


Guía de Trabajos Prácticos 2012. Primer Cuatrimestre
Guía de Trabajos Prácticos 2012. Primer Cuatrimestre
Guía de Trabajos Prácticos 2012. Primer Cuatrimestre

You also want an ePaper? Increase the reach of your titles
YUMPU automatically turns print PDFs into web optimized ePapers that Google loves.
<strong>Guía</strong> <strong>de</strong> <strong>Trabajos</strong> <strong>Prácticos</strong><br />
Departamento Ecología, Genética y Evolución<br />
1er. <strong>Cuatrimestre</strong> 2012
.<br />
DOCENTES<br />
ECOLOGÍA GENERAL – <strong>Primer</strong> <strong>Cuatrimestre</strong> 2012<br />
http://www.ege.fcen.uba.ar/materias/general/general.htm<br />
Nombre E-mail<br />
Prof. Cécere, Carla carla@ege.fcen.uba.ar<br />
Prof. Suárez, Olga osuarez@ege.fcen.uba.ar<br />
JTP Fischer, Sylvia sylvia@ege.fcen.uba.ar<br />
JTP Thompson, Gustavo gustavo@ege.fcen.uba.ar<br />
JTP Rubel, Diana dianaru@ege.fcen.uba.ar<br />
Ay 1° Paris, Carolina baikibadai@yahoo.com<br />
Ay 1° Sinistro, Rodrigo rs@ege.fcen.uba.ar<br />
Ay 1° Loetti, Verónica laloetti@hotmail.com<br />
Ay 1° Tuero Diego dttuero@ege.fcen.uba.ar<br />
Ay 2° Alvarez Costa, Agustin agusalvarezcosta@hotmail.com<br />
Ay. 2° Petino Zappala, M. Alejandra mapz@ege.fcen.uba.ar<br />
Ay. 2° Franco Tadic, Luis franco-luis@hotmail.com<br />
Lista <strong>de</strong> Alumnos EGE: es una lista <strong>de</strong> distribución <strong>de</strong> información para alumnos <strong>de</strong>l<br />
Departamento <strong>de</strong> Ecología, Genética y Evolución, acerca <strong>de</strong> becas, cursos y otras<br />
cuestiones <strong>de</strong> interés. No es una lista <strong>de</strong> discusión. Pue<strong>de</strong> Ud. suscribirse a la lista en:<br />
http://www.ege.fcen.uba.ar/mailman/listinfo/alumnos<br />
o accediendo <strong>de</strong>s<strong>de</strong> el link en la página <strong>de</strong>l Departamento.<br />
También en esta página encontrará los correos electrónicos <strong>de</strong> los representantes<br />
estudiantiles <strong>de</strong>l EGE, para realizar cualquier consulta.<br />
REGIMEN DE APROBACIÓN DEL CURSO<br />
Ecología General<br />
Los requisitos para aprobar los trabajos prácticos son:<br />
(1) Aprobar 2 exámenes parciales con un mínimo <strong>de</strong> 60 puntos. Se podrán recuperar los 2<br />
exámenes parciales. La fecha <strong>de</strong> recuperación será a posteriori <strong>de</strong>l segundo examen<br />
parcial. A los fines <strong>de</strong> calcular la nota final <strong>de</strong> la materia se consi<strong>de</strong>rarán las notas <strong>de</strong> todos<br />
los exámenes.<br />
(2) Aprobar el 80% <strong>de</strong> los informes <strong>de</strong> los trabajos prácticos. El informe <strong>de</strong>berá entregarse la<br />
semana siguiente a la finalización <strong>de</strong>l trabajo práctico. En caso <strong>de</strong> no ser aceptado será<br />
2
Ecología General<br />
<strong>de</strong>vuelto para su corrección y nueva entrega. En caso <strong>de</strong> no ser aceptado luego <strong>de</strong> la<br />
segunda corrección se consi<strong>de</strong>rará <strong>de</strong>saprobado.<br />
(3) Asistir al 80% <strong>de</strong> los trabajos prácticos. Se tomará asistencia al comienzo <strong>de</strong>l trabajo<br />
práctico. Los alumnos que lleguen 10 minutos <strong>de</strong>spués <strong>de</strong> iniciado el mismo tendrán media<br />
inasistencia y los que lleguen luego <strong>de</strong> 20 minutos tendrán ausente.<br />
Los requisitos para aprobar la materia por promoción sin dar examen final son:<br />
(1) Aprobar los 2 exámenes parciales (sin la opción <strong>de</strong>l recuperatorio) con un mínimo <strong>de</strong> 70<br />
puntos cada uno y tener un promedio mínimo en los parciales <strong>de</strong> 80 puntos.<br />
(2) Aprobar todos los informes <strong>de</strong> los trabajos prácticos.<br />
(3) Tener los finales aprobados <strong>de</strong> las materias correlativas.<br />
La nota final para aquellos alumnos que hayan promovido será elaborada en base al<br />
promedio <strong>de</strong> los 2 exámenes parciales, los informes <strong>de</strong> laboratorio y el <strong>de</strong>sempeño en<br />
<strong>Trabajos</strong> <strong>Prácticos</strong>.<br />
Aquellos alumnos que hayan aprobado los trabajos prácticos pero que no hayan promovido<br />
<strong>de</strong>berán dar un examen final escrito, cuya nota <strong>de</strong> aprobación es <strong>de</strong> 60/100. La nota final<br />
será calculada como un promedio pon<strong>de</strong>rado entre la nota <strong>de</strong>l examen final (60%) y <strong>de</strong> la<br />
nota <strong>de</strong> <strong>Trabajos</strong> <strong>Prácticos</strong> (40%).<br />
PROGRAMA ANALÍTICO<br />
INTRODUCCIÓN A LA ECOLOGÍA<br />
¿Qué es ecología? Niveles <strong>de</strong> organización. Método científico en ecología. Nociones generales <strong>de</strong><br />
biología evolutiva. Métodos <strong>de</strong> muestreo y diseño <strong>de</strong> experimentos en ecología. Escalas espaciales y<br />
temporales. Problemas ecológicos actuales.<br />
FACTORES QUE LIMITAN LA DISTRIBUCIÓN DE LOS ORGANISMOS<br />
Recursos y condiciones. Temperatura. Salinidad. Radiación. CO 2 . H 2 O. Nutrientes. Espacio.<br />
Ectotermos y endotermos. Nicho ecológico. Aclimatación, migración, almacenamiento y letargo.<br />
Principales recursos para plantas y animales. Generalistas, especialistas, oportunistas y selectivos.<br />
Biomas.<br />
POBLACIONES<br />
Concepto <strong>de</strong> población. Atributos poblacionales. Composición <strong>de</strong> la población. Abundancia y rango <strong>de</strong><br />
distribución, tamaño corporal y latitud. Densidad absoluta y relativa e índices <strong>de</strong> <strong>de</strong>nsidad. Censos.<br />
Curvas poblacionales. Métodos basados en marcado y recaptura y en la reducción <strong>de</strong>l tamaño<br />
poblacional. Disposición espacial: al azar, regular y contagiosa. Distribución <strong>de</strong> Poisson y Binomial<br />
negativa.<br />
Demografía. Estadística vital. Tablas <strong>de</strong> vida y <strong>de</strong> fecundidad. Curvas <strong>de</strong> supervivencia. Tasas <strong>de</strong><br />
reproducción, tiempo generacional y tasas <strong>de</strong> incremento. Distribución <strong>de</strong> eda<strong>de</strong>s. Valor reproductivo.<br />
Poblaciones con generaciones discretas y con solapamiento. Historias <strong>de</strong> vida. Plasticidad fenotípica.<br />
Esfuerzo reproductivo. Edad <strong>de</strong> la primera reproducción. Iteroparidad y semelparidad. Tamaño y<br />
número <strong>de</strong> crías. Senescencia.<br />
Dinámica poblacional. Denso<strong>de</strong>pen<strong>de</strong>ncia y <strong>de</strong>nsoin<strong>de</strong>pen<strong>de</strong>ncia. Competencia intraespecífica.<br />
Curvas exponencial y logística: teoría y ejemplos <strong>de</strong> poblaciones naturales y <strong>de</strong> laboratorio. Mo<strong>de</strong>los<br />
que incorporan un retraso temporal. Regulación poblacional. Demografía humana.<br />
Relaciones interespecíficas. Distintos tipos. Competencia interespecífica. Mo<strong>de</strong>lo <strong>de</strong> Lotka y Volterra.<br />
Concepto <strong>de</strong> nicho y principio <strong>de</strong> exclusión competitiva. Efectos <strong>de</strong> los predadores sobre la población<br />
<strong>de</strong> presas. Ciclos predador-presa: hipótesis sobre sus causas. Mo<strong>de</strong>lo <strong>de</strong> Lotka-Volterra y <strong>de</strong>rivados.<br />
3
Ecología General<br />
Parasitismo: Micro y macroparásitos. Infección y enfermedad. Transmisión y distribución. Efecto <strong>de</strong>l<br />
parasitismo sobre el hospedador individual y su población. Herbivoría. Relaciones positivas entre<br />
especies: comensalismo, simbiosis. Coevolución.<br />
ESTRUCTURA Y DESARROLLO DE LA COMUNIDAD<br />
Características <strong>de</strong> la comunidad. Clasificación y or<strong>de</strong>nación <strong>de</strong> las comunida<strong>de</strong>s. Descripción <strong>de</strong> la<br />
composición <strong>de</strong> la comunidad. Indices <strong>de</strong> diversidad. Análisis <strong>de</strong> gradientes. Comunidad clímax.<br />
Mecanismos <strong>de</strong>l proceso <strong>de</strong> sucesión.<br />
Organización <strong>de</strong> la comunidad. Influencia <strong>de</strong> la competencia y predación en la estructura <strong>de</strong> la<br />
comunidad. Ca<strong>de</strong>nas alimenticias y niveles tróficos. Especies principales y especies dominantes.<br />
Control "top-down" y "bottom-up" <strong>de</strong> las tramas tróficas. Gremios. Estabilidad <strong>de</strong> la comunidad.<br />
Dinámica temporal <strong>de</strong> las comunida<strong>de</strong>s: concepto <strong>de</strong> sucesión. Sucesión primaria y secundaria. Tipos<br />
<strong>de</strong> sucesión.<br />
Determinantes <strong>de</strong> la biodiversidad. Efectos <strong>de</strong>l clima, heterogeneidad espacial y temporal,<br />
perturbaciones, productividad.<br />
FLUJO DE ENERGÍA Y MATERIA A TRAVÉS DEL ECOSISTEMA<br />
Flujo <strong>de</strong> energía y materia a través <strong>de</strong>l ecosistema. Re<strong>de</strong>s y ca<strong>de</strong>nas tróficas. Productividad primaria.<br />
Productividad secundaria. Eficiencias <strong>de</strong> transferencia <strong>de</strong> energía entre niveles tróficos. ¿Qué limita el<br />
número <strong>de</strong> niveles tróficos? Factores que limitan la productividad primaria en ecosistemas terrestres y<br />
acuáticos. Factores que limitan la productividad secundaria en ecosistemas terrestres y acuáticos.<br />
Ciclos biogeoquímicos. Alteraciones <strong>de</strong> los principales ciclos biogeoquímicos.<br />
ECOLOGÍA DE PAISAJES Y REGIONES<br />
Desarrollo histórico. Conceptos <strong>de</strong> paisaje, región y ecosistema local. Mo<strong>de</strong>lo <strong>de</strong> parche-corredormatriz.<br />
Mosaicos y gradientes. Patrones espaciales. Teoría jerárquica.<br />
APLICACIONES DE LA ECOLOGÍA DE POBLACIONES<br />
Manejo y explotación <strong>de</strong> recursos naturales. Rendimientos máximo sostenible. Mo<strong>de</strong>los <strong>de</strong><br />
explotación. Rendimiento económico óptimo. Declinación <strong>de</strong> la abundancia <strong>de</strong> ballenas y otros<br />
”stocks” pesqueros. Control <strong>de</strong> plagas y malezas: control biológico, cultural, genético y químico.<br />
Manejo integrado <strong>de</strong> plagas. Pesticidas: efectos adversos y positivos sobre la plaga y otros<br />
organismos. Nivel <strong>de</strong> daño económico y <strong>de</strong> umbral <strong>de</strong> acciones.<br />
BIODIVERSIDAD Y CONSERVACIÓN<br />
Concepto <strong>de</strong> biodiversidad. Valor intrínseco y utilitario <strong>de</strong> la biodiversidad. ¿Cuántas especies<br />
existen? Patrones geográficos <strong>de</strong> distribución <strong>de</strong> especies. Relaciones especies-area. Biogeografía<br />
<strong>de</strong> islas y mo<strong>de</strong>lo <strong>de</strong>l equilibrio. Biodiversidad y estabilidad <strong>de</strong> los ecosistemas. Tasas <strong>de</strong> extinción<br />
históricas y recientes. Principales causas <strong>de</strong> extinciones recientes. Poblaciones viables mínimas.<br />
Conservación <strong>de</strong> especies amenazadas. Fragmentación <strong>de</strong>l hábitat y efecto <strong>de</strong> bor<strong>de</strong>. Diseño <strong>de</strong><br />
reservas.<br />
CONTAMINACIÓN EN ECOSISTEMAS ACUÁTICOS Y TERRESTRES<br />
Tipos principales <strong>de</strong> contaminantes en el ambiente: orígenes y fuentes <strong>de</strong> emisión, ingreso y dinámica<br />
en el ambiente. Niveles ecológicos <strong>de</strong> acción. Bioconcentración y biomagnificación. Evaluación y<br />
diagnóstico <strong>de</strong> la contaminación: parámetros físicos y químicos <strong>de</strong> referencia. Bioindicadores.<br />
Respuesta <strong>de</strong> la biota al estrés ambiental. Indices ecológicos para cuantificar el <strong>de</strong>terioro ambiental.<br />
Bioensayos.<br />
Bibliografía<br />
Begon M, Harper JL y Townsend CR (1996) Ecology: individuals, populations and communities.<br />
Blackwell Sci., Oxford (Versión en español <strong>de</strong> la 2da. edición inglesa: (1990), Ed. Omega,<br />
Barcelona).<br />
Caughley G (1977) Analysis of vertebrate populations. Wiley, New York.<br />
Dobson AP (1996) Conservation and biodiversity. Scientific American Library, New York.<br />
4
Ecología General<br />
Forman RTT (1995) Land mosaics. The ecology of landscapes and regions. Cambridge Univ. Press,<br />
Cambridge.<br />
Krebs CJ (1989) Ecological methodology. Harper Collins, New York.<br />
Krebs CJ (1994) Ecology: the experimental analysis of distribution and abundance. Harper Collins,<br />
New York (Versión en español <strong>de</strong> la 3ra. edición inglesa: (1985), Ed. Pirámi<strong>de</strong>, Madrid).<br />
Rabinovich JR (1980) Introducción a la ecología <strong>de</strong> las poblaciones animales. CECSA, Caracas.<br />
Ricklefs RE (1997) The economy of nature. W. Freeman & Co., New York (Versión en español:<br />
Invitación a la ecología. Ed. Médica Panamericana, Buenos Aires).<br />
Smith, R. & Smith, T (2001) Ecología. 4ta. edición. Addison – Wesley. Madrid.<br />
Stiling PD (1996) Ecology: theory and applications. Prentice Hall, New Jersey.<br />
Townsend CR, Harper JL y Begon M (2000) Essentials of ecology. Blackwell Sci., Oxford.*<br />
5
Ecología General<br />
ECOLOGIA GENERAL 2012 (1° <strong>Cuatrimestre</strong>). Cronograma clases Teóricas y <strong>Trabajos</strong> <strong>Prácticos</strong><br />
Cronograma <strong>de</strong> uso <strong>de</strong> instalaciones<br />
Días en que se va a usar el laboratorio<br />
29/3, 8/5, 24/5, 29/5, 31/5<br />
Días en que se va a ingresar a uno <strong>de</strong> los laboratorios durante 15 minutos para contar lemnas:<br />
3/4, 10/4, 17/4, 24/4<br />
Días en que se va a usar aula <strong>de</strong> seminario<br />
3/4, 17/4, 15/5, 22/5, 5/6, 21/6, 26/6, 28/6<br />
Días en que se van a usar computadoras<br />
19/4, 24/4, 26/4, 3/5, 8/5, 10/5, 7/5, 12/5, 14/5<br />
DIA FECHA CLASES TEÓRICAS<br />
Niveles <strong>de</strong> organización.<br />
Escalas espaciales y<br />
TRABAJOS PRÁCTICOS<br />
Martes 20 MARZO<br />
temporales en ecología.<br />
Areas <strong>de</strong> la ecología.<br />
Problemas ecológicos<br />
actuales<br />
Jueves 22 MARZO<br />
Martes 27 MARZO<br />
Jueves 29 MARZO<br />
Martes 3 ABRIL<br />
JUEVES 5 ABRIL<br />
Martes 10 ABRIL<br />
Recursos y condiciones.<br />
Nicho ecológico<br />
Efectos <strong>de</strong> las<br />
condiciones y recursos a<br />
nivel individual y<br />
poblacional.<br />
Adaptaciones. Ecología<br />
evolutiva<br />
Recursos. Tipos <strong>de</strong><br />
recursos. Tipos <strong>de</strong><br />
organismos según uso <strong>de</strong><br />
recursos<br />
Poblaciones. Atributos.<br />
Abundancia, estructura<br />
espacial y temporal.<br />
Métodos <strong>de</strong> estimación<br />
<strong>de</strong> abundancia<br />
poblacional y<br />
disposición espacial.<br />
FERIADO SEMANA<br />
SANTA<br />
Variaciones <strong>de</strong> las<br />
poblaciones en el<br />
tiempo.<br />
Ciclos <strong>de</strong> vida.<br />
(LABO)<br />
- Inscripción.<br />
- Explicación TP Recursos y<br />
Condiciones e inicio <strong>de</strong> trabajo<br />
experimental con Lemnas<br />
- Inicio TP Descomposición Foliar: peso<br />
fresco experimental (campo) y testigo<br />
(estufa), armado dispositivos +<br />
colocación en campo experimental).<br />
(LABO LEMNAS + AULA)-. Recuento<br />
<strong>de</strong> Lemnas (Semana 1).<br />
- Explicación TPs: Abundancia<br />
- Muestreo <strong>de</strong> vegetación y animales +<br />
<strong>de</strong> salida <strong>de</strong> campo<br />
- Pesada muestras TP <strong>de</strong>scomposición<br />
foliar (peso seco testigo)<br />
FERIADO SEMANA SANTA<br />
(LABO LEMNAS) - Salida <strong>de</strong> campo:<br />
Muestreo <strong>de</strong> poblaciones animales<br />
(Turno mañana)<br />
- Turno noche: libre x sabado<br />
6
Jueves 12 ABRIL<br />
Sábado 14 ABRIL<br />
Martes 17 ABRIL<br />
Jueves 19 ABRIL<br />
Martes 24 ABRIL<br />
Jueves 26 ABRIL<br />
Tablas <strong>de</strong> vida.<br />
Parámetros <strong>de</strong><br />
supervivencia y<br />
reproducción.<br />
Sin teórica por muestreo<br />
en RECS<br />
Dinámica poblacional.<br />
Mo<strong>de</strong>los <strong>de</strong> crecimiento:<br />
exponencial, logístico,<br />
con y sin estructura <strong>de</strong><br />
eda<strong>de</strong>s, <strong>de</strong>terminísticos y<br />
estocásticos.<br />
Mo<strong>de</strong>los <strong>de</strong> crecimiento<br />
con tiempo <strong>de</strong> retardo.<br />
Competencia<br />
intraespecífica.<br />
Distintos tipos <strong>de</strong><br />
historia <strong>de</strong> vida.<br />
Relaciones<br />
interespecíficas.<br />
Competencia.<br />
Mo<strong>de</strong>lo <strong>de</strong> Lotka<br />
Volterra. Mo<strong>de</strong>los <strong>de</strong><br />
Tilman.<br />
Formas <strong>de</strong> estimar la<br />
competencia<br />
interespecífica<br />
Predación,<br />
Respuesta numérica.<br />
Rta funcional<br />
Mo<strong>de</strong>lo LV.<br />
Martes 1 MAYO Feriado Feriado<br />
Jueves 3 MAYO<br />
Martes 8 MAYO<br />
Jueves 10 MAYO<br />
Modificaciones al<br />
mo<strong>de</strong>lo LV <strong>de</strong><br />
predación. Efectos <strong>de</strong> la<br />
<strong>de</strong>nso<strong>de</strong>pen<strong>de</strong>ncia, los<br />
refugios y la respuesta<br />
funcional. Aplicaciones<br />
<strong>de</strong> la ecología <strong>de</strong><br />
poblaciones (cosecha).<br />
Explotación <strong>de</strong> recursos<br />
y control <strong>de</strong> plagas.<br />
Definición <strong>de</strong><br />
comunida<strong>de</strong>s. Enfoques.<br />
Atributos<br />
- Recuento <strong>de</strong> lemnas (Semana 2)<br />
Ecología General<br />
- Salida <strong>de</strong> campo: Muestreo <strong>de</strong><br />
vegetación (Turno mañana)<br />
- Salida <strong>de</strong> campo: Muestreo <strong>de</strong><br />
poblaciones animales (Turno noche X<br />
LA MAÑANA)<br />
- Salida <strong>de</strong> campo: Muestreo <strong>de</strong><br />
vegetación (Turno noche)<br />
(LABO LEMNAS + AULA) - Recuento<br />
<strong>de</strong> lemnas (Semana 3)<br />
- Tablas <strong>de</strong> Vida I<br />
(COMPU)<br />
- Tablas <strong>de</strong> Vida II<br />
-. TP muestreo <strong>de</strong> poblaciones animales:<br />
Armado <strong>de</strong> base <strong>de</strong> datos + entrega <strong>de</strong> la<br />
base <strong>de</strong> datos<br />
(LABO LEMNAS + COMPU)- Fin<br />
recuento <strong>de</strong> lemnas (Semana 4)<br />
- Análisis, Discusión y cierre TP:<br />
Muestreo <strong>de</strong> vegetación.<br />
(COMPU)<br />
– Discusión <strong>de</strong> problemas con la carga<br />
<strong>de</strong> datos, cuentas y cierre TP muestreo<br />
poblaciones animales<br />
(COMPU)<br />
- Mo<strong>de</strong>los <strong>de</strong> Crecimiento Poblacional<br />
-Cálculos /discusión/cierre <strong>de</strong>l<br />
experimento con Lemnas (parte <strong>de</strong><br />
recursos y condiciones)<br />
(LABO + COMPU) - Cálculos,<br />
discusión y cierre <strong>de</strong>l experimento <strong>de</strong><br />
competencia entre lemnas<br />
- Predación – RF (parte experimental)<br />
(COMPU)<br />
- Cierre Predación-RF.<br />
7
Martes 15 MAYO<br />
REPASO TEÓRICAS<br />
1°PARTE<br />
(AULA)<br />
- REPASO PRACTICOS 1° PARTE<br />
Jueves 17 MAYO PRIMER PARCIAL PRIMER PARCIAL<br />
Martes 22 MAYO<br />
Jueves 24 MAYO<br />
Martes 29 MAYO<br />
Jueves 31 MAYO<br />
Mo<strong>de</strong>los especieabundancia.<br />
Descripción<br />
y comparación <strong>de</strong><br />
comunida<strong>de</strong>s.<br />
Clasificación y<br />
or<strong>de</strong>namiento<br />
Organización <strong>de</strong><br />
comunida<strong>de</strong>s: factores<br />
que estructuran<br />
comunida<strong>de</strong>s.<br />
Gremio y competencia.<br />
Predación Disturbios<br />
Dinámica temporal y<br />
espacial. Sucesión.<br />
Invasiones.<br />
Biogeografía <strong>de</strong> islas.<br />
Patrones <strong>de</strong> diversidad<br />
Patrones espaciales.<br />
Macro ecología.<br />
Ecología <strong>de</strong>l paisaje y<br />
regional<br />
Martes 5 JUNIO Conservación.<br />
Jueves<br />
7 JUNIO<br />
Martes 12 JUNIO<br />
Jueves 14 JUNIO<br />
Sábado 16 JUNIO<br />
Martes<br />
19 JUNIO<br />
Ecosistemas.<br />
Definiciones y<br />
conceptos.<br />
Flujo <strong>de</strong> materia y<br />
energía. Eficiencias<br />
ecológicas<br />
Producción primaria.<br />
Determinantes <strong>de</strong> la<br />
producción primaria<br />
Ciclos biogeoquímicos.<br />
Ciclos <strong>de</strong>l agua, el<br />
oxígeno, papel <strong>de</strong>l<br />
ozono.<br />
Agujero <strong>de</strong> ozono.<br />
Ciclos <strong>de</strong>l Fósforo y<br />
azufre<br />
Salida a Otamendi<br />
Ecología General<br />
(LABO)<br />
- Diversidad <strong>de</strong> artrópodos: Explicación<br />
y colocación <strong>de</strong> trampas pitfall en<br />
Ciudad Universitaria<br />
- Atributos <strong>de</strong> las Comunida<strong>de</strong>s (inicio)<br />
(AULA)<br />
- Atributos <strong>de</strong> las Comunida<strong>de</strong>s (fin)<br />
(LABO)<br />
- Retirar muestras campo experimental<br />
TP <strong>de</strong>scomposición foliar, colocar hojas<br />
en estufa.<br />
- Retirar muestras trampas pitfall e<br />
iniciar i<strong>de</strong>ntificación <strong>de</strong> artrópodos.<br />
(LABO)<br />
- I<strong>de</strong>ntificación <strong>de</strong> artrópodos<br />
-Retirar muestras <strong>de</strong>scomposición foliar<br />
<strong>de</strong> estufa y pesar (peso seco<br />
experimental).<br />
(AULA)<br />
-TP Invasiones Biológicas: Seminario<br />
(COMPU)<br />
-Cierre TP Diversidad <strong>de</strong> artrópodos<br />
(COMPU)<br />
-Explicación TP <strong>de</strong>scomposición foliar,<br />
analisis <strong>de</strong> resultados y cierre.<br />
(COMPU)<br />
-Ecología <strong>de</strong>l Paisaje: Explicación salida<br />
<strong>de</strong> campo y mapas (primera parte-<br />
GoogleEarth) -Cierre GoogleEarth y<br />
mapas<br />
-Ecología <strong>de</strong>l Paisaje: Salida <strong>de</strong> campo<br />
(turno noche)<br />
-Ecología <strong>de</strong>l Paisaje: Salida <strong>de</strong> campo<br />
(turno mañana). (turno noche libre)<br />
8
Jueves 21 Junio<br />
Ciclos <strong>de</strong>l carbono y<br />
nitrógeno. Efectos<br />
humanos.<br />
Cambio climático.<br />
Martes 26 JUNIO Contaminación. Película<br />
Jueves 28 JUNIO REPASO TEÓRICAS<br />
Martes 3 JULIO SEGUNDO PARCIAL<br />
Jueves 5 JULIO Recuperatorio 1<br />
Viernes 6 JULIO<br />
Entrega notas 2do<br />
parcial<br />
Jueves 12 JULIO Recuperatorio 2<br />
AULA<br />
- Cierre TP Ecología <strong>de</strong>l Paisaje<br />
(AULA)<br />
Cambio Climático<br />
(AULA)<br />
REPASO PRÁCTICOS 2°PARTE<br />
Ecología General<br />
9
Seguridad en laboratorios <strong>de</strong> docencia (<br />
Ecología General<br />
10
Ecología General<br />
11
Ecología General<br />
12
Ecología General<br />
13
Ecología General<br />
14
Ecología General<br />
15
ÍNDICE GENERAL DE TRABAJOS PRÀCTICOS<br />
Ecología General<br />
¿CÓMO ESCRIBIR UN TRABAJO CIENTÍFICO?<br />
1. PRINCIPIOS DE DISEÑO EXPERIMENTAL EN ECOLOGÍA.<br />
2. RECURSOS Y CONDICIONES<br />
(1) Introducción<br />
(2) <strong>Guía</strong> <strong>de</strong> trabajo experimental (laboratorio)<br />
3. ESTIMACIÓN DE ABUNDANCIA<br />
(1) Conceptos generales sobre estimación <strong>de</strong> abundancia. Muestreo.<br />
(2) Métodos para la estimación <strong>de</strong> la abundancia <strong>de</strong> vegetación herbácea y arbórea<br />
(3) Planillas salida <strong>de</strong> campo<br />
4. ESTIMACIÓN DE LA ABUNDANCIA EN POBLACIONES ANIMALES<br />
(1) Introducción<br />
(2) Salida a plazas <strong>de</strong> la Ciudad <strong>de</strong> Buenos Aires.<br />
(3) Mapas<br />
(4) Planillas<br />
(5) Seminario: Walker RS, Pancotto V, Schachter-Broi<strong>de</strong> J, Ackermann G, Novaro AJ. 2000.<br />
Evaluation of a fecal-pellet in<strong>de</strong>x for abundance for mountain vizcachas (Lagidium<br />
viscacia) in Patagonia. Mastozoología Neotropical (SAREM) 7(2):89-94<br />
5. TABLAS DE VIDA<br />
(1) Métodos para la estimación <strong>de</strong> la edad<br />
(2) Tablas <strong>de</strong> vida<br />
(3) Aplicaciones <strong>de</strong> tablas <strong>de</strong> vida<br />
6. MODELOS DE CRECIMIENTO POBLACIONAL<br />
(1) Introducción<br />
(2) Activida<strong>de</strong>s<br />
7. COMPETENCIA.<br />
