strong> defenses <strong>in</strong> tropical plants would thereforebe to l<strong>in</strong>k the <strong>terpene</strong> diversity and <strong>composition</strong> that we reporthere to the associated herbivore guilds and to explore therelationship with other defensive compounds <strong>in</strong> the bark and theleaves of tree species.4. Experimental0mono<strong>terpene</strong>ssesqui<strong>terpene</strong>s4.1. Study sitesFig. 2. Number of mono- and sesqui<strong>terpene</strong>s <strong>in</strong> bark only (black), leaves only (lightgrey) or found <strong>in</strong> both tissues (dark grey).Table 2Non-parametric MANOVA of the effect of tissue and species identity on VOC<strong>composition</strong> of the samples <strong>in</strong>cluded <strong>in</strong> this study. Df <strong>in</strong>dicates the degrees of freedomof the factor, F the value of the statistic, R 2 the percentage of variance expla<strong>in</strong>ed by thefactor and P the P value associated with the factor.Df F R 2 PTissue 1 137.89 0.08 0.001Species 54 14.54 0.46 0.001Tissue species 54 9.99 0.32 0.001Residuals 246 0.14Total 355 1.00This study was conducted at three sites <strong>in</strong> the Neotropical forestof French Guiana: Paracou Research Station (5°15 0 N; 52°55 0 W),Nouragues Research Station (4°05 0 N, 52°40 0 W) and MontagneTortue (4°18 0 N; 52°22 0 W). All three sites are covered with prist<strong>in</strong>etropical ra<strong>in</strong> forest. Annual ra<strong>in</strong>fall is 2990 mm for Nouragues,3160 mm for Paracou and ca. 3500 mm for Montagne Tortue. Infive plots of one hectare each, we sampled all trees with a diameterat breast height (dbh) greater than 10 cm (Baraloto et al., 2012).Two of these plots were at the Paracou site (n = 643 and 487 trees),another two plots were at Nouragues (n = 537 and 567 trees) andone plot at Montagne Tortue (n = 536 trees). Sampl<strong>in</strong>g was conducted<strong>in</strong> June 2007 for Paracou, September 2007 for Nouraguesand November 2007 for Montagne Tortue. Each tree was <strong>in</strong>dividuallyclimbed by a professional tree climber and a voucher specimenwas collected for taxonomic determ<strong>in</strong>ation. Over 98% of the treeswere identified to the species or morphospecies, and the tree speciesrichness ranged <strong>between</strong> 148 and 210 tree species per hectare(Baraloto et al., 2012).4.2. Field sampl<strong>in</strong>g protocolFor 55 species represented by at least two <strong>in</strong>dividuals <strong>in</strong> ourplots, two to five <strong>in</strong>dividuals were selected, total<strong>in</strong>g 178 <strong>in</strong>dividuals(Courtois et al., 2009). For each <strong>in</strong>dividual, we collected a 1–2cm 2 piece of leaf (20-mg), and a piece of bark us<strong>in</strong>g a punchof 1 cm 2 (20–70 mg). For each tree, recently but fully expandedsunny leaves were collected from the next-to-last shoot alongthe sampled branch at the crop (Baraloto et al., 2010). Bark sampleswere collected about 1 m above the ground. Each sample wasimmediately placed <strong>in</strong>to a glass vial (10 mL) sealed with a screwcappedtop conta<strong>in</strong><strong>in</strong>g a Teflon-l<strong>in</strong>ed septum (Varian Instruments,