

under different light regimes - Biology Department
under different light regimes - Biology Department
under different light regimes - Biology Department

Create successful ePaper yourself
Turn your PDF publications into a flip-book with our unique Google optimized e-Paper software.
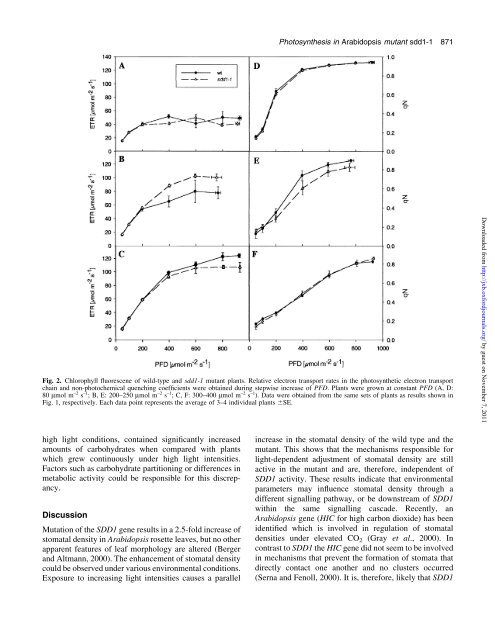
Fig. 2. Chlorophyll ¯uorescene of wild-type and sdd1-1 mutant plants. Relative electron transport rates in the photosynthetic electron transport<br />
chain and non-photochemical quenching coef®cients were obtained during stepwise increase of PFD. Plants were grown at constant PFD (A, D:<br />
80 mmol m ±2 s ±1 ; B, E: 200±250 mmol m ±2 s ±1 ; C, F: 300±400 mmol m ±2 s ±1 ). Data were obtained from the same sets of plants as results shown in<br />
Fig. 1, respectively. Each data point represents the average of 3±4 individual plants 6SE.<br />
high <strong>light</strong> conditions, contained signi®cantly increased<br />
amounts of carbohydrates when compared with plants<br />
which grew continuously <strong>under</strong> high <strong>light</strong> intensities.<br />
Factors such as carbohydrate partitioning or differences in<br />
metabolic activity could be responsible for this discrepancy.<br />
Discussion<br />
Mutation of the SDD1 gene results in a 2.5-fold increase of<br />
stomatal density in Arabidopsis rosette leaves, but no other<br />
apparent features of leaf morphology are altered (Berger<br />
and Altmann, 2000). The enhancement of stomatal density<br />
could be observed <strong>under</strong> various environmental conditions.<br />
Exposure to increasing <strong>light</strong> intensities causes a parallel<br />
Photosynthesis in Arabidopsis mutant sdd1-1 871<br />
increase in the stomatal density of the wild type and the<br />
mutant. This shows that the mechanisms responsible for<br />
<strong>light</strong>-dependent adjustment of stomatal density are still<br />
active in the mutant and are, therefore, independent of<br />
SDD1 activity. These results indicate that environmental<br />
parameters may in¯uence stomatal density through a<br />
<strong>different</strong> signalling pathway, or be downstream of SDD1<br />
within the same signalling cascade. Recently, an<br />
Arabidopsis gene (HIC for high carbon dioxide) has been<br />
identi®ed which is involved in regulation of stomatal<br />
densities <strong>under</strong> elevated CO2 (Gray et al., 2000). In<br />
contrast to SDD1 the HIC gene did not seem to be involved<br />
in mechanisms that prevent the formation of stomata that<br />
directly contact one another and no clusters occurred<br />
(Serna and Fenoll, 2000). It is, therefore, likely that SDD1<br />
Downloaded from<br />
http://jxb.oxfordjournals.org/ by guest on November 7, 2011