8. PREDACIÓN. RESPUESTA FUNCIONAL.<br />
9. COMPARACION DE LAS TASAS DE DESCOMPOSICION FOLIAR DE DISTINTAS ESPECIES<br />
VEGETALES<br />
(1) Introducción<br />
(2) <strong>Guía</strong> <strong>de</strong> trabajo experimental a realizar en Ciudad Universitaria.<br />
10. ATRIBUTOS DE LAS COMUNIDADES<br />
(1) Introducción<br />
(2) <strong>Guía</strong> <strong>de</strong> trabajo<br />
11. COMPARACIÓN DE LA COMPOSICIÓN DE LA COMUNIDAD DE INVERTEBRADOS<br />
CAMINADORES EN DOS AMBIENTES DIFERENTES DE CIUDAD UNIVERSITARIA<br />
(1) Introducción<br />
(2) <strong>Guía</strong> <strong>de</strong> trabajo experimental en Ciudad Universitaria.<br />
12. INVASIONES BIOLÓGICAS: Seminarios<br />
Zavaleta ES, Hobbs RJ, Mooney HA. 2001. Viewing invasive species removal in a wholeecosystem<br />
context. Trends in Ecology and Evolution, 16 (8): 454-459<br />
Shea K, Chesson P. 2002. Community ecology theory as a framework for biological invasions.<br />
Trends in Ecology and Evolution, 17(4): 170-176.<br />
13. ECOLOGIA DEL PAISAJE.<br />
14. CAMBIO CLIMATICO<br />
16
¿CÓMO ESCRIBIR UN TRABAJO CIENTÍFICO?<br />
(1) GUÍA PARA LA REDACCIÓN DE INFORMES<br />
Agra<strong>de</strong>cemos a la Cátedra <strong>de</strong> Limnología por permitirnos su reproducción.<br />
Así como los trabajos prácticos <strong>de</strong> las<br />
diferentes materias tienen por finalidad<br />
entrenar al estudiante en las tareas a las que<br />
se <strong>de</strong>dicará una vez recibido, los informes son<br />
ensayos <strong>de</strong> los manuscritos que los alumnos<br />
presentarán en revistas científicas en el futuro<br />
próximo.<br />
Contener buena información es el requisito<br />
más importante <strong>de</strong> un trabajo <strong>de</strong> investigación<br />
que se preten<strong>de</strong> publicar, pero <strong>de</strong> ninguna<br />
manera es el único. El lenguaje, la hilvanación<br />
<strong>de</strong> los razonamientos expuestos, la claridad, la<br />
presentación, la calidad <strong>de</strong> las ilustraciones,<br />
etc., son aspectos <strong>de</strong> gran importancia que<br />
<strong>de</strong>cidirán si el trabajo es aceptado o no.<br />
Estas facetas no son banales ni secundarias.<br />
No solamente <strong>de</strong>terminan el potencial <strong>de</strong><br />
publicación, sino también condicionan la<br />
accesibilidad <strong>de</strong> la información expuesta a los<br />
futuros lectores. Un trabajo tedioso, reiterativo,<br />
mal compuesto y mal escrito, con figuras<br />
confusas y <strong>de</strong> mala calidad, aún cuando se<br />
publique pue<strong>de</strong> no ser leído y/o comprendido<br />
jamás más que por su propio autor. Los<br />
norteamericanos, expertos en publicar o<br />
perecer, han acuñado la expresión rea<strong>de</strong>r<br />
friendly, es <strong>de</strong>cir "amistoso con el lector". Los<br />
editores <strong>de</strong> las revistas exigen que, a<strong>de</strong>más<br />
<strong>de</strong> contener información suficiente en calidad y<br />
cantidad, el manuscrito sea rea<strong>de</strong>r friendly.<br />
Esto significa que el lector, aún aquél que no<br />
está estrechamente familiarizado con el tema,<br />
pueda compren<strong>de</strong>r rápidamente <strong>de</strong> qué habla<br />
el trabajo, qué material estudió y cómo, y qué<br />
preten<strong>de</strong> <strong>de</strong>mostrar. Para ello, entre otras<br />
cosas, no tendrá que haber repetición <strong>de</strong> la<br />
misma información en diferentes lugares <strong>de</strong>l<br />
texto ni <strong>de</strong> las ilustraciones o tablas. Los<br />
razonamientos expuestos <strong>de</strong>berán estar<br />
fluidamente enca<strong>de</strong>nados entre sí. Las figuras<br />
serán ilustrativas, claras y suficientemente<br />
sencillas como para ser interpretadas <strong>de</strong> un<br />
vistazo. Las conclusiones estarán<br />
efectivamente basadas sobre las evi<strong>de</strong>ncias<br />
presentadas. La bibliografía no tendrá<br />
omisiones, inconsistencias ni errores. En fin,<br />
todos los aspectos estéticos, lingüísticos y<br />
<strong>de</strong>más no directamente vinculados con lo<br />
científico en sentido estricto estarán<br />
cuidadosamente pulidos.<br />
Ecología General<br />
Esto que podría parecer una serie <strong>de</strong><br />
requisitos superfluos y banales, es<br />
suficientemente importante como para que la<br />
mayoría <strong>de</strong> las revistas científicas <strong>de</strong>l mundo<br />
rechacen sin siquiera leer aquéllos<br />
manuscritos que no se a<strong>de</strong>cuan en forma a las<br />
normas establecidas por el editor.<br />
La habilidad para armar un buen informe (y<br />
más a<strong>de</strong>lante un buen trabajo científico) es<br />
menos común <strong>de</strong> lo que muchos creen. En<br />
realidad, cualquiera apren<strong>de</strong> rápidamente<br />
cómo se utiliza un espectrofotómetro, la<br />
clasificación <strong>de</strong> un grupo <strong>de</strong> animales o<br />
plantas, el método <strong>de</strong> Winkler, o los algoritmos<br />
<strong>de</strong>l análisis <strong>de</strong> componentes principales. Todo<br />
eso es sencillo y, en rigor, son tareas técnicas<br />
que una vez aprendidas no exce<strong>de</strong>n en<br />
complejidad a una receta <strong>de</strong> cocina. Pero el<br />
paso que separa una planilla <strong>de</strong> datos <strong>de</strong> un<br />
trabajo listo para presentar es un escollo<br />
fundamental para la mayoría <strong>de</strong> los<br />
investigadores bisoños, y frecuentemente<br />
también para los no tan bisoños. Esto se ve<br />
reflejado en el mal que aqueja a una gran<br />
parte <strong>de</strong> los investigadores <strong>de</strong> nuestro medio:<br />
la profusión en sus curricula <strong>de</strong> informes<br />
internos y <strong>de</strong> trabajos en revistas <strong>de</strong> baja<br />
categoría. Esto especialmente penoso cuando<br />
la información sobre la que están basados<br />
esos informes y trabajos es buena, y<br />
a<strong>de</strong>cuadamente armada podría merecer una<br />
buena publicación con difusión internacional.<br />
Esto no es lo más común, pero suce<strong>de</strong> con<br />
cierta frecuencia. No en vano hace unos años<br />
atrás, con apoyo <strong>de</strong>l CONICET, se organizó<br />
un curso especial para, precisamente, enseñar<br />
a los investigadores jóvenes a armar<br />
manuscritos científicos.<br />
Para el investigador principiante existen varios<br />
manuales con consejos útiles <strong>de</strong> cómo se<br />
<strong>de</strong>be encarar el armado <strong>de</strong> un trabajo<br />
científico (e.g., M. O'Connor y F.P. Woodford,<br />
1975, Writing scientific papers in English. An<br />
Else-Ciba gui<strong>de</strong> for authors. Pitman, London;<br />
W. Cochran, P. Fenner y M. Hill, 1974,<br />
Geowriting. A gui<strong>de</strong> to writing, editing and<br />
printing in earth science. Amer. Geol. Inst.,<br />
Falls Church). Sin embargo, uno <strong>de</strong> los<br />
caminos más seguros y efectivos es leyendo<br />
las publicaciones <strong>de</strong> otros autores (claro,<br />
tratando <strong>de</strong> elegir las buenas...). Deténgase a<br />
17
estudiar no solamente el contenido y los datos,<br />
sino la manera <strong>de</strong> presentar la información, la<br />
hilación entre argumentos, el tipo <strong>de</strong> datos<br />
incluidos en las diferentes secciones<br />
(introducción, resultados, discusión), la<br />
puntuación, las figuras. Cuantos más trabajos<br />
buenos lea - más se irá acostumbrando a<br />
hilvanar sus propios resultados <strong>de</strong> manera<br />
lógica y fácil <strong>de</strong> compren<strong>de</strong>r para el lector.<br />
A continuación se <strong>de</strong>tallan algunos consejos,<br />
tanto referentes a aspectos estilísticos y<br />
gramaticales, como a los estrictamente<br />
formales. Léalos con atención antes <strong>de</strong> armar<br />
su primer informe. Una vez que lo haya<br />
completado, léalo nuevamente con ojos<br />
críticos y vea si pue<strong>de</strong> mejorarlo en función <strong>de</strong><br />
estas recomendaciones. Tenga en cuenta que<br />
<strong>de</strong>trás <strong>de</strong> un buen manuscrito enviado a una<br />
revista, y <strong>de</strong>trás <strong>de</strong> un buen informe, hay al<br />
menos 5 o 10 borradores que se fueron<br />
<strong>de</strong>purando y corrigiendo hasta llegar a la<br />
versión <strong>de</strong>finitiva. No espere a que los errores<br />
los encuentre el docente o el árbitro;<br />
seguramente muchos <strong>de</strong> los que quedan<br />
<strong>de</strong>spués <strong>de</strong>l primer par <strong>de</strong> intentos son<br />
suficientemente gruesos como para que usted<br />
mismo los <strong>de</strong>tecte antes <strong>de</strong> presentar el<br />
trabajo.<br />
Este tipo <strong>de</strong> normas se pue<strong>de</strong>n consultar en<br />
las "Instrucciones para los autores" <strong>de</strong><br />
cualquier publicación científica periódica;<br />
consiga algunas y revíselas. Al final se da un<br />
ejemplo <strong>de</strong> cuestionario que recibe el árbitro<br />
<strong>de</strong>l manuscrito para evaluarlo; analice su<br />
propio informe con este cuestionario en mano.<br />
GENERALIDADES<br />
Se acostumbra dividir los trabajos <strong>de</strong><br />
investigación en las siguientes secciones:<br />
Título, autores y filiación<br />
Resumen. El resumen <strong>de</strong>be reflejar el<br />
contenido <strong>de</strong>l trabajo <strong>de</strong> una manera inteligible<br />
para el lector que no ha leído el texto <strong>de</strong>l<br />
artículo. Especifique concisamente qué se<br />
realizó, qué se encontró y qué se concluyó.<br />
Evite expresiones vagas <strong>de</strong>l tipo "Se discuten<br />
las relaciones entre la temperatura y la<br />
abundancia...", pero mencione qué tipo <strong>de</strong><br />
relación se encontró. Debe ser breve, sin<br />
referencias bibliográficas ni referencias al<br />
cuerpo principal <strong>de</strong>l trabajo mismo.<br />
Introducción: Generalmente cubre<br />
antece<strong>de</strong>ntes sobre el tema <strong>de</strong>l estudio,<br />
información pertinente previa. Ubica al lector<br />
en el objeto <strong>de</strong>l trabajo.<br />
Ecología General<br />
Sobre el final <strong>de</strong> la sección se explicitan las<br />
hipótesis y predicciones y se presentan los<br />
objetivos <strong>de</strong>l trabajo.<br />
Materiales y métodos. Es una <strong>de</strong>scripción<br />
<strong>de</strong>tallada <strong>de</strong>l área y las fechas <strong>de</strong>l muestreo,<br />
las herramientas empleadas, las metodologías<br />
<strong>de</strong> campo y <strong>de</strong> laboratorio seguidas, etc. Estos<br />
datos <strong>de</strong>berían ser suficientes como para que<br />
cualquier interesado pueda repetir las<br />
experiencias <strong>de</strong>scriptas (si es que son<br />
repetibles, obviamente). También <strong>de</strong>bería<br />
permitir al lector evaluar el nivel <strong>de</strong> precisión y<br />
confiabilidad <strong>de</strong> los resultados presentados.<br />
Por ejemplo, si se trata <strong>de</strong> un trabajo con<br />
recuentos <strong>de</strong> fitopláncteres, indicar las<br />
cantida<strong>de</strong>s <strong>de</strong> individuos contados por<br />
muestra, los tamaños <strong>de</strong> las submuestras, etc.<br />
No <strong>de</strong>be omitir nada importante, pero tampoco<br />
<strong>de</strong>tenerse en irrelevancias como, por ejemplo,<br />
el mo<strong>de</strong>lo y la marca <strong>de</strong>l microscopio utilizado,<br />
o la marca <strong>de</strong>l motor fuera <strong>de</strong> borda con que<br />
estaba equipada la embarcación.<br />
También se presentan en esta sección las<br />
técnicas estadísticas que se utilizan para el<br />
análisis <strong>de</strong> los resultados y el software<br />
empleado.<br />
Resultados. Se restringe a los resultados <strong>de</strong>l<br />
trabajo realizado estrictamente. Si éstos están<br />
presentados en tablas y/o figuras, no repita la<br />
misma información en el texto; solamente<br />
<strong>de</strong>staque los aspectos que consi<strong>de</strong>ra salientes<br />
y dignos <strong>de</strong> especial atención. No incluya<br />
discusiones ni comparaciones con resultados<br />
ajenos bajo este acápite.<br />
Discusión (o Discusión y Conclusiones). Es el<br />
análisis pormenorizado <strong>de</strong>l significado <strong>de</strong> los<br />
resultados. Incluye las implicancias <strong>de</strong> los<br />
datos obtenidos para el proceso estudiado y<br />
otros relacionados con él, comparaciones con<br />
otras experiencias propias o ajenas, etc. A<br />
veces es conveniente que las secciones <strong>de</strong><br />
resultados y discusión sean tratadas<br />
conjuntamente, aunque esta modalidad suele<br />
conllevar más problemas <strong>de</strong> interpretación por<br />
parte <strong>de</strong>l lector.<br />
Agra<strong>de</strong>cimientos. En general aquí se incluye<br />
a la o las instituciones que financiaron el<br />
trabajo y a personas particulares que<br />
colaboraron con su realización.<br />
Bibliografía. Por regla general, en los trabajos<br />
<strong>de</strong> investigación la sección bibliográfica<br />
solamente incluye las referencias citadas en el<br />
texto, y toda cita en el texto <strong>de</strong>be estar<br />
<strong>de</strong>tallada en la lista bibliográfica. (Vea más<br />
abajo indicaciones acerca <strong>de</strong> este punto).<br />
18
Al enviar un manuscrito a una revista, en<br />
general se hace con la siguiente estructura:<br />
i) Epígrafes <strong>de</strong> las tablas y figuras. Todas las<br />
revistas exigen que estas referencias se<br />
agreguen al trabajo en hojas separadas e<br />
in<strong>de</strong>pendientes <strong>de</strong>l texto.<br />
ii) Tablas y figuras. Se agregan al final <strong>de</strong>l<br />
trabajo, una por página, con indicación <strong>de</strong>l<br />
título <strong>de</strong>l trabajo y número <strong>de</strong> tabla o figura.<br />
iii)Todo el texto <strong>de</strong>be estar preferentemente<br />
dactilografiado a doble espacio, en una sola<br />
cara <strong>de</strong>l papel, y todas las páginas <strong>de</strong>ben<br />
estar correlativamente numeradas.<br />
La revista, al aceptar el trabajo, lo publica <strong>de</strong><br />
acuerdo a su propio diseño.<br />
EL TEXTO<br />
Trate <strong>de</strong> escribir con idioma claro, sencillo y<br />
preciso. Sobre todo sencillo. Las frases<br />
floridas no agregan valor al contenido, pero<br />
dificultan la lectura. Por ejemplo, "A nivel <strong>de</strong> la<br />
estructura poblacional <strong>de</strong> los artrópodos<br />
acuáticos analizados se observó una clara<br />
ten<strong>de</strong>ncia a la dominancia <strong>de</strong> los estadios<br />
avanzados <strong>de</strong> <strong>de</strong>sarrollo ontogenético" es lo<br />
mismo que "La mayoría <strong>de</strong> los copépodos<br />
eran adultos".<br />
En la medida <strong>de</strong> lo posible, en todas las<br />
secciones se <strong>de</strong>be seguir algún tipo <strong>de</strong><br />
secuencia lógica: <strong>de</strong> lo más general a lo más<br />
particular, o en or<strong>de</strong>n <strong>de</strong> complejidad<br />
creciente. Por ejemplo, en la introducción <strong>de</strong>l<br />
trabajo sobre la laguna El Burro, primero<br />
<strong>de</strong>fina brevemente las características <strong>de</strong> las<br />
lagunas pampásicas en general, y luego hable<br />
<strong>de</strong> las <strong>de</strong> El Burro en particular. Cuidado: aquí<br />
todavía no irán los resultados <strong>de</strong> su propio<br />
trabajo, sino aquello <strong>de</strong> la bibliografía que se<br />
consi<strong>de</strong>re <strong>de</strong> relevancia (por ejemplo, sus<br />
rasgos morfométricos, su conexión con<br />
lagunas vecinas, etc.). Cuando <strong>de</strong>scriba los<br />
análisis realizados, agrupe los abióticos por un<br />
lado y los biológicos por otro, Dentro <strong>de</strong> los<br />
primeros comience por los más sencillos<br />
(temperatura, transparencia), y siga con los<br />
más complejos (nutrientes, sedimentos). En<br />
los resultados biológicos es razonable<br />
<strong>de</strong>scribir primero lo referente a las plantas<br />
(producción primaria), y luego los animales.<br />
Para los listados <strong>de</strong> organismos observados,<br />
i<strong>de</strong>ntificados o cuantificados adopte un<br />
or<strong>de</strong>namiento natural y respételo en todos<br />
aquéllos lugares (texto, tablas, figuras) don<strong>de</strong><br />
se refiera al tema.<br />
A veces es conveniente dividir el tratamiento<br />
<strong>de</strong> un tópico en subtítulos breves. Por ejemplo,<br />
Ecología General<br />
en la <strong>de</strong>scripción <strong>de</strong> las mediciones realizadas<br />
en El Burro:<br />
Temperatura: con termómetro <strong>de</strong> mercurio;<br />
Transparencia: con disco <strong>de</strong> Secchi...<br />
Pero cuando trate los antece<strong>de</strong>ntes <strong>de</strong>l<br />
estudio <strong>de</strong> lagunas pampásicas el mismo<br />
estilo es totalmente ina<strong>de</strong>cuado. En vez <strong>de</strong>:<br />
Dangaus (1976): morfometría;<br />
Tell (1973): perifiton;<br />
Ringuelet et al. (1967): zooplancton...<br />
<strong>de</strong>berá armar la sección <strong>de</strong> manera más<br />
coloquial:<br />
Des<strong>de</strong> los años '60 se realizaron numerosos<br />
estudios en las lagunas pampásicas,<br />
incluyendo aspectos <strong>de</strong> su morfología<br />
(Dangaus, 1976), química (Ringuelet et al.,<br />
1967)...<br />
Trate <strong>de</strong> que las frases sean breves, pero sin<br />
caer en un estilo telegráfico.<br />
Existen reglas para el uso <strong>de</strong> los signos <strong>de</strong><br />
puntuación: apréndalas y respételas. No<br />
abuse <strong>de</strong> los puntos y aparte.<br />
Los espacios tienen tanta importancia como<br />
los signos y letras; por regla general, los<br />
signos <strong>de</strong> puntuación <strong>de</strong>ben ir seguidos <strong>de</strong> un<br />
espacio (coma, punto, punto y coma). Hay<br />
algunas excepciones, como por ejemplo en las<br />
listas bibliográficas (ver más abajo). No va<br />
espacio entre los paréntesis y su contenido.<br />
BIBLIOGRAFIA<br />
No existen normas generales, aceptadas por<br />
todas las revistas, <strong>de</strong> cómo armar las listas<br />
bibliográficas. Hay, sin embargo, una serie <strong>de</strong><br />
coinci<strong>de</strong>ncias y pautas generales que siguen<br />
la mayoría <strong>de</strong> las publicaciones periódicas<br />
científicas.<br />
En el texto<br />
Las citas son por autor-año, por ejemplo,<br />
"Martínez (1974)" (el año entre paréntesis); o<br />
"En 1974, Martínez...". No se incluyen iniciales<br />
<strong>de</strong> los nombres, a menos que haya dos o más<br />
Martínez diferentes citados en el trabajo, en<br />
cuyo caso será "A. Martínez (1974)" o "B.<br />
Martínez (1978)". Dos autores van in extenso:<br />
"Martínez y Valle (1974)", pero para más <strong>de</strong><br />
dos se cita al primero seguido <strong>de</strong> "et al." ("al."<br />
significa aliae en latín; es una abreviatura, y<br />
por en<strong>de</strong> seguida <strong>de</strong> punto): "Martínez et al.<br />
(1974)". Si hubiera varias citas sucesivas en el<br />
texto estas se or<strong>de</strong>nan cronológicamente, y<br />
alfabéticamente <strong>de</strong>ntro <strong>de</strong>l mismo año.<br />
<strong>Trabajos</strong> <strong>de</strong>l mismo autor, mismo año se<br />
i<strong>de</strong>ntifican con letras ("Martínez, 1974a").<br />
19
En el capítulo "Bibliografía"<br />
El or<strong>de</strong>n es alfabético por apellidos <strong>de</strong>l primer<br />
autor, por apellido <strong>de</strong>l segundo autor en caso<br />
<strong>de</strong> igual primer autor, etc.; y cronológico para<br />
apellidos y nombres idénticos. La forma <strong>de</strong><br />
citar los trabajos y la utilización <strong>de</strong> bastardillas,<br />
negritas, Versalitas, etc. varía mucho <strong>de</strong> una<br />
revista a otra. Sin embargo, prácticamente<br />
siempre se incluye la siguiente información:<br />
_ Autor(es),<br />
_ Iniciales,<br />
_ Año <strong>de</strong> publicación,<br />
_ Título <strong>de</strong>l trabajo,<br />
_ Lugar don<strong>de</strong> se publicó (nombre <strong>de</strong> la<br />
revista, o nombre <strong>de</strong>l libro -si es un capítulo en<br />
una obra colegiada- con su editor y ciudad <strong>de</strong><br />
la editorial),<br />
_ Volumen o tomo (para revistas periódicas; el<br />
número suele omitirse porque la paginación es<br />
correlativa <strong>de</strong>s<strong>de</strong> el primero al último números<br />
<strong>de</strong>l mismo año),<br />
_ Páginas inicial y final, o número <strong>de</strong> páginas<br />
totales para los libros.<br />
Algunos ejemplos. Un trabajo en una revista<br />
periódica:<br />
Haury, L.R., Kenyon, D.E. y Brooks, J.R. 1980.<br />
Experimental evaluation of the avoidance<br />
reaction in Calanus finmarchcus. J. Plankton<br />
Res., 2:187-202.<br />
Nótese que no se han <strong>de</strong>jado espacios entre<br />
las iniciales <strong>de</strong> los autores, ni entre el volumen<br />
<strong>de</strong> la revista y las páginas. El título <strong>de</strong>l trabajo<br />
jamás se abrevia, y <strong>de</strong>be transcribirse<br />
exactamente como fue publicado. El nombre<br />
<strong>de</strong> la revista se suele abreviar cuando consta<br />
<strong>de</strong> varias palabras ("J. Plankton Res." es<br />
"Journal of Plankton Research"), y existen<br />
normas más o menos generales para estas<br />
abreviaturas (ver "World List of Scientific<br />
Periodicals", "ISO4 International Co<strong>de</strong> for<br />
Abbreviation of Titles of Periodicals", "ISO833<br />
International List of Periodical Title Word<br />
Abbreviations"). No se abrevian los nombres<br />
<strong>de</strong> las revistas cuando constan <strong>de</strong> una sola<br />
palabra (e.g., Physis, Micropaleontology,<br />
Sarsia, Hydrobiologia).<br />
Un capítulo <strong>de</strong> un libro:<br />
Steedman, H.F. 1976. General and applied<br />
data on formal<strong>de</strong>hy<strong>de</strong> fixation and preservation<br />
of marine zooplankton. En: Zooplankton<br />
fixation and preservation (H.F. Steedamn, ed.),<br />
UNESCO Press, Paris, pp. 103-154.<br />
Un libro:<br />
Ecología General<br />
Lewis, W.M., Jr. 1979. Zooplankton community<br />
analysis. Springer, New York, 163 pp.<br />
Nuevamente, no hay normas universales para<br />
la presentación <strong>de</strong> listas bibliográficas. Es<br />
importante, sin embargo, que las citas sean<br />
consistentes a lo largo <strong>de</strong> toda la lista.<br />
TABLAS Y FIGURAS<br />
Un manuscrito está integrado por el cuerpo<br />
principal <strong>de</strong> texto, las tablas y las figuras.<br />
Nada más. No existen los "cuadros",<br />
"láminas", "diagramas", etc. Las tablas son<br />
listados <strong>de</strong> valores numéricos o alfanuméricos,<br />
y las figuras son ilustraciones lineales y/o<br />
fotografías. Ambos se numeran<br />
correlativamente <strong>de</strong> acuerdo a su or<strong>de</strong>n <strong>de</strong><br />
aparición en el texto, con numeraciones<br />
in<strong>de</strong>pendientes.<br />
Prácticamente ninguna revista <strong>de</strong>l mundo<br />
acepta un manuscrito, ni siquiera para una<br />
evaluación preliminar, si no va acompañado<br />
<strong>de</strong> figuras preparadas profesionalmente,<br />
correctamente rotuladas y entintadas. Mire sus<br />
gráficos: ¿están los ejes <strong>de</strong>bidamente<br />
rotulados? ¿están las unida<strong>de</strong>s indicadas?<br />
Normalmente, la figura y su epígrafe <strong>de</strong>ben<br />
ser suficientes para enten<strong>de</strong>r lo que se ilustra,<br />
sin necesidad <strong>de</strong> recurrir al texto. ¿Tiene<br />
epígrafe su figura? ¿Es conciso, claro e<br />
informativo el epígrafe?<br />
Las figuras <strong>de</strong>ben ser sencillas, claras e<br />
informativas. No agregue adornos, etiquetas<br />
superfluas, datos innecesarios; todo eso<br />
distrae la atención y no contribuye a que el<br />
lector entienda lo que se preten<strong>de</strong> mostrar.<br />
Por ejemplo, la tridimensionalidad en los<br />
diagramas corrientes <strong>de</strong> barras no agrega<br />
