

The Components of Plant Tissue Culture Media II - Horticultural ...
The Components of Plant Tissue Culture Media II - Horticultural ...
The Components of Plant Tissue Culture Media II - Horticultural ...

You also want an ePaper? Increase the reach of your titles
YUMPU automatically turns print PDFs into web optimized ePapers that Google loves.
130 <strong>The</strong> <strong>Components</strong> <strong>of</strong> <strong>Plant</strong> <strong>Tissue</strong> <strong>Culture</strong> <strong>Media</strong> <strong>II</strong><br />
active cell metabolism. Sucrose is generally required<br />
to be present in addition to IAA before tracheid<br />
elements are differentiated in tissue cultures. <strong>The</strong><br />
number <strong>of</strong> both sieve and xylem elements formed<br />
[and possibly the proportion <strong>of</strong> each kind - Wetmore<br />
and Rier (1963); Rier and Beslow (1967)] depends on<br />
sucrose concentration (Aloni, 1980). In Helianthus<br />
tuberosus tuber slices, although sucrose, glucose and<br />
trehalose were best for supporting cell division and<br />
tracheid formation, maltose was only a moderately<br />
effective carbon source (Minocha and Halperin,<br />
1974). Shininger (1979) has concluded that only<br />
carbohydrates which enable significant cell division<br />
are capable <strong>of</strong> promoting tracheary element<br />
formation. <strong>The</strong> occurrence <strong>of</strong> lignin in cultured cells<br />
is not invariably associated with thickened cell walls.<br />
Sycamore suspension cultures produced large<br />
amounts <strong>of</strong> lignin when grown on a medium with<br />
abnormally high sucrose (more than 6%) and 2,4-D<br />
levels. It was deposited within the cells and released<br />
into the medium (Carceller et al., 1971).<br />
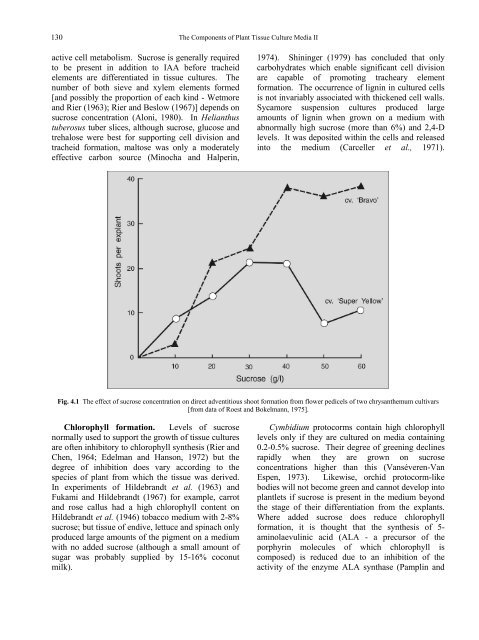
Fig. 4.1 <strong>The</strong> effect <strong>of</strong> sucrose concentration on direct adventitious shoot formation from flower pedicels <strong>of</strong> two chrysanthemum cultivars<br />
[from data <strong>of</strong> Roest and Bokelmann, 1975].<br />
Chlorophyll formation. Levels <strong>of</strong> sucrose<br />
normally used to support the growth <strong>of</strong> tissue cultures<br />
are <strong>of</strong>ten inhibitory to chlorophyll synthesis (Rier and<br />
Chen, 1964; Edelman and Hanson, 1972) but the<br />
degree <strong>of</strong> inhibition does vary according to the<br />
species <strong>of</strong> plant from which the tissue was derived.<br />
In experiments <strong>of</strong> Hildebrandt et al. (1963) and<br />
Fukami and Hildebrandt (1967) for example, carrot<br />
and rose callus had a high chlorophyll content on<br />
Hildebrandt et al. (1946) tobacco medium with 2-8%<br />
sucrose; but tissue <strong>of</strong> endive, lettuce and spinach only<br />
produced large amounts <strong>of</strong> the pigment on a medium<br />
with no added sucrose (although a small amount <strong>of</strong><br />
sugar was probably supplied by 15-16% coconut<br />
milk).<br />
Cymbidium protocorms contain high chlorophyll<br />
levels only if they are cultured on media containing<br />
0.2-0.5% sucrose. <strong>The</strong>ir degree <strong>of</strong> greening declines<br />
rapidly when they are grown on sucrose<br />
concentrations higher than this (Vanséveren-Van<br />
Espen, 1973). Likewise, orchid protocorm-like<br />
bodies will not become green and cannot develop into<br />
plantlets if sucrose is present in the medium beyond<br />
the stage <strong>of</strong> their differentiation from the explants.<br />
Where added sucrose does reduce chlorophyll<br />
formation, it is thought that the synthesis <strong>of</strong> 5aminolaevulinic<br />
acid (ALA - a precursor <strong>of</strong> the<br />
porphyrin molecules <strong>of</strong> which chlorophyll is<br />
composed) is reduced due to an inhibition <strong>of</strong> the<br />
activity <strong>of</strong> the enzyme ALA synthase (Pamplin and