nada a la información, -más aún, la<br />
enmascara. Evítelos.<br />
En los valores numéricos en el texto, las tablas<br />
y las figuras limite la cantidad <strong>de</strong> <strong>de</strong>cimales a<br />
lo significativo (y no a lo que da la<br />
computadora). En los índices <strong>de</strong> correlación,<br />
por ejemplo, no especifique más <strong>de</strong> 3<br />
<strong>de</strong>cimales.<br />
VARIOS<br />
Bastardillas o itálicas<br />
Todos los nombres latinos <strong>de</strong>s<strong>de</strong> género hasta<br />
subespecie van en este tipo <strong>de</strong> letra o<br />
subrayados con línea simple (que, para la<br />
imprenta, significa bastardillas). Las palabras<br />
en idiomas diferentes al <strong>de</strong>l texto, inclusive el<br />
latín, también suelen ir en bastardillas. Las<br />
20
abreviaturas <strong>de</strong> las locuciones latinas más<br />
comúnmente utilizadas en trabajos científicos,<br />
sin embargo, frecuentemente no se escriben<br />
en bastardillas sino en letra común (redonda),<br />
por ejemplo "etc." (por et cetera); "et al." (por<br />
et aliae); "in litt." (por in litteris).<br />
Signos especiales<br />
Los caracteres especiales (µ, Σ, Δ y otros) se<br />
agregarán a mano si la impresora o máquina<br />
<strong>de</strong> escribir no los poseyera. No los reemplace<br />
por la palabra correspondiente ("microm",<br />
"sumatoria", "<strong>de</strong>lta").<br />
Unida<strong>de</strong>s<br />
Existe una convención internacional para las<br />
abreviaturas <strong>de</strong> las unida<strong>de</strong>s <strong>de</strong> medida<br />
(distancia, peso, volumen): ninguna <strong>de</strong> estas<br />
abreviaturas va seguida <strong>de</strong> punto (por<br />
ejemplo, "m", pero no "m." ni "M"; "µm", y no<br />
"um" o "uM."). Kilómetros es "km" (no "Km").<br />
Consulte en caso <strong>de</strong> duda.<br />
MODELO DE PLANILLA QUE ENVIAN LOS<br />
EDITORES A LOS ARBITROS PARA LA<br />
EVALUACION DE LOS MANUSCRITOS<br />
RECIBIDOS<br />
Con algunas variantes, este es el mo<strong>de</strong>lo<br />
básico <strong>de</strong> formulario que utilizan la mayoría <strong>de</strong><br />
las revistas científicas al solicitar evaluación<br />
(2) EJERCICIO<br />
Ecología General<br />
<strong>de</strong> los manuscritos que reciben. Sin bien los<br />
ítems referentes a la originalidad y cantidad <strong>de</strong><br />
información no son relevantes en el caso <strong>de</strong><br />
los informes <strong>de</strong>l curso, la mayoría <strong>de</strong> los otros<br />
puntos sí lo son. Critique su propio informe<br />
sobre la base <strong>de</strong> estas preguntas y trate <strong>de</strong><br />
mejorarlo.<br />
¿Constituye el trabajo un aporte serio y<br />
original al conocimiento <strong>de</strong>l tema?<br />
¿Demuestran los autores buen conocimiento<br />
<strong>de</strong>l tema y <strong>de</strong> la bibliografía pertinente?<br />
¿La calidad y cantidad <strong>de</strong> información<br />
presentada justifica su publicación?<br />
¿Existe repetición superflua <strong>de</strong> información<br />
(en el texto, tablas y figuras)?<br />
¿Es a<strong>de</strong>cuada la organización general <strong>de</strong>l<br />
trabajo? ¿Pue<strong>de</strong> ser mejorada?<br />
¿Es el título breve y conciso? ¿Refleja<br />
a<strong>de</strong>cuadamente el contenido <strong>de</strong>l trabajo?<br />
¿Es el resumen conciso e informativo?<br />
¿Son todas las ilustraciones a<strong>de</strong>cuadas y<br />
necesarias?<br />
¿Y las tablas?<br />
¿Pue<strong>de</strong> ser mejorado el manuscrito? ¿Cómo?<br />
En su opinión, este trabajo:<br />
Pue<strong>de</strong> ser publicado sin modificaciones;<br />
Pue<strong>de</strong> ser publicado con pequeños cambios;<br />
Requiere cambios sustanciales y una nueva<br />
evaluación;<br />
Debe ser rechazado.<br />
por Haydée Pizarro<br />
A continuación figura la cita bibliográfica y el resumen <strong>de</strong> un trabajo científico publicado en una revista<br />
nacional:<br />
VIGNOLIO OR, MACEIRA NO y FERNANDEZ ON (1995) Efectos <strong>de</strong>l anegamiento sobre el po<strong>de</strong>r<br />
germinativo <strong>de</strong> las semillas <strong>de</strong> Lotus tenuis y Lotus corniculatus. Implicancias para su propagación<br />
diferencial en la Depresión <strong>de</strong>l Salado (Buenos Aires, Argentina). Ecología Austral, 5:157-163.<br />
Resumen. Lotus tenuis (L.t.) y Lotus corniculatus (L.c.) fueron introducidas como forrajeras en la<br />
Pampa Deprimida hace unos cincuenta años. Mientras que L.t. se ha difundido naturalmente en<br />
los campos bajos anegables <strong>de</strong> la región, L.c. no ha experimentado la misma expansión. En el<br />
presente trabajo se estudió la tolerancia al anegamiento <strong>de</strong> semillas <strong>de</strong> ambas especies en<br />
procura <strong>de</strong> explicar dicha distribución. Los experimentos <strong>de</strong> anegamiento se realizaron en<br />
recipientes que permanecieron en condiciones controladas <strong>de</strong> temperatura y luz. Las semillas<br />
estuvieron anegadas durante siete semanas y se realizaron muestreos a intervalos crecientes <strong>de</strong><br />
tiempo. El po<strong>de</strong>r germinativo (P.G., medida <strong>de</strong> la tolerancia al anegamiento), disminuyó en ambas<br />
especies al aumentar el tiempo <strong>de</strong> anegamiento, pero lo hizo en forma más acentuada en L.c. que<br />
en L.t.. La fracción <strong>de</strong> semillas blandas perdió antes el P.G. en L.c. que en L.t.. Con 49 días <strong>de</strong><br />
anegamiento las semillas que conservaron el P.G. fueron las duras, las cuales se hallaban en<br />
mayor proporción en L.t (aproximadamente 50% en L.t y 6% en L.c., para semillas <strong>de</strong> 5 meses <strong>de</strong><br />
edad). La capacidad <strong>de</strong> las semillas <strong>de</strong> L.t. <strong>de</strong> tolerar períodos prolongados <strong>de</strong> anegamiento<br />
jugaría un papel <strong>de</strong>terminante en la expansión <strong>de</strong> la especie en los campos sujetos a<br />
inundaciones periódicas.<br />
21
Ecología General<br />
Los siguientes párrafos y oraciones fueron tomados textualmente <strong>de</strong>l trabajo científico cuyos<br />
autores, título y resumen figuran arriba. ¿A qué sección <strong>de</strong>l trabajo pertenece cada párrafo u oración?<br />
Indicar en el paréntesis <strong>de</strong> la izquierda: I = Introducción; MM = Materiales y Métodos; R = Resultados;<br />
D = Discusión; E = Epígrafe y A = Agra<strong>de</strong>cimientos.<br />
1. (......) En experimentos anteriores se <strong>de</strong>mostró que las plantas <strong>de</strong> L. tenuis son más tolerantes<br />
al anegamiento que las <strong>de</strong> L. corniculatus (Vignolo et al., 1994 a,b), aspecto que, en parte,<br />
contribuye a explicar la ausencia <strong>de</strong> la segunda en los suelos bajos sujetos a anegamientos<br />
(Grime et al., 1988, Mazzanti et al., 1988).<br />
2. (......) Efectos <strong>de</strong>l período <strong>de</strong> anegamiento sobre el po<strong>de</strong>r germinativo <strong>de</strong> semillas blandas <strong>de</strong><br />
Lotus tenuis (L.t.) y Lotus corniculatus (L.c.) (experimento 1). Referencias: (T), testigo y (A),<br />
anegado. Letras distintas para una misma fecha <strong>de</strong> anegamiento indican diferencias significativas<br />
(P
16. (......) Se verificó la homogeneidad <strong>de</strong> varianza mediante prueba <strong>de</strong> Barlett.<br />
Ecología General<br />
17. (......) A Arturo Valver<strong>de</strong> Lyons por la información <strong>de</strong> su trabajo inédito “Lotus tenuis en la<br />
Depresión <strong>de</strong>l Salado, Prov. <strong>de</strong> Bs. As., Argentina”.<br />
18. (......) Estos resultados guardan relación con la situación que presentan ambas especies <strong>de</strong><br />
Lotus, en cuanto a su tolerancia al anegamiento y distribución diferencial en la Depresión <strong>de</strong>l<br />
Salado (Mazzanti et al., 1988; Vignolio et al., 1994a,b).<br />
19. (......) Efectos <strong>de</strong>l período <strong>de</strong> anegamiento sobre el po<strong>de</strong>r germinativo <strong>de</strong> semillas <strong>de</strong> Lotus<br />
tenuis (L.t.) y L. corniculatus (L.c.) con distinto grado <strong>de</strong> dureza, para los experimentos 2 (a) y 3<br />
(b).<br />
20. (......) Los tratamientos se realizaron en macetas <strong>de</strong> plástico <strong>de</strong> 65 ml con 90 semillas cada<br />
una, con drenaje para los testigos (T) y sin drenaje para los anegados (A).<br />
23
Trabajo práctico 1<br />
PRINCIPIOS DE DISEÑO EXPERIMENTAL EN ECOLOGÍA<br />
Ecología General<br />
por David Bilenca<br />
En los últimos años se ha verificado un creciente aumento en los porcentajes <strong>de</strong> trabajos<br />
experimentales <strong>de</strong> campo en ecología, pasando <strong>de</strong> menos <strong>de</strong>l 5% durante las décadas <strong>de</strong>l ‘50 y ‘60 a<br />
cerca <strong>de</strong>l 35% en la década <strong>de</strong>l ‘80 (Hairston 1989). Probablemente ello esté asociado a que los<br />
experimentos permiten establecer <strong>de</strong> manera unívoca mecanismos <strong>de</strong> causa y efecto, algo que,<br />
probablemente, ninguna otra forma <strong>de</strong> aproximación a la investigación sea capaz <strong>de</strong> lograr.<br />
Sin embargo, <strong>de</strong>be tenerse en cuenta que los experimentos no dicen cuáles son las preguntas<br />
que uno como científico <strong>de</strong>be formularse. Por otra parte, aún cuando la pregunta planteada fuera<br />
apropiada, pue<strong>de</strong> suce<strong>de</strong>r que durante la ejecución <strong>de</strong>l experimento no se la haya puesto a prueba <strong>de</strong><br />
la manera a<strong>de</strong>cuada por fallas en su diseño. El diseño experimental es la estructura lógica <strong>de</strong> un<br />
experimento (Galindo Leal 1997). Durante el diseño <strong>de</strong> un experimento <strong>de</strong>ben especificarse, entre<br />
otros, los siguientes aspectos:<br />
• I<strong>de</strong>ntificación <strong>de</strong>l tipo <strong>de</strong> unida<strong>de</strong>s experimentales<br />
• número y tipo <strong>de</strong> tratamientos que se han <strong>de</strong> aplicar (incluyendo los controles)<br />
• las variables o propieda<strong>de</strong>s que se han <strong>de</strong> medir sobre las unida<strong>de</strong>s experimentales<br />
• la asignación <strong>de</strong> los tratamientos a las unida<strong>de</strong>s experimentales<br />
• el número <strong>de</strong> unida<strong>de</strong>s experimentales (réplicas) que han <strong>de</strong> recibir cada uno <strong>de</strong> los tratamientos<br />
• el arreglo espacial <strong>de</strong> las unida<strong>de</strong>s experimentales<br />
• la secuencia temporal <strong>de</strong> aplicación y medición <strong>de</strong> las unida<strong>de</strong>s experimentales<br />
Probablemente, uno <strong>de</strong> los errores <strong>de</strong> diseño más frecuentes en ecología sea el <strong>de</strong> la llamada<br />
pseudorreplicación (Hurlbert 1984), que consiste en la utilización <strong>de</strong> réplicas que no son<br />
in<strong>de</strong>pendientes entre sí. La pseudorreplicación se pue<strong>de</strong> originar <strong>de</strong> un mal diseño o <strong>de</strong> un mal<br />
análisis (Galindo Leal 1997). El diseño y la ejecución <strong>de</strong> un experimento son los pasos más críticos<br />
<strong>de</strong> la experimentación. Si el análisis estadístico fue inapropiado o la interpretación fue incorrecta, los<br />
datos pue<strong>de</strong>n ser nuevamente analizados; en cambio, si los errores se cometieron durante el diseño<br />
o la ejecución, éstos suelen ser insalvables y <strong>de</strong>be necesariamente volver a repetirse el experimento.<br />
A continuación se presenta una síntesis <strong>de</strong> los conceptos relativos al diseño experimental <strong>de</strong> dos<br />
<strong>de</strong> los ecólogos con más fuerte vocación por la experimentación <strong>de</strong> campo en ecología: Charles<br />
Krebs y Nelson Hairston.<br />
(1) La visión <strong>de</strong> Krebs: principios generales <strong>de</strong>l diseño experimental (Krebs 1989)<br />
Para Krebs, “un experimento es un intento por poner a prueba una hipótesis acerca <strong>de</strong> la<br />
naturaleza”.<br />
“Las observaciones naturales (...) también pue<strong>de</strong>n ser usadas para probar hipótesis, y esto<br />
constituye <strong>de</strong> hecho otro uso <strong>de</strong>l método experimental. La característica esencial <strong>de</strong> un experimento<br />
en ciencia es un conjunto <strong>de</strong> observaciones concebidas para poner a prueba una hipótesis (Medawar<br />
1957, en Krebs 1988).<br />
De este modo, los experimentos pue<strong>de</strong>n clasificarse en dos tipos:<br />
1. Experimentos <strong>de</strong> medición (mensurative experiments, Hurlbert 1984): no se aplican tratamientos<br />
a las unida<strong>de</strong>s experimentales. Los experimentos incluyen mediciones en el espacio o en el<br />
tiempo, <strong>de</strong> modo tal que el único tratamiento o variable “experimental” es el espacio o el tiempo.<br />
Ejemplo: en el estudio <strong>de</strong> la distribución <strong>de</strong> una especie, diferentes hábitats actúan como<br />
diferentes “tratamientos”.<br />
24
Ecología General<br />
2. Experimentos manipulativos (manipulative experiments, Hurlbert 1984): sí se aplican tratamientos<br />
a las unida<strong>de</strong>s experimentales. Una o más variables <strong>de</strong> interés son manejadas <strong>de</strong> manera<br />
pre<strong>de</strong>terminada por el experimentador. En los experimentos realizados a campo se supone que<br />
las variables no controladas por el experimentador afectan <strong>de</strong> manera equivalente a todos los<br />
tratamientos, o que, al menos, lo hacen aleatoriamente.<br />
Para Krebs, la mayoría <strong>de</strong> los ecólogos sólo utilizan la palabra “experimento” para referirse a los<br />
experimentos manipulativos, pero él consi<strong>de</strong>ra que esto es muy restrictivo.<br />
Algunos consejos <strong>de</strong> Krebs:<br />
1. Cada experimento manipulativo <strong>de</strong>be tener un control. Si el control no está presente es<br />
imposible concluir nada <strong>de</strong>finitivo acerca <strong>de</strong>l experimento (NOTA 1: A veces se aplican dos o más<br />
tratamientos para saber cuál es el mejor. En este caso, un tratamiento pue<strong>de</strong> actuar como<br />
“control” <strong>de</strong>l otro. NOTA 2: Generalmente se entien<strong>de</strong> por control a aquella unidad experimental<br />
que no recibe tratamiento alguno, aunque en <strong>de</strong>terminados casos los controles pue<strong>de</strong>n recibir<br />
“tratamientos fingidos". Por ejemplo, para analizar el efecto <strong>de</strong> un fertilizante que sólo pue<strong>de</strong><br />
suministrarse mediante una solución acuosa, el control consistirá en un rociado con agua y el<br />
tratamiento en un rociado con agua más fertilizante).<br />
2. Dada la gran variación temporal en los estudios realizados a campo, cada experimento<br />
manipulativo realizado a campo <strong>de</strong>be tener un control contemporáneo. Si no se dispone <strong>de</strong><br />
este control, todas las comparaciones entre antes y <strong>de</strong>spués <strong>de</strong> la aplicación <strong>de</strong>l tratamiento<br />
<strong>de</strong>berían suponer la homogeneidad a través <strong>de</strong>l tiempo<br />
3. Aleatorice siempre que sea posible. En muchas situaciones ecológicas, la aleatorización no es<br />
posible, ya que: (a) no todas las áreas <strong>de</strong> estudio están siempre disponibles para la investigación<br />
ecológica, (b) es imposible acce<strong>de</strong>r libremente a las zonas disponibles, etc.<br />
4. Hacer buena ecología implica tener i<strong>de</strong>as excitantes y efectuar un buen diseño experimental. Ser<br />
un biómetra <strong>de</strong> primera línea no te convertirá en un buen ecólogo, pero muchas buenas i<strong>de</strong>as se<br />
pier<strong>de</strong>n porque el diseño es pobre.<br />
5. Uno <strong>de</strong> los objetivos que <strong>de</strong>be perseguir un buen diseño es el <strong>de</strong> reducir el error<br />
experimental y lograr que las conclusiones <strong>de</strong>l mismo sean lo más precisas posibles. Para<br />
ello: (a) utilice unida<strong>de</strong>s experimentales homogéneas, (b) aumente la cantidad <strong>de</strong> réplicas<br />
(siempre funciona si se cuenta con el tiempo y el dinero suficientes), y (c) aumente la eficiencia<br />
<strong>de</strong> su diseño: aspectos <strong>de</strong> balanceo, bloques, anidamiento, etc.<br />
(2) La visión <strong>de</strong> Hairston: requerimientos mínimos <strong>de</strong> diseño experimental en ecología (Hairston<br />
1989)<br />
1. Conocimiento <strong>de</strong> las condiciones iniciales: Para Hairston, una <strong>de</strong>scripción previa a<strong>de</strong>cuada <strong>de</strong>l<br />
área <strong>de</strong> estudio, con recolección <strong>de</strong> datos <strong>de</strong> base (“baseline data”) ofrece las siguientes<br />
utilida<strong>de</strong>s:<br />
• Cuando se proyecta realizar un diseño en bloques, permite seleccionar las unida<strong>de</strong>s<br />
experimentales más homogéneas para asignar a los distintos bloques.<br />
• Permite comparar las diferencias existentes entre antes y <strong>de</strong>spués <strong>de</strong> la aplicación <strong>de</strong> un<br />
tratamiento.<br />
• A mayor rango <strong>de</strong> condiciones iniciales, mayor generalidad <strong>de</strong> los resultados posteriores.<br />
2. Controles: permiten establecer qué hubiera sucedido si no se hubiera realizado el experimento<br />
3. Réplicas: Para Hairston, el gran dolor <strong>de</strong> cabeza <strong>de</strong> la experimentación ecológica es la<br />
variabilidad <strong>de</strong> las condiciones, y la principal <strong>de</strong>fensa contra ello es realizar una suficiente<br />
replicación.<br />
4. Minimizar en lo posible los efectos no <strong>de</strong>seados <strong>de</strong>l experimento: Ejemplos: efecto clausura -<br />
trampafobia - trampafilia - neofobia - cambios comportamentales <strong>de</strong>bidos a la presencia <strong>de</strong>l<br />
observador - <strong>de</strong>predación extra <strong>de</strong>bido al seguimiento <strong>de</strong> huellas <strong>de</strong> los investigadores a los<br />
nidos, etc.<br />
25
REFERENCIAS<br />
Ecología General<br />
Galindo Leal (1997) Métodos cuantitativos para el manejo <strong>de</strong> la diversidad biológica. Manual. Wasai<br />
Lodge & Research center, Center for Conservation Biology, Stanford University. Colegio <strong>de</strong><br />
Biólogos <strong>de</strong>l Perú, 64 pp.<br />
Hairston NG Sr (1989) Ecological experiments. Purpose, <strong>de</strong>sign and execution. Cambridge University<br />
Press, Cambridge. Capítulos 1-3.<br />
Hurlbert SH (1984) Pseudoreplication and the <strong>de</strong>sign of ecological experiments. Ecol. Mon. 54:187-<br />
211.<br />
Krebs CJ (1988) The experimental approach to ro<strong>de</strong>nt population dynamics. Oikos 52: 143-149.<br />
Krebs CJ (1989) Ecological Methodology. Harper & Row. New York.<br />
_______________________________________________________________________________<br />
DISEÑO EXPERIMENTAL II<br />
por Ricardo Gürtler<br />
El diseño experimental involucra <strong>de</strong>terminar la forma en que los niveles <strong>de</strong> los factores o tratamientos<br />
son asignados a las unida<strong>de</strong>s experimentales, la elección <strong>de</strong>l tamaño muestral, y la disposición<br />
espacial y temporal <strong>de</strong>l experimento. Un factor pue<strong>de</strong> estar representado por un número <strong>de</strong> niveles<br />
(i.e., categorías en que se subdivi<strong>de</strong> un factor). Una unidad experimental se <strong>de</strong>fine como la más<br />
pequeña división <strong>de</strong>l material experimental a la cual se le pue<strong>de</strong> aplicar un tratamiento (o que<br />
pue<strong>de</strong> recibir un tratamiento). El diseño experimental <strong>de</strong>termina el mo<strong>de</strong>lo <strong>de</strong> ANOVA que se usará<br />
para poner a prueba la hipótesis y no viceversa.<br />
El diseño más sencillo y común es el diseño completamente aleatorizado, en el cual los<br />
niveles <strong>de</strong>l factor se asignan aleatoriamente en el espacio. El mo<strong>de</strong>lo lineal que <strong>de</strong>scribe este diseño<br />
no incluye un término que representa la variabilidad <strong>de</strong>bida al ambiente. Si existen efectos<br />
ambientales fuertes, entonces el término <strong>de</strong> error se incrementará sensiblemente y se per<strong>de</strong>rá<br />
potencia para <strong>de</strong>tectar diferencias significativas. Este diseño entonces pue<strong>de</strong> llegar a confundir dos<br />
fuentes <strong>de</strong> variación diferentes: la <strong>de</strong>l factor (la que nos interesa) y la ambiental (que interfiere con<br />
nuestro objeto <strong>de</strong> estudio).<br />
Aún en ambientes controlados y supuestamente homogéneos, tales como inverna<strong>de</strong>ros y<br />
cámaras <strong>de</strong> cultivo experimentales, frecuentemente se <strong>de</strong>tectan gradientes <strong>de</strong> heterogeneidad en<br />
temperatura, luz, humedad relativa, etc., que afectan los resultados experimentales. Para<br />
contrarrestar estos efectos se utiliza el procedimiento <strong>de</strong> “blocking” durante la etapa <strong>de</strong>l diseño. En<br />
este procedimiento, las unida<strong>de</strong>s experimentales se agrupan en bloques que poseen condiciones<br />
ambientales relativamente similares y que difieren <strong>de</strong> las <strong>de</strong> otros bloques. Luego cada nivel <strong>de</strong>l factor<br />
es aplicado al mismo bloque. Existen dos tipos <strong>de</strong> diseños que involucran bloques y son<br />
frecuentemente utilizados cuando hay heterogeneidad ambiental: (i) el diseño <strong>de</strong> bloques<br />
aleatorizados, que se usa cuando existe un gradiente en un solo eje o cuando el ambiente es un<br />
mosaico <strong>de</strong> mejores y peores condiciones, y (ii) el diseño <strong>de</strong> cuadrados latinos, cuando los gradientes<br />
ocurren en dos direcciones perpendiculares.<br />
El diseño <strong>de</strong> bloques aleatorizados más clásico incluye una sola observación o réplica por<br />
bloque; por lo tanto, el número <strong>de</strong> unida<strong>de</strong>s experimentales <strong>de</strong>ntro <strong>de</strong> cada bloque será igual al<br />
número <strong>de</strong> niveles <strong>de</strong>l factor en estudio. Es esencial que los bloques se hallen dispuestos en forma<br />
perpendicular al gradiente ambiental que se <strong>de</strong>sea controlar. El mo<strong>de</strong>lo lineal para un diseño <strong>de</strong><br />
bloques aleatorizados incluye términos para los efectos <strong>de</strong>bidos al tratamiento, a los bloques y al<br />
error residual.<br />
26
Ecología General<br />
El diseño <strong>de</strong> cuadrados latinos involucra bloques en dos ejes y permite controlar gradientes<br />
en dos direcciones. Este diseño requiere que haya un mismo número <strong>de</strong> filas y <strong>de</strong> columnas, lo cual<br />
produce un cuadrado. Debido a que cada nivel <strong>de</strong>l factor tiene que aparecer en cada fila y en cada<br />
columna, el número total <strong>de</strong> réplicas iguala al número <strong>de</strong> filas y <strong>de</strong> columnas. El mo<strong>de</strong>lo lineal ahora<br />
incluye términos para los efectos <strong>de</strong>bidos al tratamiento, a los bloques representados por las filas, a<br />
los bloques representados por las columnas, y al error residual.<br />
Diseño <strong>de</strong> bloques aleatorizados<br />
bloque 1 C E F B D A<br />
bloque 2 F C B E D A<br />
bloque 3 F D E C A B gradiente ambiental<br />
bloque 4 C B E F D A<br />
bloque 5 F C E B D A<br />
bloque 6 B F D A C E<br />
Diseño <strong>de</strong> cuadrados latinos<br />
col 1 col 2 col 3 col 4 col 5 col 6<br />
fila 1 A B C D E F<br />
fila 2 B F D C A E<br />
fila 3 C D E F B A<br />
fila 4 D A F E C B<br />
fila 5 E C A B F D<br />
fila 6 F E B A D C<br />
BIBLIOGRAFÍA<br />
gradientes ambientales<br />
Cochran WG y Cox GM (1957) Experimental <strong>de</strong>signs. 2da. ed., Wiley, New York.<br />
Winer BJ, Brown DR y Michaels KM (1991) Statistical principles in experimental <strong>de</strong>sign. McGraw-Hill,<br />
New York.<br />
Krebs CJ (1989) Ecological methodology. Harper Collins.<br />
27
Trabajo práctico 2<br />
RECURSOS Y CONDICIONES<br />
INTRODUCCIÓN<br />
Ecología General<br />
La vida <strong>de</strong>pen<strong>de</strong> totalmente <strong>de</strong>l mundo físico-químico. Los organismos necesitan <strong>de</strong> la energía solar y<br />
en muchas situaciones <strong>de</strong>ben adaptarse a condiciones extremas <strong>de</strong> temperatura, humedad, salinidad,<br />
aci<strong>de</strong>z y otros factores físicos o químicos que actúan a su alre<strong>de</strong>dor. Una condición es un factor<br />
ambiental abiótico que varía en espacio y tiempo y que influye sobre el <strong>de</strong>senvolvimiento <strong>de</strong> los<br />
individuos.<br />
Los recursos son para los organismos principalmente las materias que necesitan para formar sus<br />
cuerpos, la energía que interviene en sus activida<strong>de</strong>s, y los lugares o espacios en los que pasan sus<br />
ciclos vitales. A diferencia <strong>de</strong> las condiciones, los recursos se caracterizan por su abundancia, que<br />
pue<strong>de</strong> ser reducida a medida que los organismos los consumen o utilizan. El cuerpo <strong>de</strong> un vegetal<br />
está formado por iones y moléculas orgánicas que sintetiza a partir <strong>de</strong> sustancias inorgánicas que<br />
representan sus recursos alimenticios, mientras que la radiación solar proporciona el recurso<br />
energético al ser transformada en energía química a través <strong>de</strong>l proceso <strong>de</strong> fotosíntesis. A su vez, las<br />
plantas ver<strong>de</strong>s representan los recursos <strong>de</strong> los herbívoros, que por su parte son los recursos <strong>de</strong> los<br />
carnívoros. Tanto para los herbívoros, como para los carnívoros y los <strong>de</strong>scomponedores, tanto la<br />
energía como los elementos necesarios para formar su biomasa provienen <strong>de</strong> los alimentos. Aparte<br />
<strong>de</strong> los recursos alimentarios, los organismos requieren <strong>de</strong> otra variedad <strong>de</strong> recursos para po<strong>de</strong>r<br />
sobrevivir y <strong>de</strong>sarrollar poblaciones viables.<br />
El estudio <strong>de</strong> los recursos es particularmente importante cuando se consi<strong>de</strong>ra que aquello que un<br />
organismo consume afecta lo que queda disponible para los <strong>de</strong>más individuos <strong>de</strong> la misma o <strong>de</strong> otras<br />
especies y, por lo tanto, pue<strong>de</strong> generar competencia intra e interespecífica.<br />
Dos recursos son esenciales cuando uno es incapaz <strong>de</strong> sustituir al otro. El crecimiento <strong>de</strong><br />
una población <strong>de</strong>pen<strong>de</strong> <strong>de</strong> la disponibilidad <strong>de</strong> una cantidad <strong>de</strong>terminada <strong>de</strong>l recurso esencial,<br />
in<strong>de</strong>pendientemente <strong>de</strong> que otros recursos sean abundantes. Por ejemplo, nitrógeno y potasio como<br />
recursos para las plantas ver<strong>de</strong>s.<br />
Hay otros tipos <strong>de</strong> recursos que pue<strong>de</strong>n reemplazarse en distinto grado, los recursos<br />
sustituibles. Dos recursos son perfectamente sustituibles (Fig a) cuando cualquiera <strong>de</strong> ellos pue<strong>de</strong><br />
sustituir totalmente al otro. Para la mayoría <strong>de</strong> las plantas ver<strong>de</strong>s, esto es lo que suce<strong>de</strong> con el nitrato<br />
y los iones amonio como fuentes <strong>de</strong> nitrógeno. Pero estos dos compuestos no son iguales, ya que las<br />
plantas <strong>de</strong>ben consumir energía metabólica en la reducción <strong>de</strong>l nitrato a amonio antes <strong>de</strong> po<strong>de</strong>r<br />
asimilarlo y sintetizar sus proteínas. También distintas fuentes <strong>de</strong> carne para un carnívoro pue<strong>de</strong>n ser<br />
recursos sustituibles (liebres pue<strong>de</strong>n ser reemplazadas por roedores pequeños, por ejemplo).<br />
Los recursos sustituibles son complementarios (Fig b) si el organismo necesita una menor cantidad<br />
total <strong>de</strong> recursos (o sea, una menor suma <strong>de</strong> ambos recursos) cuando se consumen juntos que<br />
cuando son consumidos por separado. En el caso <strong>de</strong> herbívoros, el consumo <strong>de</strong> distintos tipos <strong>de</strong><br />
material vegetal, con aportes nutricionales complementarios, pue<strong>de</strong> llevar a un crecimiento mayor que<br />
el esperado por lo que aporta cada alimento por separado. En el caso <strong>de</strong>l hombre, el arroz y las<br />
arvejas constituyen ejemplos <strong>de</strong> recursos complementarios.<br />
Los recursos sustituibles son antagonistas (Fig c) si el organismo necesita una cantidad total<br />
proporcionalmente mayor <strong>de</strong> recursos cuando son consumidos juntos que cuando son consumidos<br />
por separado. Este efecto se pue<strong>de</strong> producir cuando hay un efecto sinérgico <strong>de</strong> compuestos tóxicos,<br />
y por más que cada recurso pueda ser utilizado por separado, y que los recursos puedan<br />
reemplazarse mutuamente, al ser consumidos juntos es necesaria una mayor cantidad total para<br />
obtener un mismo crecimiento.<br />
Sin embargo, es necesario recordar que tanto para el caso <strong>de</strong> los recursos perfectamente sustituibles,<br />
como los complementarios y antagonistas, la cantidad necesaria <strong>de</strong> un recurso disminuye si se<br />
28
Ecología General<br />
consume el otro (Figuras a, b y c). Lo que difiere es la proporcionalidad, en el primer caso son<br />
exactamente equivalentes, en el caso <strong>de</strong> los complementarios son más que equivalentes, y los<br />
antagonistas menos que equivalentes.<br />
R1<br />
a b c<br />
1.- La radiación como recurso<br />
La radiación solar es la fuente <strong>de</strong> energía que utilizan las plantas ver<strong>de</strong>s. La energía radiante llega<br />
hasta la planta en forma <strong>de</strong> radiación proce<strong>de</strong>nte <strong>de</strong>l sol, ya sea <strong>de</strong> modo directo o bien <strong>de</strong>spués <strong>de</strong><br />
haber sido difundida por la atmósfera o reflejada o transmitida por otros objetos. Cuando una hoja<br />
intercepta energía radiante, ésta pue<strong>de</strong> ser reflejada, transmitida o absorbida. Una parte <strong>de</strong> la<br />
fracción absorbida pue<strong>de</strong> llegar hasta los cloroplastos y activar el proceso <strong>de</strong> fotosíntesis. La energía<br />
radiante que ha sido fijada sólo pasa una vez por la tierra (es un flujo continuo). Esto es exactamente<br />
lo contrario <strong>de</strong> lo que ocurre con un átomo <strong>de</strong> nitrógeno o <strong>de</strong> carbono, o con una molécula <strong>de</strong> agua,<br />
que pue<strong>de</strong>n ciclar repetidamente a través <strong>de</strong> innumerables generaciones <strong>de</strong> organismos (cumplen<br />
ciclos biogeoquímicos).<br />
La radiación solar es un recurso continuo (un espectro <strong>de</strong> diferentes longitu<strong>de</strong>s <strong>de</strong> onda), pero<br />
el aparato fotosintético sólo es capaz <strong>de</strong> acce<strong>de</strong>r a la energía <strong>de</strong> una banda restringida <strong>de</strong> dicho<br />
espectro que se encuentra <strong>de</strong>ntro <strong>de</strong> las longitu<strong>de</strong>s <strong>de</strong> onda <strong>de</strong>l visible (entre 380 y 710 nm).<br />
La mayoría <strong>de</strong> las hojas <strong>de</strong> las plantas viven en un régimen <strong>de</strong> luz que varía a lo largo <strong>de</strong>l día<br />
y <strong>de</strong>l año, y en un medio ambiente con otras hojas que modifican la calidad y la cantidad <strong>de</strong> luz<br />
recibida. Esto ilustra dos propieda<strong>de</strong>s importantes <strong>de</strong> todos los recursos: su provisión pue<strong>de</strong> variar <strong>de</strong><br />
modo sistemático o pre<strong>de</strong>cible (variación <strong>de</strong> la intensidad luminosa <strong>de</strong>bida a los ritmos diarios y<br />
anuales <strong>de</strong> la radiación solar), o <strong>de</strong> modo no sistemático o impre<strong>de</strong>cible (variación <strong>de</strong> la intensidad<br />
luminosa recibida por una hoja causada por la naturaleza y la posición <strong>de</strong> las hojas vecinas). Las<br />
formas en que un organismo o un órgano reaccionan ante el abastecimiento pre<strong>de</strong>cible o<br />
impre<strong>de</strong>cible <strong>de</strong> un recurso reflejan su fisiología actual y su historia evolutiva.<br />
Las comunida<strong>de</strong>s vegetales terrestres (pastizales, bosques, arbustos o campos cultivados)<br />
están constituidas por uno o más estratos <strong>de</strong> vegetación. Cada estrato, cada planta y cada hoja<br />
intercepta la luz y crea así una zona <strong>de</strong> privación <strong>de</strong>l recurso. Así, por ejemplo, en un bosque <strong>de</strong>nso y<br />
pluriestratificado, las plantas <strong>de</strong>l sotobosque se encontrarán con una variación impre<strong>de</strong>cible <strong>de</strong>l<br />
recurso durante el transcurso <strong>de</strong> sus vidas (a medida que los árboles <strong>de</strong>l bosque crecen y mueren,<br />
creando y llenando vacíos en la cobertura <strong>de</strong>l bosque) e incluso durante el transcurso <strong>de</strong> un mismo<br />
día (a medida que cambia el ángulo <strong>de</strong> inci<strong>de</strong>ncia <strong>de</strong> los rayos solares).<br />
En los ecosistemas acuáticos (mares, lagos, lagunas), la energía radiante que llega es en<br />
parte absorbida y reflejada por el agua. En el océano la energía contenida en la fracción visible <strong>de</strong>l<br />
espectro disminuye un 50% respecto <strong>de</strong> la superficie a los 10 m <strong>de</strong> profundidad. El agua absorbe<br />
radiación <strong>de</strong> onda larga y virtualmente casi toda la radiación infrarroja <strong>de</strong>saparece en los primeros<br />
metros <strong>de</strong> profundidad. La absorción y reflexión <strong>de</strong> la luz por parte <strong>de</strong>l agua limita la profundidad a la<br />
cual las algas pue<strong>de</strong>n vivir. Por otro lado, las algas se ubican estratégicamente según la profundidad<br />
<strong>de</strong> acuerdo a la calidad <strong>de</strong> luz disponible y el tipo <strong>de</strong> pigmento accesorio para la fotosíntesis que<br />
poseen.<br />
2.- Nutrientes minerales<br />
Para que una planta pueda vivir, a<strong>de</strong>más <strong>de</strong> energía lumínica, dióxido <strong>de</strong> carbono y agua, necesita<br />
recursos minerales que <strong>de</strong>be obtener <strong>de</strong>l suelo o <strong>de</strong>l agua. Estos se clasifican en macronutrientes<br />
(necesarios en cantida<strong>de</strong>s relativamente elevadas): N, P, S, K, Ca, Mg y Fe, y elementos traza o<br />
micronutrientes: Mn, Zn, Cu, Bo, etc.<br />
R2<br />
29
Ecología General<br />
Cada uno <strong>de</strong> estos elementos penetra en la planta in<strong>de</strong>pendientemente, como ion o como<br />
molécula, y cada uno tiene sus propieda<strong>de</strong>s características <strong>de</strong> absorción y difusión, que afectan su<br />
capacidad <strong>de</strong> acceso a la planta. Las plantas adquieren los nutrientes minerales <strong>de</strong>l agua que está en<br />
contacto con sus raíces. La disponibilidad <strong>de</strong> estos elementos <strong>de</strong>pen<strong>de</strong> <strong>de</strong> la estructura molecular en<br />
la que se encuentran presentes en el suelo o en el agua, y <strong>de</strong> la temperatura, aci<strong>de</strong>z y presencia <strong>de</strong><br />
otros elementos.<br />
OBJETIVO<br />
El objetivo <strong>de</strong>l presente trabajo práctico es evaluar los efectos <strong>de</strong> distintas concentraciones <strong>de</strong><br />
nutrientes (recurso alimenticio) y niveles <strong>de</strong> iluminación (recurso energético) sobre el crecimiento<br />
poblacional <strong>de</strong> una especie <strong>de</strong> Lemnaceae (plantas acuáticas flotantes, comúnmente llamadas<br />
lentejas <strong>de</strong> agua) bajo condiciones controladas <strong>de</strong> laboratorio.<br />
• Formule la/s hipótesis que pondrá/n a prueba y las predicciones asociadas<br />
MATERIALES Y MÉTODOS<br />
Se utilizarán individuos <strong>de</strong> lentejas <strong>de</strong> aguas los que se colocarán en recipientes <strong>de</strong> plástico<br />
conteniendo distintas concentraciones <strong>de</strong> nutrientes y se los someterá a diferentes intensida<strong>de</strong>s<br />
lumínicas. Del recurso lumínico se probarán dos niveles (luz y sombra), mientras que <strong>de</strong>l recurso<br />
nutritivo se probarán cinco niveles (cuatro correspondientes a diluciones progresivas al 50% <strong>de</strong> la<br />
concentración anterior a partir <strong>de</strong> una solución madre <strong>de</strong> 25 ml <strong>de</strong> solución nutritiva BBM más 1 l <strong>de</strong><br />
agua bi<strong>de</strong>stilada, y uno sin nutrientes, conteniendo sólo agua bi<strong>de</strong>stilada).<br />
El BBM [Bold’s Basal Medium] es un medio <strong>de</strong> cultivo que contiene macro y micronutrientes<br />
minerales en agua <strong>de</strong>stilada estéril. Los macronutrientes son: nitrato <strong>de</strong> sodio; cloruros <strong>de</strong> calcio y <strong>de</strong><br />
sodio; fosfatos <strong>de</strong> potasio y <strong>de</strong> magnesio; sulfato férrico; hidróxido <strong>de</strong> boro. Los Micronutrientes son:<br />
sulfatos <strong>de</strong> zinc y <strong>de</strong> cobre; cloruro <strong>de</strong> manganeso, nitrato <strong>de</strong> cobalto y óxido <strong>de</strong> molib<strong>de</strong>no).<br />
Se formarán grupos <strong>de</strong> alumnos por comisión. Cada grupo trabajará con recipientes a los que se<br />
le agregarán 350 ml <strong>de</strong> BBM <strong>de</strong> una <strong>de</strong>terminada concentración a <strong>de</strong>finir por el docente (eg. 50%,<br />
25%, 12,5% ó 0%). Luego, a cada recipiente se le colocarán 10 fron<strong>de</strong>s <strong>de</strong> lenteja <strong>de</strong> agua<br />
(separados mediante una lupa <strong>de</strong> mano). Cada recipiente será cubierto con una tapa (perforada) para<br />
evitar una posible contaminación, y se lo expondrá a la luz directamente o cubierto por un papel que<br />
atenúe la radiación directa. Cada grupo <strong>de</strong>berá rotular su recipiente con los siguientes datos: turno,<br />
grupo, tratamiento (concentración <strong>de</strong> BBM y luz o sombra) y nivel inicial <strong>de</strong>l volumen <strong>de</strong>l<br />
liquido.<br />
• Discuta dón<strong>de</strong> y cómo se dispondrán espacialmente las ban<strong>de</strong>jas y en que medida la<br />
distribución seleccionada podría afectar los resultados <strong>de</strong>l experimento<br />
Una vez iniciado el experimento, y durante 45 días, los recipientes <strong>de</strong>ben ser revisados<br />
exactamente cada 7 días con la ayuda <strong>de</strong> una lupa <strong>de</strong> mano y una pinza, para:<br />
1. Controlar que el volumen <strong>de</strong> solución se mantenga constante. Si el volumen se reduce, agregarle<br />
agua <strong>de</strong>stilada, hasta recuperar el volumen original.<br />
2. Contar el número <strong>de</strong> individuos vivos y muertos en cada ban<strong>de</strong>ja. Se consi<strong>de</strong>rará como un<br />
individuo a cada fron<strong>de</strong>. Los fron<strong>de</strong>s ver<strong>de</strong>s o amarillos en menos <strong>de</strong> un 50% serán consi<strong>de</strong>rados<br />
como individuos vivos; los fron<strong>de</strong>s amarillos en más <strong>de</strong> un 50 % o marrones serán consi<strong>de</strong>rados<br />
como muertos. Estos últimos <strong>de</strong>ben ser retirados <strong>de</strong> la ban<strong>de</strong>ja.<br />
3. Escribir los resultados <strong>de</strong> cada análisis semanal en una planilla, que estará disponible y a la vista<br />
en el lugar en el que se dispongan los recipientes.<br />
El análisis <strong>de</strong> los datos y el efecto <strong>de</strong> los tratamientos se realizarán sobre la base <strong>de</strong> la<br />
información obtenida. Cuántas factores tiene este diseño? Son todos <strong>de</strong>l mismo tipo? Qué tipo <strong>de</strong><br />
ANOVA <strong>de</strong>bería emplearse para el análisis <strong>de</strong> los datos?<br />
30
Ecología General<br />
Para los objetivos <strong>de</strong> este práctico solo se utilizará la medición en t4 (última semana) y los datos se<br />
analizarán con un ANOVA <strong>de</strong> 2 factores. Si los datos no cumplen con los supuestos <strong>de</strong> distribución<br />
normal y <strong>de</strong> homocedasticidad, se aplicará una transformación.<br />
• Materiales que <strong>de</strong>ben traer los alumnos para realizar el presente Trabajo Práctico: lupa <strong>de</strong><br />
mano, pinza, guantes <strong>de</strong>scartables (1 par), papel ma<strong>de</strong>ra, tijera, marcador in<strong>de</strong>leble, cinta scotch<br />
(al menos un elemento cada cuatro alumnos).<br />
AGRADECIMIENTOS<br />
Una versión anterior <strong>de</strong> este práctico fue amablemente cedido por la Dra. María Busch <strong>de</strong>l<br />
Laboratorio <strong>de</strong> Ecología <strong>de</strong> Poblaciones.<br />
BIBLIOGRAFÍA<br />
Begon M, Harper JL y Townsend CR (1987) Ecología: individuos, poblaciones y comunida<strong>de</strong>s. Ed.<br />
Omega, Barcelona.<br />
Cochran WG y Cox GM (1957) Experimental <strong>de</strong>signs. 2nd. ed., Wiley, New York.<br />
Krebs CJ (1989) Ecological methodology. Harper Collins.<br />
Ricklefs RE (1993) The economy of nature. Freeman & Company.<br />
Winer BJ, Brown DR y Michaels KM (1991) Statistical principles in experimental <strong>de</strong>sign. McGraw-Hill,<br />
New York.<br />
Zar JH (1996) Bioestatistical analysis. 3rd. ed. Prentice-Hall, Upper Saddle River.<br />
31
Trabajo práctico 3<br />
ESTIMACIÓN DE ABUNDANCIA<br />
Ecología General<br />
(1) Conceptos generales sobre estimación <strong>de</strong> abundancia<br />
La abundancia <strong>de</strong> una población es un importante indicador <strong>de</strong> una serie <strong>de</strong> problemas ecológicos, <strong>de</strong><br />
carácter físico-ambiental, históricos, o <strong>de</strong> sus relaciones con otras poblaciones (Rabinovich, 1980). La<br />
mayor parte <strong>de</strong> las investigaciones ecológicas involucran la <strong>de</strong>scripción y/o explicación <strong>de</strong> patrones<br />
<strong>de</strong> distribución y abundancia <strong>de</strong> organismos (Walker y col., 2000).<br />
El nivel <strong>de</strong> abundancia <strong>de</strong> una población pue<strong>de</strong> expresarse a través <strong>de</strong> distinto tipo <strong>de</strong> indicadores:<br />
Indicadores absolutos:<br />
Densidad poblacional: Número <strong>de</strong> individuos/área o volumen. Ejemplo: número <strong>de</strong> guanacos por<br />
hectárea, número <strong>de</strong> diatomeas por litro <strong>de</strong> agua.<br />
Indicadores relativos:<br />
En casos en que es dificultoso <strong>de</strong>terminar el área o volumen correspondiente al número <strong>de</strong><br />
individuos, se recurre a una estimación <strong>de</strong> abundancia relativa, en la que se pon<strong>de</strong>ra el número <strong>de</strong><br />
individuos observados al esfuerzo <strong>de</strong> muestreo realizado. Ejemplo: nº <strong>de</strong> aves avistadas por hora,<br />
kilos <strong>de</strong> pescado obtenidos con cierta medida <strong>de</strong> red y cierto esfuerzo <strong>de</strong> captura (horas-hombrepotencia<br />
<strong>de</strong>l barco), nº <strong>de</strong> perros por persona, nº <strong>de</strong> roedores capturados por nº <strong>de</strong> trampas<br />
colocadas<br />
A su vez, estos indicadores podrán ser directos o indirectos según el conteo involucre a los individuos<br />
(estimador directo) o a signos <strong>de</strong> la presencia <strong>de</strong> los individuos (estimador indirecto) en los cuales se<br />
cuentan indicios tales como nidos, huellas, heces, que se pue<strong>de</strong>n referir a un área o volumen<br />
<strong>de</strong>terminado, o a un esfuerzo <strong>de</strong> muestreo.<br />
Muestreo aplicado a estimación <strong>de</strong> abundancia<br />
Existen diversos métodos para <strong>de</strong>terminar la abundancia <strong>de</strong> una población animal o vegetal. La<br />
manera más directa <strong>de</strong> <strong>de</strong>terminar el tamaño poblacional es la <strong>de</strong> contar a todos los individuos<br />
(censo) o a una parte <strong>de</strong> la población (muestra). El censo <strong>de</strong> una población pue<strong>de</strong> ser irrealizable por<br />
cuestiones <strong>de</strong> costo (tiempo, personal, dinero), interferencia o <strong>de</strong>strucción <strong>de</strong> la población,<br />
imposibilidad <strong>de</strong> acce<strong>de</strong>r a todos los individuos, y <strong>de</strong>strucción <strong>de</strong>l hábitat. En consecuencia, en la<br />
mayoría <strong>de</strong> los casos se recurre a técnicas <strong>de</strong> muestreo que permiten estimar el tamaño <strong>de</strong> la<br />
población a partir <strong>de</strong>l recuento <strong>de</strong> los individuos o sus productos en una serie <strong>de</strong> unida<strong>de</strong>s<br />
muestrales. En síntesis, el muestreo es preferible a los censos por cuestiones <strong>de</strong> costo, rapi<strong>de</strong>z, y<br />
posibilidad <strong>de</strong> contar con mayor número <strong>de</strong> datos para cada individuo.<br />
Los valores <strong>de</strong> los parámetros obtenidos a partir <strong>de</strong> un muestreo se caracterizan por su exactitud,<br />
precisión y sesgo. La exactitud es una medida <strong>de</strong> cuán dispersos se hallan las estimaciones <strong>de</strong>l valor<br />
real (=paramétrico), y <strong>de</strong>termina la idoneidad <strong>de</strong>l método <strong>de</strong> muestreo. La precisión es una medida <strong>de</strong><br />
cuán dispersos se hallan los valores estimados respecto <strong>de</strong> su valor esperado <strong>de</strong> acuerdo a los<br />
valores obtenidos a partir <strong>de</strong>l muestreo, por ejemplo la media; a mayor variabilidad entre<br />
observaciones menor precisión. La precisión <strong>de</strong>pen<strong>de</strong> <strong>de</strong>l número <strong>de</strong> muestras. El sesgo es la<br />
diferencia entre el valor real y el valor promedio <strong>de</strong> las estimaciones.<br />
El objetivo <strong>de</strong> un programa <strong>de</strong> muestreo es proveer una estimación poblacional con la mayor<br />
exactitud y precisión posible con relación a la cantidad <strong>de</strong> esfuerzo (= trabajo o costo) invertido. No<br />
existe un diseño <strong>de</strong> muestreo perfecto, sino que cada problema requerirá su propio diseño adaptado<br />
a la distribución y ciclo <strong>de</strong> vida <strong>de</strong>l organismo en cuestión. Por lo tanto, es condición indispensable<br />
estar profundamente familiarizado con el organismo que vamos a estudiar, tanto en lo que respecta a<br />
información preexistente como a experiencias a campo y laboratorio propias.<br />
En todo muestreo existe una serie <strong>de</strong> pasos necesarios:<br />
32
Ecología General<br />
1. Determinar los objetivos <strong>de</strong> la investigación.<br />
2. Definir la población objetivo (la que es <strong>de</strong> nuestro interés y sobre la que haremos inferencias) y la<br />
muestreada.<br />
3. Especificar los datos necesarios para evitar un exceso <strong>de</strong> información.<br />
4. Especificar el nivel <strong>de</strong> precisión <strong>de</strong>seado: se halla afectado por la variabilidad natural <strong>de</strong> lo que<br />
<strong>de</strong>seamos medir y los errores <strong>de</strong> medición <strong>de</strong>l método, entre otros factores.<br />
5. Definir los métodos <strong>de</strong> medición: se halla relacionada con la vali<strong>de</strong>z, el costo y la precisión.<br />
6. Definir la unidad <strong>de</strong> muestreo; por ejemplo, una hoja, o todas las hojas <strong>de</strong> una rama, o todas las<br />
hojas <strong>de</strong> un árbol. Si fuera posible, realizar una lista <strong>de</strong> todas las unida<strong>de</strong>s <strong>de</strong> muestreo posible,<br />
sin repetición ni superposición <strong>de</strong> las mismas.<br />
7. Definir el programa (o diseño) <strong>de</strong> muestreo: <strong>de</strong>terminado por las características biológicas y<br />
ecológicas <strong>de</strong> la población en estudio.<br />
8. Realizar una encuesta piloto para probar la metodología a campo y recabar estimaciones<br />
preliminares <strong>de</strong> media, varianza, etc., si no existen datos previos pertinentes que puedan ser<br />
usados.<br />
9. Organizar el trabajo <strong>de</strong> campo: incluye el entrenamiento y la supervisión <strong>de</strong>l personal que<br />
intervendrá.<br />
10. Resumen y análisis <strong>de</strong> datos: incluye la edición, corrección y eliminación <strong>de</strong> observaciones<br />
anómalas.<br />
11. Como regla general, toda muestra obtenida es una guía potencial para muestreos futuros.<br />
Veremos ahora en más <strong>de</strong>talle algunos <strong>de</strong> los ítems anteriores. El objetivo <strong>de</strong> la investigación<br />
incluye plantear claramente la hipótesis <strong>de</strong> investigación para po<strong>de</strong>r ponerla a prueba teniendo en<br />
cuenta las escalas temporal y espacial <strong>de</strong> trabajo.<br />
La selección <strong>de</strong>l área <strong>de</strong> estudio es necesariamente subjetiva y <strong>de</strong>pen<strong>de</strong> <strong>de</strong>l objetivo <strong>de</strong>l estudio, si<br />
éste es extensivo (cubriendo una gran área) o intensivo (<strong>de</strong>tallado con esfuerzos <strong>de</strong> muestreos<br />
frecuentes). El criterio <strong>de</strong> selección <strong>de</strong>be expresarse claramente puesto que los resultados y<br />
conclusiones son <strong>de</strong> aplicación inmediata al área <strong>de</strong> estudio y <strong>de</strong> extrapolación posible a otras áreas<br />
comparables.<br />
Respecto a cómo situar las unida<strong>de</strong>s <strong>de</strong> muestreo, se <strong>de</strong>fine como programa <strong>de</strong> muestreo al<br />
diseño bajo el cual se cuentan los organismos en unida<strong>de</strong>s muestrales <strong>de</strong> tamaño y número ya<br />
fijados. Aquí el aspecto esencial es asegurar que las muestras sean representativas <strong>de</strong> la población<br />
que se está muestreando.<br />
Los programas <strong>de</strong> muestreo más frecuentemente utilizados son el muestreo al azar simple, el<br />
muestreo al azar estratificado y el muestreo sistemático.<br />
(a) Muestreo al azar simple: se seleccionan n unida<strong>de</strong>s muestrales al azar entre N posibles unida<strong>de</strong>s,<br />
<strong>de</strong> tal manera que cada una <strong>de</strong> las posibles combinaciones <strong>de</strong> selección tenga la misma probabilidad<br />
<strong>de</strong> ser elegida. Lo que verda<strong>de</strong>ramente importa es el proceso <strong>de</strong> selección aleatorio más que el<br />
propio resultado; por ejemplo, es posible que luego <strong>de</strong> un muestreo aleatorio en un campo un 80% <strong>de</strong><br />
las muestras provengan <strong>de</strong> una <strong>de</strong> las mita<strong>de</strong>s <strong>de</strong>l campo, lo cual no invalida al muestreo si bien<br />
pue<strong>de</strong> alterar la representatividad <strong>de</strong> las muestras seleccionadas.<br />
La ubicación <strong>de</strong> las muestras se selecciona con números al azar extraídos <strong>de</strong> una tabla o mediante<br />
un programa <strong>de</strong> computación. Varios métodos frecuentemente usados por comodidad no son<br />
realmente aleatorios y <strong>de</strong>ben evitarse (e.g., arrojar un cuadrado hacia cualquier dirección luego <strong>de</strong><br />
dar vueltas sobre si mismo).<br />
El muestreo aleatorio simple presenta varios inconvenientes. En zonas heterogéneas el error <strong>de</strong><br />
muestreo es consi<strong>de</strong>rable; algunas porciones <strong>de</strong> la zona pue<strong>de</strong>n resultar sub o sobrerepresentadas;<br />
algunas unida<strong>de</strong>s <strong>de</strong> muestreo pue<strong>de</strong>n caer en sitios inaccesibles o muy <strong>de</strong>teriorados. Por ello, el<br />
muestreo aleatorio simple es excluido para el estudio <strong>de</strong> zonas muy extensas y heterogéneas ya que<br />
no se obtiene información sobre variaciones entre sus subdivisiones homogéneas dado que se<br />
promedian todos los datos. Esto ha propiciado el uso <strong>de</strong>l muestreo al azar estratificado y el muestreo<br />
sistemático.<br />
(b) Muestreo al azar estratificado: es preferible al muestreo al azar simple cuando el ambiente a<br />
muestrear es heterogéneo y la probabilidad <strong>de</strong> encontrar a los organismos es diferente en diferentes<br />
33
Ecología General<br />
porciones <strong>de</strong>l hábitat. Para aumentar la precisión <strong>de</strong> las estimaciones y disminuir los costos, se<br />
subdivi<strong>de</strong> el hábitat en estratos para que la muestra esté constituida por elementos <strong>de</strong> cada uno <strong>de</strong><br />
ellos; esto mejora la representatividad y precisión en comparación a un muestreo aleatorio simple. Un<br />
estrato es una porción <strong>de</strong> terreno (o subconjunto) <strong>de</strong> características homogéneas. Los estratos no se<br />
eligen al azar. Si la selección <strong>de</strong> unida<strong>de</strong>s muestrales en cada estrato es por muestreo aleatorio<br />
simple, entonces el procedimiento se <strong>de</strong>nomina muestreo aleatorio estratificado. La elección sobre<br />
qué aspecto usar para estratificar se basa en el sentido común y en una apreciación <strong>de</strong> los factores<br />
que pue<strong>de</strong>n afectar la magnitud <strong>de</strong> la variable que se estudia. Si se muestrean insectos plaga<br />
forestales, el diámetro <strong>de</strong>l tronco probablemente sea un factor relevante para estratificar el muestreo.<br />
En general, se subdivi<strong>de</strong> al área <strong>de</strong> estudio <strong>de</strong> forma tal que se minimiza la varianza <strong>de</strong> la <strong>de</strong>nsidad<br />
<strong>de</strong> individuos <strong>de</strong>ntro <strong>de</strong>l estrato, y por lo tanto se maximizan las diferencias entre estratos.<br />
Generalmente es suficiente usar entre 3 y 6 estratos.<br />
Si el tamaño <strong>de</strong> la muestra en cada estrato (nh) es proporcional al tamaño <strong>de</strong> cada estrato Nh, el<br />
muestreo se llama estratificado con asignación proporcional. En este caso, las medias <strong>de</strong>l muestreo<br />
aleatorio estratificado y <strong>de</strong>l muestreo aleatorio simple son iguales. Las fórmulas <strong>de</strong> cálculo <strong>de</strong> medias<br />
y varianzas se pue<strong>de</strong>n hallar en Rabinovich (1980) y Cochran (1963).<br />
(c) Muestreo sistemático: en estos muestreos las unida<strong>de</strong>s muestrales se or<strong>de</strong>nan <strong>de</strong> acuerdo a<br />
algún criterio espacial o temporal <strong>de</strong>l 1 a N, se toma al azar una unidad k, y luego se extrae un nuevo<br />
elemento repetidamente cada k unida<strong>de</strong>s hasta completar la muestra <strong>de</strong> tamaño n. El principal<br />
inconveniente <strong>de</strong>l muestreo sistemático es que inadvertidamente el k pue<strong>de</strong> coincidir con algún patrón<br />
regular <strong>de</strong>l ambiente o <strong>de</strong> los organismos, dando como resultado una estimación sesgada. El<br />
muestreo sistemático no es al azar y por eso es frecuentemente criticado bajo argumentos <strong>de</strong> pureza<br />
estadística. Los <strong>de</strong>fensores <strong>de</strong>l muestreo sistemático aducen ventajas <strong>de</strong> costo y representatividad <strong>de</strong><br />
toda el área <strong>de</strong> estudio con relación al muestreo aleatorio simple.<br />
NOTA: las referencias bibliográficas aparecen al final <strong>de</strong> los <strong>Trabajos</strong> <strong>Prácticos</strong> 3 y 4, ambos sobre estimación <strong>de</strong><br />
abundancia en poblaciones.<br />
Agra<strong>de</strong>cimientos: Agra<strong>de</strong>cemos al Dr. Ricardo Gürtler la primer versión <strong>de</strong> esta introducción teórica.<br />
34
Ecología General<br />
Trabajo Práctico 3<br />
(2) MÉTODOS PARA LA ESTIMACIÓN DE LA ABUNDANCIA DE LA<br />
VEGETACIÓN HERBÁCEA Y ARBÓREA<br />
INTRODUCCIÓN<br />
por Ricardo Vicari<br />
El Trabajo Práctico se <strong>de</strong>sarrollará en el área <strong>de</strong>clarada Reserva Natural Costanera Sur <strong>de</strong> la Ciudad<br />
<strong>de</strong> Buenos Aires, que compren<strong>de</strong> 350 ha.<br />
Origen <strong>de</strong>l Parque Natural Costanera Sur:<br />
En 1972 surge el proyecto <strong>de</strong> "ganarle" tierras al Río <strong>de</strong> la Plata con el objetivo <strong>de</strong> construir una<br />
ciudad satélite. El relleno comienza a realizarse en 1978, utilizando un sistema similar al <strong>de</strong> los<br />
pol<strong>de</strong>rs holan<strong>de</strong>ses, pero por diversos motivos se abandonó oficialmente en 1981. Des<strong>de</strong> el inicio <strong>de</strong><br />
las obras, plantas y animales colonizan el área en forma natural. El 5 <strong>de</strong> Junio <strong>de</strong> 1986, por<br />
or<strong>de</strong>nanza municipal se crea el Parque Natural y Zona <strong>de</strong> Reserva Ecológica Costanera Sur.<br />
Características ambientales <strong>de</strong> la Reserva:<br />
Los sustratos son <strong>de</strong> relleno, ya que los terrenos fueron ganados al río. El material presenta distintos<br />
orígenes: sedimentos <strong>de</strong>l Río <strong>de</strong> la Plata, tierra y material <strong>de</strong> <strong>de</strong>molición. En la reserva existen<br />
distintos tipos <strong>de</strong> comunida<strong>de</strong>s vegetales. Se encuentran presentes dos tipos <strong>de</strong> bosques: el sauzal y<br />
el alisal. El trabajo práctico se <strong>de</strong>sarrollará en el bosque <strong>de</strong> alisos. Éste presenta una fisonomía<br />
pluriestratificada. En el estrato arbóreo domina Tessaria integrifolia, en el arbustivo se encuentran:<br />
Ricinus communis, Ficus sp, Lantana sp, etc. y el estrato herbáceo está formado por parches<br />
dominados por distintas herbáceas y latifoliadas tales como: Tra<strong>de</strong>scantia sp., Solidago chilensis,<br />
He<strong>de</strong>ra helix, Solanum amygdalifolium, etc.<br />
Características ecológicas <strong>de</strong> Tessaria integrifolia:<br />
Tessaria integrifolia pertenece a la familia <strong>de</strong> las compuestas. Es un árbol <strong>de</strong> 10-12 m <strong>de</strong> altura,<br />
<strong>de</strong>ciduo, que pue<strong>de</strong> alcanzar hasta 30 cm <strong>de</strong> diámetro en su tronco. Su distribución abarca <strong>de</strong>s<strong>de</strong><br />
Colombia hasta la <strong>de</strong>sembocadura <strong>de</strong>l Río <strong>de</strong> la Plata; se lo encuentra asociado a las cuencas <strong>de</strong> los<br />
ríos andinos <strong>de</strong> la vertiente atlántica, como límite oriental. Recibe el nombre vernáculo <strong>de</strong> “aliso <strong>de</strong><br />
río” en la región mesopotámica y en el <strong>de</strong>lta o “palo bobo” en la región chaqueña. Forma masas<br />
boscosas en forma <strong>de</strong> franjas <strong>de</strong> extensa longitud en las zonas <strong>de</strong> inundación <strong>de</strong> numerosos ríos <strong>de</strong><br />
América. Según Cabrera (1976), las especies <strong>de</strong>l género Tessaria colonizan fácilmente los bancos <strong>de</strong><br />
arena <strong>de</strong>bido a que poseen raíces cundidoras, gemíferas, con multiplicación vegetativa, razón por la<br />
cual se propaga fácilmente formando colonias puras, a<strong>de</strong>más <strong>de</strong> dispersarse por semillas, ya que las<br />
mismas germinan fácilmente en las arenas húmedas <strong>de</strong> los bancos <strong>de</strong> los ríos formando verda<strong>de</strong>ros<br />
almácigos artificiales. Su distribución geográfica está fuertemente condicionada por las dinámicas<br />
geomorfológicas <strong>de</strong> las cuencas y por el régimen hidrosedimentológico <strong>de</strong> los ríos. La colonización y<br />
el afianzamiento <strong>de</strong> T. integrifolia y la posibilidad <strong>de</strong> crecer formando rodales <strong>de</strong>nsos están muy<br />
limitados con la ocurrencia <strong>de</strong> pulsos periódicos <strong>de</strong> inundación <strong>de</strong> los ríos. Su ma<strong>de</strong>ra, blanca y<br />
liviana, es usada para leña, cajonería, confección <strong>de</strong> papel y en la construcción <strong>de</strong> ranchos. Por la<br />
extensión que los bosques <strong>de</strong> alisos ocupan en la cuenca <strong>de</strong>l Plata, por su rápido crecimiento y<br />
productividad y por el aporte <strong>de</strong> materia orgánica al sistema fluvial, constituyen un subsistema <strong>de</strong><br />
gran importancia en la bioproductividad general <strong>de</strong> la cuenca y un recurso renovable <strong>de</strong> gran interés<br />
para el manejo ecológico <strong>de</strong> esa región. En la reserva se presenta formando dos masas boscosas<br />
próximas al Río <strong>de</strong> la Plata<br />
En este práctico nos centraremos en:<br />
A.- Estimación <strong>de</strong> la <strong>de</strong>nsidad <strong>de</strong> árboles por un método <strong>de</strong> área y otro <strong>de</strong> distancia<br />
A.1.- Métodos <strong>de</strong> área<br />
Consisten en la <strong>de</strong>limitación <strong>de</strong> un área que pue<strong>de</strong> tomar diversas formas (cuadrados, círculos, etc.) y<br />
dimensiones y que representa la unidad muestral. En cada una <strong>de</strong> las unida<strong>de</strong>s se registra el número<br />
<strong>de</strong> individuos. Como el área es conocida, es posible <strong>de</strong>terminar la <strong>de</strong>nsidad. La disposición espacial<br />
35
Ecología General<br />
<strong>de</strong> la población a muestrear <strong>de</strong>be ser consi<strong>de</strong>rada al momento <strong>de</strong> seleccionar el tamaño <strong>de</strong> unidad<br />
muestral (i.e. tamaño <strong>de</strong>l cuadrado). Si la disposición espacial <strong>de</strong> la población es al azar, el tamaño<br />
<strong>de</strong> la unidad muestral no alterará la exactitud <strong>de</strong> la estimación; su selección <strong>de</strong>pen<strong>de</strong>rá <strong>de</strong><br />
consi<strong>de</strong>raciones prácticas.<br />
A.2.- Métodos <strong>de</strong> distancia<br />
Se utilizan en general para individuos inmóviles o vegetación. Son un conjunto <strong>de</strong> métodos que no<br />
utilizan una superficie ó área <strong>de</strong> muestreo para obtener información <strong>de</strong> la abundancia, en inglés son<br />
<strong>de</strong>nominados “plotless methods”. Estas técnicas fueron <strong>de</strong>sarrolladas principalmente por el<br />
Laboratorio <strong>de</strong> Ecología Vegetal <strong>de</strong> Wisconsin. En estos métodos se trata <strong>de</strong> estimar, a través <strong>de</strong> la<br />
distancia media entre árboles, el área media ocupada por un individuo, a partir <strong>de</strong> la cual se<br />
<strong>de</strong>termina el número <strong>de</strong> individuos en un área dada. Estos métodos suponen que la disposición<br />
espacial <strong>de</strong> los individuos es al azar.<br />
La mayor ventaja <strong>de</strong> estimar el número <strong>de</strong> individuos a través <strong>de</strong> su distancia media en vez<br />
<strong>de</strong> utilizar el sistema más común <strong>de</strong> establecer áreas <strong>de</strong> muestreo, <strong>de</strong>ntro <strong>de</strong> las cuales se cuentan<br />
los individuos, radica en que se evita la marcación <strong>de</strong> los bor<strong>de</strong>s <strong>de</strong> los cuadrados. Esto en muchas<br />
situaciones salva una consi<strong>de</strong>rable cantidad <strong>de</strong> tiempo <strong>de</strong>bido a que las distancias entre los árboles<br />
son, generalmente, menores y más fáciles <strong>de</strong> medir que los bor<strong>de</strong>s <strong>de</strong> la parcela.<br />
Entre los métodos <strong>de</strong> distancia se encuentran algunos que utilizan mediciones <strong>de</strong> distancias<br />
entre plantas o <strong>de</strong>s<strong>de</strong> plantas a puntos elegidos en el terreno para estimar la distancia media. Los<br />
más utilizados son el método <strong>de</strong> los Pares al azar y el método <strong>de</strong> los cuartos. El último parece ser el<br />
mejor <strong>de</strong> los métodos <strong>de</strong> distancia por ser más fácil <strong>de</strong> aplicar y por ser aparentemente más eficiente,<br />
es una adaptación <strong>de</strong>l método usado por los topógrafos fe<strong>de</strong>rales <strong>de</strong> EEUU.<br />
El método <strong>de</strong> los cuartos es principalmente aplicable a comunida<strong>de</strong>s o estratos dominados<br />
por plantas leñosas arbóreas o arbustivas o a vegetación en la que los individuos se encuentran<br />
separados por gran<strong>de</strong>s espacios, sin embargo pue<strong>de</strong> adaptarse para otros tipos <strong>de</strong> estudios <strong>de</strong><br />
ecología vegetal o animal (por ejemplo para el cálculo <strong>de</strong> la <strong>de</strong>nsidad <strong>de</strong> bocas <strong>de</strong> cuevas <strong>de</strong><br />
animales excavadores).<br />
B.- Estimación <strong>de</strong> la cobertura <strong>de</strong> las especies <strong>de</strong>l sotobosque<br />
La cobertura <strong>de</strong> una especie u otra categoría vegetal, es la proporción <strong>de</strong> terreno ocupado por<br />
la proyección perpendicular <strong>de</strong> las partes aéreas <strong>de</strong> los individuos <strong>de</strong> las especies consi<strong>de</strong>radas. Se<br />
expresa como porcentaje <strong>de</strong> la superficie total. Esta medida es muy usada especialmente cuando la<br />
estimación <strong>de</strong> la <strong>de</strong>nsidad resulta difícil por la ausencia <strong>de</strong> límites netos entre los individuos, como<br />
ocurre en los pastizales, en plantas macollantes y cespitosas o en cojín. Para la estimación objetiva<br />
<strong>de</strong> la cobertura hay dos técnicas fundamentales: mediante unida<strong>de</strong>s muestrales lineales<br />
(interceptación lineal) y mediante unida<strong>de</strong>s muestrales puntuales.<br />
La primera técnica consiste en exten<strong>de</strong>r una línea <strong>de</strong> longitud (L) y medir la longitud (li)<br />
interceptada por la especie o categoría <strong>de</strong> interés. La cobertura <strong>de</strong> la especie xi es equivalente a la<br />
proporción <strong>de</strong> la longitud total interceptada por la especie consi<strong>de</strong>rada. La media y el <strong>de</strong>svío estándar,<br />
que permite estimar el error involucrado en la estimación, se pue<strong>de</strong>n obtener sobre la base <strong>de</strong> una<br />
serie <strong>de</strong> transectas lineales ubicadas al azar. Esta técnica resulta difícil <strong>de</strong> aplicar cuando los<br />
individuos están muy entremezclados, o cuando los espacios vacíos y ocupados forman un mosaico<br />
muy intrincado (Mateucci y Colma 1982).<br />
La técnica <strong>de</strong> estimar cobertura a partir <strong>de</strong> unida<strong>de</strong>s muestrales puntuales consiste en<br />
registrar la presencia o ausencia <strong>de</strong> una especie en una serie <strong>de</strong> puntos ubicados al azar. La<br />
proporción <strong>de</strong> puntos en los que la especie está presente equivale a la cobertura. Este diseño pue<strong>de</strong><br />
modificarse estimando la cobertura a partir <strong>de</strong> puntos ubicados sistemáticamente sobre una transecta<br />
(interceptación puntual). En este caso se registra el número <strong>de</strong> veces en que la especie vegetal es<br />
tocada por una aguja que se hace <strong>de</strong>scen<strong>de</strong>r verticalmente hacia la vegetación. La cobertura es el<br />
porcentaje que le correspon<strong>de</strong> a los “toques” positivos respecto <strong>de</strong>l total <strong>de</strong> puntos en la transecta. Al<br />
igual que en el caso anterior, si se ubican aleatoriamente varias <strong>de</strong> estas transectas, es posible<br />
obtener una estimación <strong>de</strong> la cobertura media y el <strong>de</strong>svío estándar.<br />
36
OBJETIVOS<br />
Ecología General<br />
El objetivo general es familiarizarse con distintas técnicas <strong>de</strong> muestreo para el estudio <strong>de</strong> poblaciones<br />
y comunida<strong>de</strong>s vegetales. El trabajo práctico se realizará en el alisal <strong>de</strong> Tessaria integrifolia ya que es<br />
la comunidad arbórea que ocupa una mayor extensión.<br />
Los objetivos específicos son:<br />
1) Estimar la <strong>de</strong>nsidad <strong>de</strong> alisos a través <strong>de</strong> dos métodos: uno <strong>de</strong> área y otro <strong>de</strong> distancia.<br />
2) Estimar la cobertura <strong>de</strong>l sotobosque en el bosque <strong>de</strong> alisos mediante dos métodos: uno <strong>de</strong><br />
interceptación lineal y otro <strong>de</strong> interceptación puntual.<br />
2) Determinar para cada método utilizado en 1 y 2 el costo en función <strong>de</strong>l tiempo y el grado<br />
<strong>de</strong> precisión a fin <strong>de</strong> seleccionar el más a<strong>de</strong>cuado según la relación costo/beneficio.<br />
DESARROLLO<br />
C.- Trabajo <strong>de</strong> campo<br />
C.1.- Mediciones en el estrato arbóreo<br />
• Materiales que <strong>de</strong>ben traer los alumnos para realizar el presente Trabajo Práctico<br />
• 5 estacas <strong>de</strong> ma<strong>de</strong>ra <strong>de</strong> 1’ x 1’<strong>de</strong> 1m <strong>de</strong> longitud<br />
• 1 cinta métrica <strong>de</strong> 10 m (o cinta métrica <strong>de</strong> 2 metros + una <strong>de</strong> las sogas <strong>de</strong> 10 m marcada cada<br />
10 cm)<br />
• 4 sogas <strong>de</strong> 10 m<br />
• tiras <strong>de</strong> plástico (para marcar los árboles)<br />
• fotocopia <strong>de</strong> las planillas <strong>de</strong> campo que se encuentran en la guía y lápiz<br />
• calculadora<br />
• repelente para mosquitos<br />
• ropa y calzado a<strong>de</strong>cuados<br />
• guantes<br />
Ubicación <strong>de</strong> las unida<strong>de</strong>s muestrales:<br />
En el bosque se realizará un muestreo al azar sobre rutas preestablecidas: se avanzará a lo largo <strong>de</strong>l<br />
camino principal y cada 20 m se seleccionarán al azar dos puntos, uno a la izquierda y el otro a la<br />
<strong>de</strong>recha <strong>de</strong>l sen<strong>de</strong>ro. Cada uno se ubicará sorteando un número (por ejemplo, 15), que indicará los<br />
metros que se <strong>de</strong>berá avanzar, hacia la izquierda o <strong>de</strong>recha <strong>de</strong>l camino principal, para ubicar el<br />
centro <strong>de</strong> la parcela. Cada punto establecido por el método aleatorio ya <strong>de</strong>scripto será el centro <strong>de</strong><br />
una parcela <strong>de</strong> 10 m x 10 m para el método areal y el punto para el no areal, método que será<br />
explicado mas a<strong>de</strong>lante.<br />
NOTA: Se evitarán los bor<strong>de</strong>s <strong>de</strong>l sen<strong>de</strong>ro. Si el número sorteado fuera pequeño y la parcela quedara<br />
<strong>de</strong>ntro <strong>de</strong> la zona disturbada cercana al camino, este número será <strong>de</strong>sechado y se sorteará otro.<br />
Cada comisión <strong>de</strong> TP ubicará 10 puntos <strong>de</strong> muestreo (en a<strong>de</strong>lante puntos) en cada uno <strong>de</strong> los cuales<br />
registrará la información que se <strong>de</strong>talla en a<strong>de</strong>lante.<br />
Registro <strong>de</strong> datos para el método no areal:<br />
Se utilizará el método <strong>de</strong> los cuartos para cuya realización se cumplirán los siguientes pasos: con<br />
cada punto como centro, se traza un par <strong>de</strong> coor<strong>de</strong>nadas ortogonales; se mi<strong>de</strong> la distancia entre el<br />
punto y los cuatro individuos más cercanos ubicados en cada uno <strong>de</strong> los cuatro cuadrantes como se<br />
indica en la figura. Por cada punto se obtienen cuatro distancias.<br />
IMPORTANTE: Un alumno registrará el tiempo <strong>de</strong>s<strong>de</strong> el momento en que se comienza a instalar las<br />
coor<strong>de</strong>nadas hasta que se termina la medición. Los datos se vuelcan en la planilla <strong>de</strong> campo.<br />
37
Ecología General<br />
NOTA: Cada árbol medido se marcará con una cinta plástica <strong>de</strong> color ya que este método requiere<br />
que se tengan en cuenta lo siguiente: (a) cada cuadrante <strong>de</strong>berá tener una medida <strong>de</strong> distancia; (b)<br />
no se <strong>de</strong>be medir la distancia a un mismo árbol <strong>de</strong>s<strong>de</strong> dos puntos <strong>de</strong> muestreo diferentes.<br />
Registro <strong>de</strong> datos para el método areal:<br />
En cada sitio seleccionado una vez <strong>de</strong>limitada la parcela con ayuda <strong>de</strong> las sogas y las estacas, se<br />
contarán todos los individuos <strong>de</strong> alisos cuyos troncos se encuentren <strong>de</strong>ntro <strong>de</strong> la parcela. Se<br />
consi<strong>de</strong>rarán los árboles mayores a 5 cm <strong>de</strong> diámetro (16 cm <strong>de</strong> perímetro).<br />
IMPORTANTE: Un alumno registrará el tiempo <strong>de</strong>s<strong>de</strong> el momento que se establece el punto hasta<br />
que se termine la <strong>de</strong>terminación <strong>de</strong> la <strong>de</strong>nsidad por el método areal, lo que significa que incluye el<br />
tiempo <strong>de</strong> la <strong>de</strong>limitación <strong>de</strong>l cuadrado. Los datos se vuelcan en la planilla <strong>de</strong> campo, al igual que el<br />
tiempo.<br />
C.2.- Mediciones en el sotobosque<br />
• Materiales que <strong>de</strong>ben traer los alumnos para realizar el presente Trabajo Práctico:<br />
• 1 cinta métrica <strong>de</strong> 10 m (o cinta métrica <strong>de</strong> 2 metros + una <strong>de</strong> las sogas <strong>de</strong> 10 m marcada cada<br />
10 cm)<br />
• 2 estacas<br />
• 1 aguja <strong>de</strong> tejer<br />
• fotocopia <strong>de</strong> las planillas <strong>de</strong> campo que se encuentran en la guía y lápiz<br />
Se utilizará el lado más alejado <strong>de</strong>l camino <strong>de</strong> cada cuadrado para esta estimación, que será utilizado<br />
como transecta para las estimaciones. A lo largo <strong>de</strong> la transecta se colocará la cinta métrica. Para el<br />
método <strong>de</strong> interceptación lineal, se registrará la longitud <strong>de</strong> la transecta interceptada por vegetación<br />
(herbácea latifoliada o herbácea graminiforme) o suelo <strong>de</strong>snudo. En el caso <strong>de</strong> interceptación puntual,<br />
se registrará con ayuda <strong>de</strong> la aguja <strong>de</strong> tejer, el número <strong>de</strong> puntos (<strong>de</strong> un total <strong>de</strong> 100, separados entre<br />
sí cada 10 cm) en los cuales se interceptó vegetación (herbácea latifoliada o herbácea graminiforme)<br />
y suelo <strong>de</strong>snudo. La información será registrada en planilla. Para cada método <strong>de</strong> interceptación<br />
(lineal y puntual) se calcularán y anotarán los totales (distancias sumadas o número <strong>de</strong> puntos) para<br />
cada una <strong>de</strong> las categorías consi<strong>de</strong>radas. En cada caso se tomará el tiempo involucrado en la<br />
obtención <strong>de</strong> la medición y se registrará en la planilla.<br />
D.- Trabajo <strong>de</strong> laboratorio<br />
D.1.- Cálculo <strong>de</strong> la <strong>de</strong>nsidad <strong>de</strong> alisos<br />
Método areal:<br />
Se expresa el número <strong>de</strong> individuos por unidad <strong>de</strong> superficie y se calcula la <strong>de</strong>nsidad total en Ind./ha.<br />
Se estima el <strong>de</strong>svío estándar y los límites <strong>de</strong> confianza para la media con un nivel <strong>de</strong> significación <strong>de</strong>l<br />
95%.<br />
Método <strong>de</strong> distancia:<br />
Distancias<br />
Árbol<br />
38
Ecología General<br />
Se calculará la <strong>de</strong>nsidad en individuos por hectárea (ind./ha) con la siguiente fórmula (Pollard 1971):<br />
don<strong>de</strong>:<br />
NP = Estimación <strong>de</strong> la <strong>de</strong>nsidad <strong>de</strong> la población por el método <strong>de</strong> los cuartos (ind./m 2 )<br />
n = Número <strong>de</strong> puntos al azar<br />
rij= Distancia <strong>de</strong>s<strong>de</strong> el punto al azar i hasta el organismo más cercano en el cuadrante j<br />
(j = 1, 2, 3, 4; i = 1, ..., n)<br />
La varianza <strong>de</strong> Np es:<br />
Y el error estándar <strong>de</strong> NP es:<br />
Los intervalos <strong>de</strong> confianza <strong>de</strong>l 95% para NP pue<strong>de</strong>n obtenerse con la siguiente fórmula<br />
cuando 4n > 30 (Seber 1982):<br />
Límite inferior para Nˆ es:<br />
Y el superior para Nˆ<br />
es:<br />
P<br />
P<br />
S<br />
2<br />
ˆ<br />
4(<br />
4n<br />
−1)<br />
=<br />
π<br />
N P<br />
2<br />
ˆ 2<br />
N P =<br />
4n<br />
− 2<br />
∑<br />
∑<br />
ij<br />
2 ( rij<br />
)<br />
( rij<br />
)<br />
16n<br />
−1<br />
−1.<br />
96<br />
π<br />
∑<br />
ij<br />
16n<br />
−1<br />
+ 1.<br />
96<br />
π<br />
ij<br />
2 ( rij<br />
)<br />
Nota: Tanto el valor Np como los valores <strong>de</strong> los límites <strong>de</strong>l intervalo <strong>de</strong> confianza <strong>de</strong>ben multiplicarse<br />
por 10.000 para obtener número <strong>de</strong> alisos/hectárea.<br />
Discuta comparativamente las estimaciones <strong>de</strong> la <strong>de</strong>nsidad obtenidas con los dos métodos.<br />
Distribución espacial <strong>de</strong> los alisos:<br />
Para analizar la distribución espacial <strong>de</strong> los alisos (regular, al azar o agrupada) se utilizará la relación<br />
varianza/media. Si la distribución es al azar la distribución <strong>de</strong>l número <strong>de</strong> alisos por parcela se<br />
ajustará a una Poisson y la relación varianza/media será igual a 1.<br />
Estos cálculos se realizarán con los datos sin transformar a árboles por hectárea.<br />
39
D.2.- Cálculo <strong>de</strong> la cobertura <strong>de</strong>l sotobosque<br />
Ecología General<br />
Interceptación lineal:<br />
Una vez registrada la longitud <strong>de</strong> interceptación <strong>de</strong> cada línea, la cobertura se estimará como el<br />
porcentaje <strong>de</strong> la longitud interceptada con respecto a la longitud total <strong>de</strong> la línea. Posteriormente se<br />
estimará la cobertura media y el <strong>de</strong>svío estándar usando la información <strong>de</strong> todos los turnos.<br />
Interceptación puntual:<br />
Una vez registrado el número <strong>de</strong> puntos “tocados” por el sotobosque, la cobertura se estimará como<br />
el porcentaje <strong>de</strong> “toques” con respecto al total <strong>de</strong> puntos consi<strong>de</strong>rados. La cobertura media y el<br />
<strong>de</strong>svío estándar se estimarán con la información <strong>de</strong> todos los turnos.<br />
D.3.- Cálculo <strong>de</strong> la eficiencia<br />
Para <strong>de</strong>terminar la máxima eficiencia al comparar los dos métodos <strong>de</strong> estimación <strong>de</strong> abundancia <strong>de</strong><br />
alisos y los dos métodos correspondientes a la estimación <strong>de</strong> la cobertura <strong>de</strong>l sotobosque, siguiendo<br />
la i<strong>de</strong>a <strong>de</strong> Wiegert (1962) se <strong>de</strong>be MINIMIZAR el producto entre los VALORES RELATIVIZADOS <strong>de</strong><br />
tiempo (Como indicador <strong>de</strong> COSTO) y el coeficiente <strong>de</strong> variación (Como indicador <strong>de</strong> la Precisión y<br />
VARIABILIDAD). En otras palabras el método mas eficiente es aquel que minimiza el producto entre<br />
variabilidad relativa y costo relativo (ver Parte 1)<br />
Ejemplo: Para el criterio <strong>de</strong> “elección” <strong>de</strong> los dos métodos utilizados en el sotobosque (el <strong>de</strong><br />
interceptación puntual y el lineal) <strong>de</strong>terminamos la eficiencia mediante el producto <strong>de</strong>l tiempo total (1)<br />
y el coeficiente <strong>de</strong> variación (2):<br />
(1) Tiempo = Promedio <strong>de</strong> los tiempos empleados en realizar cada uno <strong>de</strong> los muestreos<br />
(2) Coeficiente <strong>de</strong> variación (CV) promedio = promedio <strong>de</strong> los CV <strong>de</strong> las tres categorías (gramíneas,<br />
latifoliadas y suelo <strong>de</strong>snudo)<br />
Calculo <strong>de</strong> la eficiencia:<br />
Para los objetivos <strong>de</strong> este trabajo práctico, tomaremos como la técnica más ineficiente la que<br />
presente el mayor producto <strong>de</strong>l costo por la variabilidad (expresados como el coeficiente <strong>de</strong> variación)<br />
<strong>de</strong> los datos obtenidos<br />
BIBLIOGRAFÍA<br />
Krebs CJ (1989) Ecological methodology. Harper Collins, New York.<br />
Cochran WG (1963) Técnicas <strong>de</strong> muestreo. 2da. ed. Interamericana.<br />
Matteucci SD y Colma A (1982) Metodología para el estudio <strong>de</strong> la vegetación. Secretaría general <strong>de</strong><br />
la OEA, Washington, D.C.<br />
Pollard JH (1971) On distance estimators of <strong>de</strong>nsity in randomly distributed forests. Biometrics 27:991-<br />
1002.<br />
Rabinovich JE (1980) Introducción a la ecología <strong>de</strong> poblaciones animales. CECSA, México.<br />
Seber GAF (1982) The Estimation of Animal Abundance and Related Parameters, 2 nd ed. Griffin,<br />
London.<br />
Wiegert RG (1962) The selection of an optimum quadrat size for sampling the standing crop of<br />
grasses and forbs. Ecology 43:125-129.<br />
40
(3) PLANILLA CAMPO - TRABAJO PRACTICO # 3<br />
Turno: .<br />
Grupo: .<br />
B.1. Estimación <strong>de</strong> la <strong>de</strong>nsidad <strong>de</strong> árboles (Tessaria integrifolia)<br />
1. Método Areal: 2. Método No Areal<br />
Muestra #<br />
Alisos/100 m 2<br />
Hora inicial<br />
Hora final<br />
Tiempo (min.)<br />
B.2- Estimación <strong>de</strong> la cobertura <strong>de</strong> las especies <strong>de</strong>l sotobosque<br />
B.2-a. Interceptación lineal<br />
Hora inicial: Hora final: Tiempo: MINUTOS<br />
Longitud (cm)<br />
Gramineas<br />
Latifoliadas<br />
Suelo Desnudo<br />
Longitud (cm)<br />
Gramineas<br />
Latifoliadas<br />
Suelo Desnudo<br />
SUMATORIA<br />
Gramineas<br />
Latifoliadas<br />
Suelo Desnudo<br />
B.2-b. Interceptación puntual<br />
Hora inicial: Hora final: Tiempo: MINUTOS<br />
Muestra #<br />
Cuadrante<br />
C1<br />
C2<br />
C3<br />
C4<br />
Distancia (m)<br />
Hora inicial<br />
Ecología General<br />
Punto 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25<br />
Gramineas<br />
Latifoliadas<br />
Suelo Desnudo<br />
Punto 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50<br />
Gramineas<br />
Latifoliadas<br />
Suelo Desnudo<br />
Punto 51 52 53 54 55 56 57 58 59 60 61 62 63 64 65 66 67 68 69 70 71 72 73 74 75<br />
Gramineas<br />
Latifoliadas<br />
Suelo Desnudo<br />
Punto 76 77 78 79 80 81 82 83 84 85 86 87 88 89 90 91 92 93 94 95 96 97 98 99 100<br />
Gramineas<br />
Latifoliadas<br />
Suelo Desnudo<br />
SUMATORIA<br />
Gramineas<br />
Latifoliadas<br />
Suelo Desnudo<br />
Hora final<br />
Tiempo (min.)<br />
41
COMO LLEGAR A LA RESERVA COSTANERA SUR<br />
Entradas y colectivos:<br />
Ecología General<br />
Entrada por Brasil (Lola Mora)<br />
1 - líneas 2 (cartel amarillo) y 4 (cartel rojo)<br />
ambas Ramal Central Costanera<br />
2 - Alicia Moreau <strong>de</strong> Justo y EE.UU. líneas<br />
103, 111<br />
3 - Ing.Huergo y EE.UU. líneas 20, 129<br />
4 -en Paseo Colon y EE.UU. líneas 4, 33, 54,<br />
61, 62, 64, 74, 86, 93, 130, 143, 152, 159<br />
(800 mts.)<br />
42
Trabajo práctico 4<br />
Ecología General<br />
ESTIMACIÓN DE LA ABUNDANCIA EN POBLACIONES ANIMALES:<br />
COMPARACIÓN DE 3 MÉTODOS PARA EL CASO DE LA<br />
POBLACIÓN CANINA EN DOS ÁREAS DE LA CIUDAD DE BUENOS<br />
AIRES<br />
por Diana Rubel<br />
Objetivos Generales <strong>de</strong>l Trabajo Práctico<br />
- Familiarizar a los alumnos con algunos <strong>de</strong> los métodos <strong>de</strong> trabajo para estimar abundancia <strong>de</strong><br />
poblaciones animales, tomando una población animal urbana (Canis familiaris).<br />
- Ejercitar a los alumnos en el análisis <strong>de</strong> datos <strong>de</strong> campo.<br />
- Cumplir con los dos primeros objetivos <strong>de</strong>sarrollando una investigación <strong>de</strong> interés para la comunidad<br />
que integran.<br />
Objetivos específicos<br />
El Trabajo Práctico estará enfocado a:<br />
Evaluar la abundancia <strong>de</strong> la población canina en 2 áreas <strong>de</strong> la Ciudad <strong>de</strong> Buenos Aires <strong>de</strong><br />
diferente <strong>de</strong>nsidad poblacional humana.<br />
Introducir a los alumnos en la utilización <strong>de</strong> los métodos <strong>de</strong> estimación <strong>de</strong> abundancia por recuento<br />
<strong>de</strong> signos (heces), por marcado/recaptura y por conteo <strong>de</strong> individuos en una muestra aleatoria.<br />
Comparar los resultados <strong>de</strong> abundancia obtenidos por medio <strong>de</strong> los 3 métodos empleados y<br />
discutir sobre sus respectivos sesgos, ventajas y <strong>de</strong>sventajas.<br />
INTRODUCCIÓN TEÓRICA<br />
El perro doméstico (Canis familiaris) es la única especie doméstica <strong>de</strong> las 38 especies incluidas en la<br />
Familia Canidae. Los miembros <strong>de</strong>l género Canis (perros, lobos, chacales, coyotes, 9 especies) son<br />
interfértiles y comparten un pool genético casi idéntico, como se ha <strong>de</strong>mostrado utilizando técnicas<br />
genéticas y bioquímicas (Simonsen, 1976).<br />
Los estudios fósiles indican que hace aproximadamente 14.000 años el perro ya acompañaba a las<br />
poblaciones humanas, por lo que se consi<strong>de</strong>ra el primer animal doméstico (Clutton-Brock, 1995).<br />
Los procesos <strong>de</strong> urbanización han traído como consecuencia una asociación cada vez más íntima<br />
entre el hombre y el perro, lo que produjo reacciones contradictorias en nuestras poblaciones urbanas<br />
actuales: por un lado, se concibe el perro como un potencial enemigo (causante <strong>de</strong> mor<strong>de</strong>duras,<br />
contaminación <strong>de</strong> espacios públicos, transmisión <strong>de</strong> enfermeda<strong>de</strong>s 1 ) y por otro, recibe un trato casi<br />
humano (antropomorfismo), remarcando distinto tipo <strong>de</strong> beneficios asociados a su cría: psicológicos,<br />
terapéuticos o económicos (Serpell, 1995).<br />
El conocimiento y monitoreo <strong>de</strong> la <strong>de</strong>nsidad y la <strong>de</strong>mografía <strong>de</strong> la población canina – la domiciliada y<br />
la <strong>de</strong>nominada “callejera” – es necesario tanto para el diseño <strong>de</strong> campañas <strong>de</strong> vacunación, la<br />
planificación <strong>de</strong> campañas educativas, prevención <strong>de</strong> mor<strong>de</strong>duras y enfermeda<strong>de</strong>s transmisibles a la<br />
población humana, etc.<br />
1 Entre las numerosas infecciones transmitidas por los perros (más <strong>de</strong> <strong>de</strong> 70 según Acha y Szyfres, 1989) se<br />
encuentran helmintiasis (por ejemplo: toxocariasis, anquilostomiasis, filariasis, dipilidiasis). Los huevos <strong>de</strong> estos<br />
helmintos parásitos cosmopolitas se eliminan con las heces caninas y pue<strong>de</strong>n sobrevivir meses en condiciones<br />
a<strong>de</strong>cuadas.<br />
43
Ecología General<br />
La Ciudad <strong>de</strong> Buenos Aires cuenta con una única evaluación <strong>de</strong> abundancia <strong>de</strong> perros domiciliados.<br />
El método empleado fue el <strong>de</strong> una encuesta domiciliaria aleatoria y los resultados obtenidos indicaron<br />
que para 1994 la población canina porteña estaba entre 378.139 y 417.943 perros y una relación 7,45<br />
personas/perro (An<strong>de</strong>rson y col., 1996).<br />
El ambiente urbano tiene un alto nivel <strong>de</strong> heterogeneidad, por lo que es esperable que la abundancia<br />
<strong>de</strong> la población canina presente diferencias entre zonas <strong>de</strong> distintas características. Los datos<br />
<strong>de</strong>mográficos humanos tienen relevancia <strong>de</strong>bido a que la <strong>de</strong>nsidad canina suele presentar una<br />
relación directa con la <strong>de</strong>nsidad humana, y una relación inversa con el nivel socioeconómico (Matus<br />
y col., 1974; Agostini y col., 1980; An<strong>de</strong>rson y col., 1994). El nivel <strong>de</strong> contaminación fecal canina <strong>de</strong><br />
los espacios públicos (veredas, plazas) probablemente también esté asociado con la <strong>de</strong>nsidad<br />
canina, si bien no hay estudios que propongan un índice calibrado para ambas variables (Rubel y<br />
Wisnivesky, 2005).<br />
Evaluaciones realizadas alumnos <strong>de</strong> la Cátedra Parasitología General <strong>de</strong> la FCEyN (De<br />
Francesco y col., 1997, 1998) en las plazas <strong>de</strong> la Ciudad <strong>de</strong> Buenos Aires indicaron que:<br />
El número <strong>de</strong> perros que visitan diariamente las plazas varía entre 28 y 223<br />
La frecuencia media <strong>de</strong> visitas fue <strong>de</strong> 1,8 ± 0,1 veces/perro/día<br />
El número <strong>de</strong> perros que visitan diariamente las plazas no presentó fluctuaciones estacionales<br />
El 91% <strong>de</strong> los perros que visita las plazas porteñas resi<strong>de</strong> en viviendas que distan 500 metros<br />
como máximo <strong>de</strong> la plaza en cuestión<br />
El número <strong>de</strong> perros que visitan una plaza entre las 10:30 y las 12:30 hs. multiplicado por la<br />
Constante =3,38 es un estimador <strong>de</strong>l número total <strong>de</strong> perros que visitan la plaza (De Francesco y col.,<br />
1998)<br />
DESARROLLO DEL TRABAJO PRACTICO<br />
1- Diseño general <strong>de</strong> la investigación<br />
Se realizará un estudio transversal <strong>de</strong> abundancia <strong>de</strong> la población canina en dos áreas<br />
correspondientes a los barrios <strong>de</strong> Villa Ortúzar (CGP 11) y Belgrano (CGP 13) <strong>de</strong> la Ciudad <strong>de</strong><br />
Buenos Aires. Estas áreas tienen superficie equivalente y fueron seleccionadas por sus<br />
características diferenciales en cuanto a la <strong>de</strong>nsidad humana según la Zonificación <strong>de</strong>l Código <strong>de</strong><br />
Planeamiento Urbano <strong>de</strong>l Gobierno <strong>de</strong> la Ciudad <strong>de</strong> Buenos Aires.<br />
Las estimaciones se realizarán tomando como áreas <strong>de</strong> estudio una plaza <strong>de</strong> cada barrio y la zona<br />
circundante.<br />
Area Villa Ortúzar: Plaza 25 <strong>de</strong> agosto y área circundante. Zonificación R2b, que <strong>de</strong>nota zona<br />
resi<strong>de</strong>ncial general <strong>de</strong> <strong>de</strong>nsidad media-baja 2 , 790 hogares, 127.000 m 2 (ver Figura 1 en el ANEXO).<br />
2<br />
Todas las manzanas tienen entre 1 y 248 habitantes según datos <strong>de</strong> la Secretaría <strong>de</strong> Descentralización <strong>de</strong>l<br />
GCBA.<br />
44
Ecología General<br />
Area Belgrano: Plaza Alberti y área circundante. Zonificación R2a, que <strong>de</strong>nota zona resi<strong>de</strong>ncial<br />
general <strong>de</strong> <strong>de</strong>nsidad alta 3 , 2595 hogares, 182.000 m 2 (ver Figura 1 en el ANEXO).<br />
Se utilizarán tres métodos para estimar la abundancia <strong>de</strong> la población canina domiciliada en cada una<br />
<strong>de</strong> las áreas <strong>de</strong> estudio:<br />
a) recuento <strong>de</strong> signos (heces en la plaza);<br />
b) captura/recaptura (fotografiando los animales con un esfuerzo <strong>de</strong> muestreo prefijado <strong>de</strong> 2<br />
horas);<br />
c) recuento <strong>de</strong> animales por encuesta en puntos pre-fijados.<br />
Una vez obtenidos los datos, se analizarán las relaciones entre las abundancias estimadas para cada<br />
área con cada uno <strong>de</strong> los métodos empleados.<br />
Hipótesis:<br />
- La razón <strong>de</strong> abundancias entre áreas es aproximadamente constante para los tres métodos.<br />
- La abundancia <strong>de</strong> la población canina difiere entre las áreas<br />
2- Trabajo <strong>de</strong> campo. Metodología <strong>de</strong> recolección <strong>de</strong> datos<br />
El trabajo <strong>de</strong> campo se realizará durante una clase <strong>de</strong> <strong>Trabajos</strong> <strong>Prácticos</strong> en el horario <strong>de</strong> la clase.<br />
En cada una <strong>de</strong> las áreas <strong>de</strong> estudio trabajará la mitad <strong>de</strong> los alumnos <strong>de</strong>l TP. Para el trabajo <strong>de</strong><br />
campo los alumnos <strong>de</strong>l turno se dividirán en 6 grupos <strong>de</strong> igual cantidad <strong>de</strong> integrantes. Todos los<br />
asistentes llevarán guardapolvo los días <strong>de</strong> trabajo <strong>de</strong> campo sin excepción.<br />
Estim. <strong>de</strong> abundancia I – Recuento <strong>de</strong> signos<br />
El número <strong>de</strong> heces en la plaza se tomará como un estimador <strong>de</strong> abundancia canina en el área.<br />
Durante el trabajo <strong>de</strong> campo, se recorrerá sistemáticamente cada sector <strong>de</strong> la plaza contando las<br />
heces presentes. Los fragmentos cercanos entre sí y con igual aspecto se contabilizarán como una<br />
única feca. Los conteos se anotarán en el mapa <strong>de</strong> la plaza provisto por la cátedra (ver ANEXO).<br />
Estim. <strong>de</strong> abundancia II - Método <strong>de</strong> marcado/recaptura <strong>de</strong> Petersen<br />
La captura se realizará durante 2 horas en días diferentes (según el turno). El horario <strong>de</strong> captura será<br />
preferentemente <strong>de</strong>10:30 a 12:30 hs.<br />
Una pareja <strong>de</strong> alumnos se ubicará en cada vértice <strong>de</strong> la plaza y “capturará”, a través <strong>de</strong>l registro en<br />
una planilla, a todos los animales que ingresen por la cuadra asignada a esa esquina.<br />
Se consignarán los datos <strong>de</strong>l perro en la planilla provista por la cátedra (ingreso, la hora, el nombre<br />
<strong>de</strong>l perro preguntando a su dueño y su <strong>de</strong>scripción: raza, color, tamaño).<br />
Estim. <strong>de</strong> abundancia III: Recuento <strong>de</strong> animales en muestra aleatoria<br />
Se encuestará sobre la tenencia <strong>de</strong> perros (si/no) y su número a todas las personas que pasen por el<br />
punto fijado durante un intervalo <strong>de</strong> 90 minutos. Se preguntará la cuadra <strong>de</strong> resi<strong>de</strong>ncia para ubicarla<br />
en un croquis.<br />
Toda la información obtenida se consignará en las planillas provistas por la cátedra. Cuando no<br />
pueda obtenerse respuesta se consignará “no contesta” en la planilla correspondiente.<br />
3<br />
Todas las manzanas tienen entre 249 y 1082 habitantes según datos <strong>de</strong> la Secretaría <strong>de</strong> Descentralización <strong>de</strong>l<br />
GCBA.<br />
45
Elementos indispensables para el trabajo <strong>de</strong> campo:<br />
varias copias <strong>de</strong> las planillas y esquemas provistos por la<br />
cátedra, varios lápices y gomas <strong>de</strong> borrar<br />
planos <strong>de</strong> los espacios ver<strong>de</strong>s provistos por la cátedra<br />
carpeta para apoyar las planillas<br />
guardapolvo con cre<strong>de</strong>ncial provista por la cátedra<br />
TODAS LAS PLANILLAS DEBERÁN ENTREGARSE AL DOCENTE<br />
A CARGO EN LA PLAZA DEL AREA DE ESTUDIO<br />
ANTES DE RETIRARSE DEL TRABAJO PRÁCTICO<br />
Ecología General<br />
3- Análisis <strong>de</strong> los datos<br />
Una vez en el laboratorio, se confeccionarán las bases <strong>de</strong> datos en excel y se compararán<br />
posteriormente las abundancias entre áreas estimadas por cada método calculando el cociente:<br />
Abun<strong>de</strong>stim área Belgrano<br />
Abun<strong>de</strong>stim área V. Ort.<br />
Se discutirá si se cumple la hipótesis planteada.<br />
Estim. <strong>de</strong> abundancia I<br />
Se calculará la media <strong>de</strong> los censos <strong>de</strong> materia fecal realizados por cada turno.<br />
Estim. <strong>de</strong> abundancia II<br />
Recor<strong>de</strong>mos los supuestos <strong>de</strong>l método <strong>de</strong> Petersen:<br />
- La población es cerrada<br />
- Todos los animales tienen la misma probabilidad <strong>de</strong> ser capturados la primera vez<br />
- El marcado no afecta la capturabilidad (los animales marcados y no marcados tienen igual<br />
probabilidad <strong>de</strong> ser capturados)<br />
- Los animales no pier<strong>de</strong>n las marcas entre capturas<br />
El método se basa en la ecuación:<br />
Nestim = Capt2<br />
Marc1 Recapt<br />
Despejando, obtenemos:<br />
Nestim= Capt2.Marc1<br />
Recapt<br />
Nestim = población estimada<br />
Marc1 = animales capturados inicialmente y marcados<br />
Capt2 = animales capturados el segundo muestreo<br />
Recapt = animales capturados el segundo muestreo que presentan marca<br />
Una corrección con mejor ajuste cuando R >7 es:<br />
Nestim = Marc1 (Capt2 +1)<br />
Recapt +1<br />
46
Ecología General<br />
Se calcularán los límites <strong>de</strong> confianza siguiendo a Krebs (1989).<br />
Estim. <strong>de</strong> abundancia III<br />
Consi<strong>de</strong>rando las planillas obtenidas por todos los turnos, se calculará la distribución <strong>de</strong> frecuencias<br />
<strong>de</strong> hogares con 0, 1, 2, 3, 4 y 5 o más perros. En base a estos datos se testeará a qué distribución<br />
estadística ajusta la variable perros por hogar, y se calculará la media y el intervalo <strong>de</strong> confianza para<br />
los datos obtenidos.<br />
En base los resultados obtenidos, se estimará la <strong>de</strong>nsidad <strong>de</strong> la población canina en el área según<br />
los datos <strong>de</strong> número <strong>de</strong> hogares totales en el área (INDEC, Censo Nacional 2001) y la superficie <strong>de</strong> la<br />
misma en km 2 .<br />
Cuestionario y material complementario para reflexionar<br />
1- ¿Qué semejanzas y diferencias pue<strong>de</strong>n encontrar entre los perros <strong>de</strong> Buenos Aires y una<br />
población <strong>de</strong> animales no domésticos?<br />
2- ¿Con cuál/es <strong>de</strong> los métodos se obtiene el dato <strong>de</strong> <strong>de</strong>nsidad absoluta? ¿Qué aplicaciones<br />
secundarias pue<strong>de</strong> tener la estimación con cada uno <strong>de</strong> los métodos?<br />
3- ¿Qué sesgos pue<strong>de</strong>n señalarse para cada uno <strong>de</strong> los métodos y cómo podrían subsanarse?<br />
¿Qué frecuencias es necesario conocer o estimar para ajustar cada método? ¿Cómo<br />
modificarían el método II para estimar la abundancia <strong>de</strong> perros callejeros?<br />
4- ¿Qué estudio realizarían para calibrar un índice <strong>de</strong> <strong>de</strong>nsidad canina en función <strong>de</strong> <strong>de</strong>nsidad<br />
<strong>de</strong> heces en los espacios públicos?<br />
Artículos recomendados:<br />
Novaro AJ, Funes MC, Rambeaud C y Monsalvo O, 2000. Calibración <strong>de</strong>l índice <strong>de</strong> estaciones<br />
odoríferas para estimar ten<strong>de</strong>ncias poblacionales <strong>de</strong>l zorro colorado (Pseudalopex culpaeus)<br />
en Patagonia. Mastozoología Neotropical (SAREM) 7(2)81-88 (en castellano).<br />
Walker RS, Pancotto V, Schachter-Broi<strong>de</strong> J, Ackermann G y Novaro AJ, 2000. Evaluation of a fecalpellet<br />
in<strong>de</strong>x for abundance for mountain vizcachas (Lagidium viscacia) in Patagonia.<br />
Mastozoología Neotropical (SAREM) 7(2):89-94 (en inglés).<br />
Los artículos completos están disponibles en Internet en formato .pdf.<br />
Bibliografía<br />
Acha PN y Szyfres B, 1989. Zoonosis y enfermeda<strong>de</strong>s transmisibles comunes al hombre y a los<br />
animales, 2da. Edición, Organización Panamericana <strong>de</strong> la Salud.<br />
Agostini A, Franco A, Sommerfelt I, Arando <strong>de</strong> Lema J y Kistermann J, 1986. Aspectos <strong>de</strong> la<br />
<strong>de</strong>mografía canina y felina en el Partido <strong>de</strong>l Gral. San Martín, Provincia <strong>de</strong> Buenos Aires,<br />
1980. Rev. Med. Vet. 67:141-143.<br />
An<strong>de</strong>rson P, Beaudoin J, Castro J, González B, Landi P, Marcos E y Molina J, 1996. Relevamiento<br />
<strong>de</strong>mográfico <strong>de</strong> animales domésticos en la ciudad <strong>de</strong> Bs.As. 1994. Rev. Med. Vet. 77(3):206-<br />
212.<br />
Arredondo N, Vernetti M, Noriega E, Rubel D y Wisnivesky C. Vernetti, 2003. Evaluación <strong>de</strong>l riesgo <strong>de</strong><br />
transmisión <strong>de</strong> parásitos caninos en función <strong>de</strong> variables <strong>de</strong>mográficas, Buenos Aires. XVI<br />
47
Ecología General<br />
Congreso Latinoamericano <strong>de</strong> Parasitología, 1-3 <strong>de</strong> octubre, La Paz, Bolivia. Resumen<br />
183:82.<br />
Beck A, 1973. The Ecology of Stray Dogs. A Study of Free-Ranging Urban Animals. York Press,<br />
Baltimore.<br />
Clutton-Brock J, 1995. Origins of the dog: domestication and early history. In: The Domestic Dog: Its<br />
Evolution, behaviour and interactions with people. 1 st edition, edited by Serpell J, University<br />
Press, Cambridge, Great Britain.<br />
De Francesco MV, Zunino MG, Kuruc JA y Schweigmann N, 1997. Estimación <strong>de</strong>l tamaño <strong>de</strong> la<br />
población canina que visita los espacios ver<strong>de</strong>s porteños. XVIII Reunión Argentina <strong>de</strong><br />
Ecología, Bs. As.:36.<br />
De Francesco MV, Zunino MG, Kuruc JA, Rosselot C, Burroni N, Orellano P, Oliverio K, Jiménez L,<br />
Fontanarrosa S y Schweigmann N, 1998. Estimación <strong>de</strong> la población canina en los espacios<br />
ver<strong>de</strong>s porteños. II Congreso Argentino <strong>de</strong> Zoonosis y I Congreso Argentino y<br />
Latinoamericano <strong>de</strong> Enfermeda<strong>de</strong>s Emergentes, 13 al 17 abril, Buenos Aires. Resumen<br />
G3:230.<br />
De Francesco MV, Zunino MG, Kuruc JA y Schweigmann N, 1998. Estimación <strong>de</strong> la población canina<br />
en los espacios ver<strong>de</strong>s porteños. Artículo inédito.<br />
Krebs CJ, 1989. Ecological Methodology. HarperCollins Publishers, New York<br />
Matus PM, Morales MA, Loyola RC y Román DA, 1974. Estudio <strong>de</strong>mográfico <strong>de</strong> la población canina<br />
<strong>de</strong>l Gran Santiago. Rev. Soc. Med. Vet. <strong>de</strong> Chile XXIV:31-43.<br />
Rabinovich J, 1980. Introducción a la ecología <strong>de</strong> poblaciones animales. CECSA (Compañía Editorial<br />
Continental S.A.), México.<br />
Rubel D y Wisnivesky C, 2005. Magnitu<strong>de</strong> and distribution of canine faecal contamination and<br />
helminth eggs in two areas of different urban structure, Greater Buenos Aires, Argentina. Vet.<br />
Parasitol., 133: 339-347<br />
Serpell J, 1995. Dogs as human companions: a review of the relationship. In: The Domestic Dog: Its<br />
Evolution, behaviour and interactions with people. 1 st edition, edited by Serpell J, University<br />
Press, Cambridge, Great Britain.<br />
Walker RS, Novaro AJ y Nichols JD, 2000. Consi<strong>de</strong>raciones para la estimación <strong>de</strong> abundancia <strong>de</strong><br />
poblaciones <strong>de</strong> mamíferos. Mastozoología Neotropical / J.Neotrop.Mammal 7(2):73-80.<br />
48
Ecología General<br />
49
Trabajo Práctico Muestreo <strong>de</strong> poblaciones animales<br />
- Area <strong>de</strong> estudio Belgrano -<br />
Congreso<br />
Av. Cabildo<br />
Ugarte<br />
Arcos<br />
FFCC Mitre<br />
O’Higgins<br />
Roosvelt<br />
Monroe<br />
Ecología General<br />
N<br />
Escala aproximada 1:20.000<br />
50
Trabajo Práctico Muestreo <strong>de</strong> poblaciones animales<br />
- Area <strong>de</strong> estudio Villa Ortuzar -<br />
Tronador<br />
Giribone<br />
Heredia<br />
Charlone<br />
14 <strong>de</strong> Julio<br />
Guevara<br />
Elcano<br />
Ecología General<br />
N<br />
Escala aproxim.:1:20.000<br />
51
Nro<br />
PLANILLA CAPTURA/RECAPTURA - PLAZA ................................................ - FECHA: / /<br />
Hora<br />
Entrada<br />
Vereda calle ......................................<br />
Nombre<br />
Sexo<br />
(M/H)<br />
Raza<br />
Color/es<br />
Tamaño<br />
(chico,<br />
mediano,<br />
gran<strong>de</strong>)<br />
Recaptura<br />
(S/N)<br />
Observaciones
PLANILLA ENCUESTA EN EL SITIO............................................................... …………………………<br />
BARRIO ......................... - FECHA: / /<br />
¿Tiene<br />
perro?<br />
(si – no)<br />
¿A cuántas<br />
cuadras <strong>de</strong><br />
aquí vive?<br />
Nº perros<br />
(0-1-2-3)<br />
OBSERVACIONES (No contesta, etc.)
Trabajo práctico 5<br />
TABLAS DE VIDA<br />
Parte 1<br />
Métodos para la estimación <strong>de</strong> la edad<br />
54<br />
Ecología General<br />
por David Bilenca<br />
La <strong>de</strong>terminación <strong>de</strong> la edad <strong>de</strong> los individuos <strong>de</strong> una población constituye un parámetro fundamental<br />
para cualquier estudio ecológico. Ello es <strong>de</strong> particular importancia en los estudios <strong>de</strong>mográficos, en<br />
los cuales resulta indispensable establecer el número <strong>de</strong> individuos <strong>de</strong> la población que se encuentra<br />
en cada clase <strong>de</strong> edad [ a(x) ] como paso previo a la construcción <strong>de</strong> tablas <strong>de</strong> vida y el cálculo <strong>de</strong><br />
sus diversos parámetros (mortalidad, reproducción, esperanza <strong>de</strong> vida, etc.).<br />
Son diversos los métodos empleados para la <strong>de</strong>terminación <strong>de</strong> la edad en los seres vivos,<br />
<strong>de</strong>pendiendo en buena medida <strong>de</strong> las características <strong>de</strong> la especie.<br />
Las poblaciones naturales no viven en un ambiente constante, y tales fluctuaciones <strong>de</strong>l<br />
ambiente se reflejan en el organismo a través <strong>de</strong> cambios en su tasa <strong>de</strong> crecimiento. Buena parte <strong>de</strong><br />
los métodos para estimar la edad se basan en el aprovechamiento <strong>de</strong>l registro ontogenético <strong>de</strong> los<br />
cambios sufridos por el individuo que quedan conservados fundamentalmente en los tejidos duros<br />
como "marcas <strong>de</strong> crecimiento". Así, por ejemplo, la <strong>de</strong>terminación <strong>de</strong> la edad en el thar <strong>de</strong>l Himalaya<br />
se realiza mediante el recuento <strong>de</strong> anillos <strong>de</strong> crecimiento en los cuernos (Caughley 1977), en la perca<br />
amarilla se cuentan las marcas <strong>de</strong> crecimiento presentes en las escamas (Hen<strong>de</strong>rson 1985) y en la<br />
ballena piloto se cuentan las bandas <strong>de</strong> <strong>de</strong>ntina (Crespo et al. 1985). El recuento <strong>de</strong> anillos <strong>de</strong><br />
crecimiento para la <strong>de</strong>terminación <strong>de</strong> la edad es ampliamente utilizada en especies <strong>de</strong> árboles, y se<br />
conoce como <strong>de</strong>ndrocronología.<br />
Otros métodos para la <strong>de</strong>terminación <strong>de</strong> la edad se basan en medir estructuras que tengan un<br />
crecimiento continuo y lo más prolongado posible durante la vida <strong>de</strong>l individuo aún cuando no <strong>de</strong>jen<br />
"marcas <strong>de</strong> crecimiento". Previamente a la aplicación <strong>de</strong> estos métodos <strong>de</strong>be conocerse la relación<br />
entre el estimador a emplear y la edad <strong>de</strong>l individuo, para lo cual se efectúan curvas <strong>de</strong> calibración<br />
con individuos <strong>de</strong> edad conocida. En numerosas especies <strong>de</strong> micromamíferos, particularmente en<br />
roedores, el peso <strong>de</strong>l cristalino representa un buen estimador <strong>de</strong> la edad durante los primeros 3-4<br />
meses <strong>de</strong> vida (Lord 1959, Kozakiewicz 1976); a partir <strong>de</strong> esa edad en a<strong>de</strong>lante suele utilizarse como<br />
estimador al largo <strong>de</strong> las raíces <strong>de</strong> los molares (Pucek y Zejda 1968). Entre las aves, el largo y<br />
diámetro <strong>de</strong>l cálamo <strong>de</strong> las plumas primarias son utilizados como estimadores <strong>de</strong> edad (Szuba y<br />
Ben<strong>de</strong>ll 1987). En otros casos, en lugar <strong>de</strong> utilizarse al crecimiento como estimador <strong>de</strong> la edad se<br />
emplea al grado <strong>de</strong> <strong>de</strong>sgaste <strong>de</strong> las estructuras. El grado <strong>de</strong> <strong>de</strong>sgaste <strong>de</strong> los molares es una técnica<br />
ampliamente utilizada para estimar la edad en roedores (Pearson 1967, Zuleta et al. 1988).<br />
Buena parte <strong>de</strong> las técnicas para <strong>de</strong>terminar la edad hasta aquí enunciadas requieren<br />
sacrificar a los individuos estudiados, lo que en muchos casos resulta incompatible con los objetivos<br />
<strong>de</strong> estudios ecológicos que requieren un seguimiento <strong>de</strong> la población en el tiempo. En estos casos, la<br />
edad pue<strong>de</strong> estimarse indirectamente mediante el registro <strong>de</strong> medidas externas <strong>de</strong> los individuos<br />
(peso, largo <strong>de</strong>l cuerpo, largo <strong>de</strong>l tarso, etc.) cuya asociación con estimadores más precisos <strong>de</strong> la<br />
edad es conocida. Alternativamente, una técnica usada particularmente en aves y reptiles consiste en<br />
el seguimiento mediante muestreos sucesivos <strong>de</strong> individuos cuya fecha <strong>de</strong> nacimiento pudo ser<br />
establecida empíricamente mediante la inspección <strong>de</strong> nidos (Perrins 1965, Bellocq 1988).<br />
Finalmente, cabe <strong>de</strong>stacar que si bien los mo<strong>de</strong>los basados en la teoría clásica <strong>de</strong> la<br />
<strong>de</strong>mografía (Euler 1760, Sharpe y Lotka 1911), partiendo <strong>de</strong> la premisa <strong>de</strong> que las características <strong>de</strong><br />
la historia <strong>de</strong> vida <strong>de</strong> los individuos (natalidad, mortalidad) son <strong>de</strong>terminadas fundamentalmente por la<br />
edad, han sido muy útiles en la predicción <strong>de</strong> la dinámica <strong>de</strong> numerosas poblaciones, se ha<br />
encontrado con frecuencia que factores tales como el tamaño corporal <strong>de</strong> un individuo (Kirkpatrick
55<br />
Ecología General<br />
1984), la cohorte a la que pertenece (Kravetz et al. 1981, Zuleta et al. 1988), o su estado fisiológico<br />
pue<strong>de</strong>n tener un impacto mayor que su edad absoluta en <strong>de</strong>terminar los patrones <strong>de</strong> natalidad y<br />
mortalidad. Es por este motivo que en ciertos casos se utilizan "eda<strong>de</strong>s fisiológicas" en lugar <strong>de</strong><br />
eda<strong>de</strong>s absolutas para la construcción <strong>de</strong> tablas <strong>de</strong> vida. Las eda<strong>de</strong>s fisiológicas son frecuentemente<br />
utilizadas en poblaciones <strong>de</strong> insectos. Tal es el caso <strong>de</strong> Anopheles darlingi, un mosquito transmisor<br />
<strong>de</strong>l paludismo, en el cual la edad fisiológica en las hembras es estimada a partir <strong>de</strong>l número <strong>de</strong><br />
oviposiciones previas (Charlwood y Wilkes 1979).<br />
REFERENCIAS<br />
Bellocq MI (1988) Predación <strong>de</strong> roedores por aves en ecosistemas agrarios. Tesis Doctoral, FCEyN,<br />
UBA, Buenos Aires.<br />
Caughley G (1977) Analysis of vertebrate populations. Wiley, New York.<br />
Charlwood JD y Wilkes TJ (1979) Studies on the age-composition of samples of Anopheles darlingi<br />
Root (Diptera: Culicidae) in Brazil. Bull. Ent. Res. 69: 337-342.<br />
Crespo EA, Pagnoni G y Pedraza SN (1985) Structure of a long-finned pilot whale school stran<strong>de</strong>d in<br />
Patagonia. Sci. Rep. Whales Res. Inst. 36:97-106.<br />
Euler L (1760) Recherches generales sur la mortalité et la multiplication. Mémoires <strong>de</strong> l'Aca<strong>de</strong>mie<br />
Royale <strong>de</strong>s Sciences et Belles Lettres 16:144-164.<br />
Hen<strong>de</strong>rson BA (1985) Factors affecting growth and recruitment of yellow perch, Perca flavescens<br />
Mitchill, in South Bay, Lake Huron. J. Fish Biol. 26:449-458.<br />
Kirkpatrick M (1984) Demographic mo<strong>de</strong>ls based on size, not age, for organisms with in<strong>de</strong>terminate<br />
growth. Ecology 65:1874-1884.<br />
Kozakiewicz M (1976) The weight of eye lens as the proposed age indicator of the bank vole. Acta<br />
Theriologica 21:314-316.<br />
Kravetz FO, Manjon MC, Busch M, Percich RE, Marconi PN y Torres MP (1981) Ecología <strong>de</strong> Calomys<br />
laucha (Ro<strong>de</strong>ntia, Cricetidae) en el <strong>de</strong>partamento Río Cuarto (Córdoba). I. Dinámica <strong>de</strong> la<br />
población. Ecología 6:15-22.<br />
Lord RD (1959) The lens as an indicator of age in the cottontail rabbits. J. Wildl. Manage. 23:358-360.<br />
Pearson OP (1967) La estructura por eda<strong>de</strong>s y la dinámica reproductiva <strong>de</strong> una población <strong>de</strong>l roedor<br />
<strong>de</strong> campo Akodon azarae. Physis 27:53-58.<br />
Perrins CM (1965) Population fluctuations and clutch size in the great tit, Parus major L. J. Anim. Ecol.<br />
34:601-647.<br />
Pucek Z y Zejda J (1968) Technique for <strong>de</strong>termining age in the red-backed vole, Clethrionomys<br />
glareolus (Scheber 1780). Small Mammal Newslett. 2:51-60.<br />
Sharpe FR y Lotka AJ (1911) A problem in age-<strong>de</strong>termination. Philosophical Magazine, Series 6<br />
21:435-438.<br />
Szuba KJ y Ben<strong>de</strong>ll JF (1987) Age <strong>de</strong>termination of Hudsonian spruce grouse using primy feathers.<br />
Wildl. Soc. Bull. 15:539-543.<br />
Zuleta GA, Kravetz FO, Busch M y Percich RE (1988) Dinámica poblacional <strong>de</strong>l ratón <strong>de</strong>l pastizal<br />
pampeano (Akodon azarae), en ecosistemas agrarios <strong>de</strong> Argentina. Revista Chilena <strong>de</strong> Historia<br />
Natural 61:231-244.
Parte 2<br />
Tablas <strong>de</strong> vida<br />
56<br />
Ecología General<br />
por Ricardo Gürtler<br />
OBJETIVOS<br />
Compren<strong>de</strong>r y manejar los parámetros que componen una tabla <strong>de</strong> vida y sus fórmulas <strong>de</strong> cálculo<br />
para poblaciones <strong>de</strong> animales y plantas que exhiben ciclos <strong>de</strong> vida anuales, bianuales y plurianuales,<br />
<strong>de</strong> reproducción discreta o continua.<br />
DEFINICIONES<br />
* Cohorte: Grupo <strong>de</strong> individuos que pertenecen a la misma clase <strong>de</strong> edad.<br />
* Tiempo generacional: Período que transcurre en promedio entre el nacimiento <strong>de</strong> una madre y el<br />
nacimiento <strong>de</strong> su <strong>de</strong>scen<strong>de</strong>ncia.<br />
* Camada: número <strong>de</strong> nacidos en promedio por hembra.<br />
* Tabla <strong>de</strong> vida <strong>de</strong> cohortes: (= horizontal, específica por eda<strong>de</strong>s): se sigue a lo largo <strong>de</strong>l tiempo el<br />
<strong>de</strong>stino <strong>de</strong> un grupo <strong>de</strong> individuos que ha nacido en la misma época (cohorte).<br />
* Tabla <strong>de</strong> vida estática: (= vertical, tiempo-específica): se obtiene a partir <strong>de</strong> la estructura <strong>de</strong> eda<strong>de</strong>s<br />
<strong>de</strong> una población observada en un momento dado. Bajo la suposición <strong>de</strong> que la población se hallaba<br />
en estado estacionario entre la menor y mayor edad observada, se consi<strong>de</strong>ra que la muestra recogida<br />
constituye el <strong>de</strong>stino <strong>de</strong> una cohorte imaginaria.<br />
Ambas tablas <strong>de</strong> vida contienen los mismos parámetros:<br />
ax = número <strong>de</strong> individuos al inicio <strong>de</strong> la edad x.<br />
lx = probabilidad <strong>de</strong> supervivencia <strong>de</strong>s<strong>de</strong> la edad 0 a la edad x. (ax / a0)<br />
Por <strong>de</strong>finición, lx=0 = 1,000 o 1000.<br />
px = probabilidad <strong>de</strong> supervivencia entre x y x+1 = 1 - qx = lx+1 / lx .<br />
Es la probabilidad <strong>de</strong> terminar el intervalo (x, x+1) que tienen aquellos que estaban vivos al comienzo<br />
<strong>de</strong>l mismo. En tablas <strong>de</strong> vida vertical, se la refiere como tasa <strong>de</strong> supervivencia. En la última edad, px = 0.<br />
dx = proporción (o número) <strong>de</strong> individuos muertos <strong>de</strong> la cohorte original durante el intervalo <strong>de</strong> edad<br />
(x,x+1) = lx - lx+1 . En la última edad dx = lx .<br />
qx = probabilidad <strong>de</strong> mortalidad entre x y x+1 = dx / lx = 1 - (lx+1 / lx ) = 1 - px .<br />
Es la probabilidad <strong>de</strong> morir durante el intervalo (x, x+1) que tienen aquellos que se hallaban vivos al<br />
comienzo <strong>de</strong>l intervalo. En el contexto <strong>de</strong> una tabla <strong>de</strong> vida vertical, se la refiere como la tasa <strong>de</strong><br />
mortalidad.<br />
kx = killing-power = potencia <strong>de</strong> la mortalidad = log10 (ax / ax+1 ) = log10 ax - log10 ax+1 .<br />
Fx = fecundidad <strong>de</strong> la edad x. Es la cantidad total <strong>de</strong> <strong>de</strong>scendientes que <strong>de</strong>jan todos los individuos <strong>de</strong><br />
la edad x.<br />
mx = fecundidad medida como número promedio <strong>de</strong> crías totales por individuo o <strong>de</strong> crías hembra<br />
nacidas por hembra <strong>de</strong> edad x.<br />
Ro = tasa <strong>de</strong> reproducción (<strong>de</strong> reemplazo) básica = Σ lxmx .<br />
Tc = tiempo generacional <strong>de</strong> la cohorte = Σ (xlxmx) / R0 Es un promedio pon<strong>de</strong>rado que indica la edad<br />
promedio a la que los individuos tienen su <strong>de</strong>scen<strong>de</strong>ncia promedio.
= tasa instantánea <strong>de</strong> crecimiento poblacional =(log e Ro) / T c<br />
e x = expectativa <strong>de</strong> vida <strong>de</strong> la clase <strong>de</strong> edad x.<br />
última edad<br />
e x = Σ l y / l x<br />
y = x<br />
*Existen otras fórmulas para calcular la expectativa <strong>de</strong> vida<br />
Vx = valor reproductivo o expectativa <strong>de</strong> futura <strong>de</strong>scen<strong>de</strong>ncia.<br />
Vx = Σ ( ly / lx ) my, <strong>de</strong>s<strong>de</strong> y=x hasta la ultima edad.<br />
EJEMPLO DE UNA TABLA DE VIDA ARTIFICIAL PARA ILUSTRAR LOS CÁLCULOS.<br />
.<br />
x l(x) d(x) q(x) e(x)<br />
.<br />
0-1 1000 750 75 1.30<br />
1-2 250 210 84 1.22<br />
2-3 40 30 75 1.35<br />
3-4 10 7 70 1.40<br />
4-5 3 2 67 1.33<br />
5-6 1 1 100 1.00<br />
6-7 0 - - -<br />
TAREA<br />
57<br />
Ecología General<br />
A continuación se enuncian tres ejemplos <strong>de</strong> tablas <strong>de</strong> vida para distintas poblaciones específicas. En<br />
cada caso:<br />
(1) Calcule las columnas sin datos y grafique log10 (l(x).1000), px, qx y ex en función <strong>de</strong> la edad o<br />
estadio.<br />
(2) Calcule R0, T c y r.<br />
EJEMPLO 1: TABLA DE VIDA DE COHORTE PARA UN INSECTO ANUAL, EL SALTAMONTES<br />
Chortippus brunneus (Richards y Waloff 1954)<br />
En Ascot, Inglaterra, se estudió una población aislada <strong>de</strong> saltamontes en la cual se siguió el <strong>de</strong>stino<br />
<strong>de</strong> cada individuo <strong>de</strong>s<strong>de</strong> su nacimiento hasta su muerte. Las ninfas 1 emergen al principio <strong>de</strong> la<br />
primavera, en marzo, y llegan a adultos en agosto; las hembras ponen los huevos en el suelo y<br />
mueren todas a mediados <strong>de</strong> noviembre. Los huevos siguen el <strong>de</strong>sarrollo hasta la primavera siguiente<br />
en la cual eclosionan y constituyen la nueva generación.
Estadio (x) a(x) l(x) log10 (l(x).1000) d(x) q(x) p(x) F(x) m(x)<br />
huevo 44000 0 0<br />
ninfa 1 3513 0 0<br />
ninfa 2 2529 0 0<br />
ninfa 3 1922 0 0<br />
ninfa 4 1461 0 0<br />
adulto 1300 22617 17<br />
58<br />
Ecología General<br />
EJEMPLO 2: TABLA DE VIDA DE COHORTES PARA LA GRAMÍNEA Poa annua (Law 1975, en<br />
Begon et al. 1987)<br />
Cada individuo fue i<strong>de</strong>ntificado y mapeado <strong>de</strong>ntro <strong>de</strong> un campo, y su <strong>de</strong>stino fue seguido hasta la<br />
muerte <strong>de</strong> la última planta.<br />
Edad(x)en<br />
trimestres<br />
a(x) l(x) log10(l(x).1000) d(x) q(x) p(x) m(x)<br />
0 843 0<br />
1 722 300<br />
2 527 620<br />
3 316 430<br />
4 144 210<br />
5 54 60<br />
6 15 30<br />
7 3 10<br />
8 0 0<br />
EJEMPLO 3: TABLA DE VIDA VERTICAL PARA UN MAMÍFERO CON GENERACIONES<br />
SUPERPUESTAS, EL THAR DEL HIMALAYA Hemitragus jemlahicus (Caughley 1966)<br />
El thar es un ungulado similar a una cabra que vive aproximadamente 17 años y se reproduce<br />
anualmente. Fue introducido en Nueva Zelandia en 1909 y <strong>de</strong>s<strong>de</strong> entonces se distribuyó por todo el<br />
país. Deambula por zonas montañosas en tres tipos <strong>de</strong> grupos sociales: (1) hembras con cabritos, (2)<br />
machos juveniles, y (3) machos seniles. Su edad se <strong>de</strong>termina por el número <strong>de</strong> anillos <strong>de</strong><br />
crecimiento en los cuernos. Para muestrear se eligió un valle a mitad <strong>de</strong> camino entre el punto <strong>de</strong><br />
liberación original y el extremo <strong>de</strong> la distribución. La estructura <strong>de</strong> eda<strong>de</strong>s obtenida se muestra en la<br />
siguiente tabla (ax ); <strong>de</strong>bido a la dificultad <strong>de</strong> cazar a los cabritos menores <strong>de</strong> un año éstos no fueron<br />
capturados. La serie ax fue ajustada por una regresión cuadrática <strong>de</strong> la forma log ax = a + bx + cx2 para producir una serie monotónicamente <strong>de</strong>creciente <strong>de</strong>l número <strong>de</strong> individuos a lo largo <strong>de</strong> las<br />
clases <strong>de</strong> edad. De esta forma se eliminan algunos efectos no <strong>de</strong>seados <strong>de</strong>bidos a la variabilidad<br />
muestral y a una mortalidad diferencial a lo largo <strong>de</strong> las diferentes estaciones <strong>de</strong> reproducción.<br />
La función ajustada entre 1 y 12 años <strong>de</strong> edad es:<br />
log [ a' (x)] = 1.9673 +0.0246x - 0.01036 x 2
59<br />
Ecología General<br />
La tabla <strong>de</strong> vida fue construida para una cohorte hipotética <strong>de</strong> hembras ya que los animales no<br />
nacieron <strong>de</strong>ntro <strong>de</strong> un intervalo <strong>de</strong> tiempo corto.<br />
Edad (x) a(x) l(x) log10(l(x).1000) d(x) q(x) p(x) m(x)<br />
0 205 0<br />
1 96 0.005<br />
2 94 0.135<br />
3 89 0.440<br />
4 79 0.420<br />
5 68 0.465<br />
6 55 0.425<br />
7 43 0.460<br />
8 32 0.485<br />
9 22 0.500<br />
10 15 0.500<br />
11 10 0.470<br />
12 6 0.470<br />
>12 11 -<br />
REFERENCIAS<br />
Caughley G (1966) Mortality patterns in mammals. Ecology 47:906-918.<br />
Richards OW y Waloff N (1954) Studies on the biology and population dynamics of British grasshoppers.<br />
Anti-Locust Bull. 17:1-182.<br />
BIBLIOGRAFIA GENERAL<br />
Caughley G (1977) Analysis of vertebrate populations. Wiley, New York.<br />
Seber GAF (1982) The estimation of animal abundance. Griffin, London.<br />
Begon, Harper y Townsend (op. cit.).
60<br />
Ecología General<br />
Parte 3<br />
Aplicaciones <strong>de</strong> Tablas <strong>de</strong> vida.<br />
Interpretación <strong>de</strong> la dinámica poblacional <strong>de</strong> la laucha <strong>de</strong> campo,<br />
Calomys laucha, mediante la integración <strong>de</strong> los patrones <strong>de</strong><br />
supervivencia y reproducción <strong>de</strong> sus distintas cohortes.<br />
OBJETIVOS<br />
por David Bilenca<br />
• I<strong>de</strong>ntificar, mediante la aplicación <strong>de</strong> tablas <strong>de</strong> vida, los patrones <strong>de</strong> supervivencia y reproducción<br />
<strong>de</strong> las distintas cohortes que integran una población formada por generaciones superpuestas en<br />
un ambiente estacional.<br />
• Integrar la dinámica poblacional <strong>de</strong> una especie a partir <strong>de</strong> la dinámica <strong>de</strong> sus respectivas<br />
cohortes.<br />
MATERIALES<br />
Información <strong>de</strong>mográfica <strong>de</strong> campo (Kravetz et al. 1981, parcialmente modificada a los efectos<br />
didácticos) correspondientes a la laucha <strong>de</strong> campo Calomys laucha, roedor característico <strong>de</strong> los<br />
campos <strong>de</strong> cultivo <strong>de</strong> la región pampeana. Esta especie presenta habitualmente una estación<br />
reproductiva que se prolonga por 9-10 meses, tras lo cual comienza un breve receso reproductivo<br />
hasta la estación reproductiva siguiente. Bajo condiciones naturales, la longevidad <strong>de</strong> los individuos<br />
raramente exce<strong>de</strong> los 8 meses.<br />
Los datos proporcionados en la Tabla 1 correspon<strong>de</strong>n a los valores <strong>de</strong>: 1) abundancia, Nx<br />
(estimados a partir <strong>de</strong>l éxito <strong>de</strong> captura: No. <strong>de</strong> capturas / esfuerzo <strong>de</strong> captura), y 2) fecundidad<br />
específica por edad, mx, <strong>de</strong> tres cohortes características:<br />
-cohorte primaveral (nacidos en noviembre)<br />
-cohorte estival (nacidos en febrero)<br />
-cohorte otoñal (nacidos en mayo)<br />
Asimismo, se suministran los valores <strong>de</strong> Nx y mx obtenidos en los muestreos realizados en esos<br />
mismos meses.<br />
Finalmente, se proporcionan para cada cohorte los valores <strong>de</strong> Ro, Tc y r, así como los respectivos<br />
gráficos <strong>de</strong> log(lx*1000), ex y Vx en función <strong>de</strong> la edad.
9<br />
8<br />
7<br />
6<br />
5<br />
4<br />
3<br />
2<br />
1<br />
0<br />
Mes<br />
Edad (x)<br />
0.0016<br />
0<br />
0.0018<br />
3<br />
0<br />
0<br />
0<br />
0<br />
0<br />
0<br />
0.0078<br />
0<br />
0.094<br />
0<br />
0.02<br />
0<br />
N<br />
0.018<br />
0.61<br />
0.0168<br />
2.5<br />
0.0018<br />
0<br />
0.0018<br />
0<br />
0.015<br />
2.85<br />
0.0053<br />
0<br />
0.0176<br />
0<br />
0.034<br />
0.012<br />
3<br />
0.0266<br />
0.5<br />
D E<br />
0<br />
F M<br />
0.009<br />
1.9<br />
0.0266<br />
3.2<br />
0.0015<br />
0<br />
0.0038<br />
0.92<br />
0.0103<br />
1.15<br />
0.0174<br />
0.78<br />
0.0181<br />
2.9<br />
0.0929<br />
2.33<br />
0.1003<br />
0<br />
0.1581<br />
0.0028<br />
0<br />
0.009<br />
0<br />
0.0973<br />
0<br />
61<br />
0.0012<br />
0<br />
0.005<br />
0<br />
0.0366<br />
0<br />
0.0005<br />
0<br />
0.0017<br />
0<br />
0.0032<br />
0<br />
0.0009<br />
0.58<br />
0.0027<br />
0<br />
0.0004<br />
0.78<br />
0.0022<br />
3<br />
Ecología General<br />
0.0018<br />
3<br />
0.0013<br />
3<br />
0.0008<br />
A<br />
0<br />
M J J A S O N D E F<br />
Tabla 1. Estructura etaria <strong>de</strong> Calomys laucha. En cada intersección se presenta información <strong>de</strong> abundancia<br />
(Nx: bor<strong>de</strong> superior) y fecundidad (mx: bor<strong>de</strong> inferior)<br />
Parámetro<br />
COHORTE<br />
Noviembre<br />
COHORTE<br />
Febrero<br />
COHORTE<br />
Mayo<br />
Ro 7.616 4.463 0.123<br />
Tc 3.110 2.288 6.292<br />
R 0.653 0.654 -0.333<br />
Tabla 2. Valores <strong>de</strong>: 1) tasa <strong>de</strong> reproducción básica (Ro), 2) tiempo generacional (Tc ), y 3) tasa<br />
instantánea <strong>de</strong> crecimiento poblacional (r) estimados para las cohortes <strong>de</strong> Calomys laucha nacidas<br />
en noviembre, febrero y mayo.<br />
3<br />
0.0004<br />
3
Figuras: log(lx*1000), ex y Vx en función <strong>de</strong> la edad.<br />
Log 10 (l(x)*1000)<br />
V(x)<br />
3,000<br />
2,500<br />
2,000<br />
1,500<br />
1,000<br />
0,500<br />
9,000<br />
8,000<br />
7,000<br />
6,000<br />
5,000<br />
4,000<br />
3,000<br />
2,000<br />
1,000<br />
0,000<br />
Curvas <strong>de</strong> supervivencia: log (l(x)*1000)<br />
0,000<br />
0 1 2 3 4 5 6 7 8 9<br />
edad (meses)<br />
Valor reproductivo: V(x)<br />
0 1 2 3 4 5 6 7 8 9<br />
edad (meses)<br />
62<br />
log l(x) 1000 N<br />
log l(x) 1000 F<br />
log l(x) 1000 M<br />
V(x)NOV<br />
V(x)FEB<br />
V(x)MAY<br />
Ecología General
DESARROLLO<br />
Sobre la base <strong>de</strong> la información proporcionada, conteste las siguientes preguntas:<br />
63<br />
Ecología General<br />
1) Indique el momento <strong>de</strong> inicio y <strong>de</strong> finalización <strong>de</strong> la estación reproductiva <strong>de</strong> Calomys laucha.<br />
2) ¿Existe alguna cohorte que se reproduzca más tardíamente? ¿Cuál? ¿A qué lo atribuye?<br />
3) I<strong>de</strong>ntifique la(s) cohorte(s) que realizan el mayor aporte reproductivo.<br />
4) ¿A qué cohortes pertenecen los individuos que inician la nueva estación reproductiva?<br />
5) ¿A qué factores atribuye la forma <strong>de</strong> las curvas <strong>de</strong> log (lx *1000), ex y Vx que <strong>de</strong>scribe cada<br />
cohorte?<br />
6) ¿Qué limitaciones consi<strong>de</strong>ra Usted que encontraría si tratara <strong>de</strong> compren<strong>de</strong>r la dinámica<br />
poblacional <strong>de</strong> Calomys laucha contando exclusivamente con los datos <strong>de</strong> una <strong>de</strong> sus cohortes?<br />
¿Qué papel funcional le asignaría a cada cohorte?<br />
7) ¿Consi<strong>de</strong>ra posible construir una tabla <strong>de</strong> vida e interpretar la dinámica poblacional <strong>de</strong> Calomys<br />
laucha contando exclusivamente con los datos provenientes <strong>de</strong> un único muestreo (noviembre,<br />
febrero o mayo)? En caso <strong>de</strong> que su respuesta sea afirmativa, indique cuál <strong>de</strong> los muestreos utilizaría<br />
y justifique su respuesta.<br />
REFERENCIAS<br />
Kravetz FO, Manjón MC, Busch M, Percich RE, Marconi PN y Torres MP (1981) Ecología <strong>de</strong> Calomys<br />
laucha (Ro<strong>de</strong>ntia, Cricetidae) en el <strong>de</strong>partamento <strong>de</strong> Río Cuarto (Córdoba). I. Dinámica <strong>de</strong><br />
población. Ecología 6:15-22.<br />
BIBLIOGRAFIA GENERAL<br />
Caughley G (1977) Analysis of vertebrate populations. Wiley, New York.<br />
Tinkle DW (1969) The concept of reproductive effort and its relation to the evolution of life histories of<br />
lizards. Am. Nat. 103:501-516.<br />
Wittenberger JF (1979) A mo<strong>de</strong>l for <strong>de</strong>layed reproduction in iteroparous animals. Am. Nat. 114:439-<br />
446.<br />
Zammuto RM (1987) Life histories of mammals: analyses among and within Spermophilus columbianus life<br />
tables. Ecology 68:1351-1363.<br />
AGRADECIMIENTOS<br />
El presente TP ha sido rediseñado a partir <strong>de</strong> un TP realizado en el año 1989 por integrantes <strong>de</strong> Ecología<br />
General. A Ricardo Gürtler, Mónica Castañera y Nancy López por su gentileza para revisar y terminar <strong>de</strong><br />
compaginar el TP.
Trabajo práctico 6<br />
MODELOS DE CRECIMIENTO POBLACIONAL<br />
(1) INTRODUCCIÓN<br />
64<br />
Ecología General<br />
A fin <strong>de</strong> po<strong>de</strong>r <strong>de</strong>scribir los cambios en los números poblacionales a través <strong>de</strong>l tiempo se han<br />
<strong>de</strong>sarrollado distintos mo<strong>de</strong>los matemáticos.<br />
Los objetivos <strong>de</strong> estos mo<strong>de</strong>los pue<strong>de</strong>n ser:<br />
a) Describir el comportamiento <strong>de</strong> una serie <strong>de</strong> valores <strong>de</strong> <strong>de</strong>nsidad poblacional en el tiempo a fin <strong>de</strong><br />
pre<strong>de</strong>cir el comportamiento futuro <strong>de</strong> la población. Este caso respon<strong>de</strong> a un mo<strong>de</strong>lo simplemente<br />
<strong>de</strong>scriptivo y no requiere <strong>de</strong>l planteo <strong>de</strong> hipótesis sobre las causas <strong>de</strong> variación <strong>de</strong> los números<br />
poblacionales.<br />
b) Consi<strong>de</strong>rar características conocidas <strong>de</strong> la población, incorporando variables que se supone que<br />
influyen sobre los cambios <strong>de</strong> la abundancia. La corrida <strong>de</strong>l mo<strong>de</strong>lo en una serie <strong>de</strong> tiempo (cuyas<br />
unida<strong>de</strong>s pue<strong>de</strong>n variar según el organismo, como horas para bacterias o años para otros grupos),<br />
comparada con los datos reales, permite evaluar si las variables incorporadas eran relevantes y si<br />
servían para <strong>de</strong>scribir el crecimiento poblacional.<br />
Los cambios que experimenta una población en el tiempo <strong>de</strong>pen<strong>de</strong>n <strong>de</strong> las tasas <strong>de</strong> natalidad,<br />
mortalidad, inmigración y emigración.<br />
b= Tasa <strong>de</strong> natalidad= número <strong>de</strong> individuos nacidos/(intervalo <strong>de</strong> tiempo*número <strong>de</strong> individuos <strong>de</strong> la<br />
población)<br />
d= Tasa <strong>de</strong> mortalidad= número <strong>de</strong> individuos muertos/(intervalo <strong>de</strong> tiempo*número <strong>de</strong> individuos <strong>de</strong><br />
la población)<br />
i= Tasa <strong>de</strong> inmigración= número <strong>de</strong> individuos ingresados a la población/(intervalo <strong>de</strong> tiempo*número<br />
<strong>de</strong> individuos <strong>de</strong> la población)<br />
e= Tasa <strong>de</strong> emigración= número <strong>de</strong> individuos que se van <strong>de</strong> la población/(intervalo <strong>de</strong><br />
tiempo*número <strong>de</strong> individuos <strong>de</strong> la población)<br />
La tasa intrínseca <strong>de</strong> crecimiento per cápita sintetiza el balance entre los factores <strong>de</strong> crecimiento<br />
(natalidad e inmigración) y <strong>de</strong>crecimiento (mortalidad y emigración) poblacional = b+i-d-e, y<br />
representa por cuánto se multiplicará la población por unidad <strong>de</strong> tiempo.<br />
Hay dos tipos principales <strong>de</strong> mo<strong>de</strong>los, los que asumen un crecimiento <strong>de</strong>nso in<strong>de</strong>pendiente,<br />
exponencial o geométrico, y los que asumen que la velocidad <strong>de</strong> crecimiento <strong>de</strong>pen<strong>de</strong> <strong>de</strong> la <strong>de</strong>nsidad,<br />
y la población crece en forma logística. Los que vamos a ver en este trabajo práctico tienen dos<br />
supuestos compartidos:<br />
1- Todos los individuos contribuyen <strong>de</strong> la misma manera al crecimiento poblacional.<br />
2- No hay variaciones en la estructura <strong>de</strong> eda<strong>de</strong>s a lo largo <strong>de</strong>l tiempo.<br />
3- La emigración y la inmigración se compensan mutuamente<br />
EL MODELO EXPONENCIAL<br />
En el mo<strong>de</strong>lo exponencial o geométrico se asume que la tasa <strong>de</strong> crecimiento es in<strong>de</strong>pendiente <strong>de</strong> la<br />
<strong>de</strong>nsidad, es <strong>de</strong>cir, que se mantiene constante para todo el rango <strong>de</strong> <strong>de</strong>nsida<strong>de</strong>s (es <strong>de</strong>cir, b+i-d-e=<br />
constante). La cantidad neta <strong>de</strong> individuos que se agregan a la población, o tasa <strong>de</strong> reclutamiento<br />
neto, aumenta linealmente con la <strong>de</strong>nsidad (porque resulta <strong>de</strong> la multiplicación <strong>de</strong> la tasa <strong>de</strong><br />
crecimiento per cápita, que es constante, por N que aumenta). La población crece exponencialmente.
65<br />
Ecología General<br />
Este mo<strong>de</strong>lo también asume que la disponibilidad <strong>de</strong> recursos <strong>de</strong>l ambiente es ilimitada, <strong>de</strong> manera<br />
que el aumento en el número <strong>de</strong> individuos <strong>de</strong> la población no produce una disminución en la<br />
cantidad <strong>de</strong> recursos disponible para cada uno.<br />
Crecimiento exponencial discreto<br />
El cambio <strong>de</strong> la población en el tiempo pue<strong>de</strong> ser <strong>de</strong>scripto por:<br />
N(t+1) = N(t)*R = N(t-1).R*R = N(t-1)*R 2<br />
don<strong>de</strong> R es la tasa <strong>de</strong> crecimiento per cápita por unidad <strong>de</strong> tiempo discreto<br />
lo que da lugar a la fórmula general:<br />
N(t) = N(0) *R t don<strong>de</strong> N(0) es el valor <strong>de</strong> <strong>de</strong>nsidad inicial. EC 1<br />
Esta es una forma <strong>de</strong> expresar el crecimiento consi<strong>de</strong>rando unida<strong>de</strong>s discretas <strong>de</strong> tiempo, es <strong>de</strong>cir<br />
que el crecimiento se produce <strong>de</strong> a saltos, sin pasos intermedios. Por ejemplo, en una población <strong>de</strong><br />
insectos cuyos adultos se mueren luego <strong>de</strong> reproducirse, el salto se produce cuando hay<br />
reproducción, luego la población se mantiene relativamente estable (aunque pue<strong>de</strong> haber muertes)<br />
hasta que los juveniles maduran y se reproducen, produciendo otro incremento.<br />
Si R=1, la población mantiene su tamaño constante, si R1, la<br />
población crece.<br />
Crecimiento exponencial continuo.<br />
La forma <strong>de</strong> expresar el crecimiento en forma continua pue<strong>de</strong> <strong>de</strong>rivarse consi<strong>de</strong>rando un intervalo <strong>de</strong><br />
tiempo infinitamente chico, entonces la velocidad <strong>de</strong> crecimiento pue<strong>de</strong> ser <strong>de</strong>scripta mediante la<br />
ecuación diferencial, dN/dt. Esta velocidad <strong>de</strong> crecimiento <strong>de</strong>pen<strong>de</strong> <strong>de</strong> la diferencia entre las tasas <strong>de</strong><br />
natalidad (b) y mortalidad (d).<br />
dN/dt = (b-d)*N. La diferencia (b-d) es constante y se <strong>de</strong>fine como la tasa instantánea <strong>de</strong> crecimiento<br />
poblacional, r.<br />
Para po<strong>de</strong>r pre<strong>de</strong>cir el tamaño poblacional a lo largo <strong>de</strong>l tiempo se <strong>de</strong>be integrar la ecuación anterior:<br />
N(t) = N(t- Δ t) * e r* Δt don<strong>de</strong> r es la tasa <strong>de</strong> crecimiento per cápita consi<strong>de</strong>rando el tiempo<br />
continuo<br />
O, como se expresa habitualmente,<br />
N(t)= N(0)* e r* t , don<strong>de</strong> N(0) es el valor inicial <strong>de</strong>l que parte la población EC 2<br />
Esta es la solución <strong>de</strong> la ecuación diferencial para el crecimiento exponencial <strong>de</strong> la tasa <strong>de</strong><br />
reclutamiento neto, dN/dt =r*N<br />
En el mo<strong>de</strong>lo exponencial continuo, si r0 la población crece.<br />
Algunas <strong>de</strong> las relaciones que se <strong>de</strong>rivan <strong>de</strong> este mo<strong>de</strong>lo se muestran en la Figura 1.
dN/dt*N<br />
N<br />
66<br />
Ecología General<br />
Figura 1. Variación <strong>de</strong> la tasa <strong>de</strong> crecimiento per cápita (r = dN/(dt*N)) y la tasa <strong>de</strong> reclutamiento neto<br />
(dN/dt) en función <strong>de</strong> N, y <strong>de</strong> N en función <strong>de</strong> t para el mo<strong>de</strong>lo exponencial continuo.<br />
LOS LÍMITES AL CRECIMIENTO<br />
N<br />
dN/dt<br />
t<br />
Hace aproximadamente 200 años, Thomas Malthus argumentó que las poblaciones no pue<strong>de</strong>n crecer<br />
in<strong>de</strong>finidamente sino que son limitadas por escasez <strong>de</strong> alimento, espacio u otros recursos esenciales.<br />
40 años <strong>de</strong>spués, el matemático Verhulst dio una expresión matemática para esta i<strong>de</strong>a, que fue<br />
llamada la ecuación logística. En forma in<strong>de</strong>pendiente, Pearl y Reed <strong>de</strong>sarrollaron el mismo mo<strong>de</strong>lo<br />
para <strong>de</strong>scribir el crecimiento <strong>de</strong> la población <strong>de</strong> Estados Unidos <strong>de</strong> América entre los años 1790 y<br />
1910.<br />
La i<strong>de</strong>a <strong>de</strong> que existe un límite al crecimiento poblacional pue<strong>de</strong> ser formalizada como sigue:<br />
Consi<strong>de</strong>remos una población <strong>de</strong> N individuos en las que cada organismo necesita l unida<strong>de</strong>s <strong>de</strong><br />
recursos para sobrevivir hasta dar un hijo (y no más que uno). l será la <strong>de</strong>manda <strong>de</strong> subsistencia<br />
individual. Si todos los miembros <strong>de</strong> la población reciben continuamente su ración <strong>de</strong> subsistencia, o<br />
sea que la cantidad total <strong>de</strong> alimento disponible para la población es L= l *N, la población no<br />
aumentará ni disminuirá <strong>de</strong> número, sino que permanecerá en estado <strong>de</strong> equilibrio. Este valor <strong>de</strong><br />
<strong>de</strong>nsidad en el cual la población se mantiene en equilibrio se <strong>de</strong>nomina capacidad <strong>de</strong> carga <strong>de</strong>l<br />
ambiente. Si la cantidad total <strong>de</strong> alimento disponible para la población es L> l *N, habrá un<br />
suplemento <strong>de</strong> recursos disponible para reproducción adicional y la población crecerá. Si, por el<br />
contrario, L< l *N, la población disminuirá <strong>de</strong> tamaño. En un ambiente dado, L pue<strong>de</strong> ser un valor más<br />
o menos constante (por renovación <strong>de</strong> los recursos), o pue<strong>de</strong> variar estacionalmente o por otros<br />
factores. Si L se mantiene constante a medida que la <strong>de</strong>nsidad poblacional aumenta, como la<br />
<strong>de</strong>manda (l * N) se va haciendo cada vez mayor, llega un momento en que iguala a L, en ese<br />
momento, que correspon<strong>de</strong> a cuando N= K, el crecimiento se <strong>de</strong>tiene. En otras palabras, la<br />
<strong>de</strong>nsidad <strong>de</strong> equilibrio, K, es igual a la relación entre la disponibilidad total <strong>de</strong> recursos y la <strong>de</strong>manda<br />
individual. Este parámetro refleja la <strong>de</strong>nsidad <strong>de</strong> individuos que pue<strong>de</strong> mantenerse in<strong>de</strong>finidamente<br />
con un suministro constante <strong>de</strong> recursos.<br />
K= L/ l<br />
N
67<br />
Ecología General<br />
Como la cantidad <strong>de</strong> <strong>de</strong>scen<strong>de</strong>ncia que produce cada individuo <strong>de</strong>pen<strong>de</strong> <strong>de</strong> la relación entre<br />
suministro y <strong>de</strong>manda (L/l), cuanto mayor sea esta relación, más rápido será el crecimiento. Pero a<br />
medida que aumenta la población, la <strong>de</strong>manda aumenta, con lo que el crecimiento será más lento.<br />
Esto conduce a que haya una relación inversa entre el tamaño poblacional y la tasa intrínseca <strong>de</strong><br />
crecimiento per cápita.<br />
Teniendo en cuenta que la tasa <strong>de</strong> crecimiento per cápita disminuye cuando aumenta la <strong>de</strong>nsidad<br />
poblacional, la ecuación que <strong>de</strong>scribe los cambios en los números poblacionales a través <strong>de</strong>l tiempo<br />
se convierte en:<br />
N t+1= Nt + (R-1) Nt (1-Nt/K) Ecuación logística discreta EC 3<br />
La forma diferencial, que asume crecimiento continuo, se expresa como:<br />
Nt= K*N0/(N0+(K- N0)*e (-r*t ) ) Ecuación logística continua EC 4<br />
y el reclutamiento neto dN/dt= r*N* [1-Nt/K],<br />
La ecuación logística (tanto discreta como contínua) es una ecuación fundamental en la dinámica<br />
poblacional, conteniendo los elementos básicos <strong>de</strong> feedback positivo (el crecimiento al principio se<br />
acelera con N) y negativo (la tasa <strong>de</strong> crecimiento per cápita disminuye con N). Es un mo<strong>de</strong>lo simple,<br />
que asume:<br />
1- La tasa <strong>de</strong> crecimiento per cápita está linealmente relacionada con la <strong>de</strong>nsidad, o sea que la<br />
intensidad <strong>de</strong>l feedback negativo es el mismo a todas las <strong>de</strong>nsida<strong>de</strong>s.<br />
2- El feedback negativo sobre la tasa <strong>de</strong> crecimiento ocurre inmediatamente, sin tiempo <strong>de</strong> retardo<br />
(en el mo<strong>de</strong>lo discreto <strong>de</strong> una generación a la siguiente).<br />
3- No hay efectos cooperativos, en los que un aumento en <strong>de</strong>nsidad se traduciría en mayor<br />
reproducción o supervivencia.<br />
4- El sistema tiene un sólo punto <strong>de</strong> equilibrio, K.<br />
Para el caso <strong>de</strong>l mo<strong>de</strong>lo continuo se verifican las siguientes relaciones:<br />
dN/dt*N<br />
rmax<br />
0<br />
N<br />
Pendiente<br />
= rmax/K<br />
dN/dt<br />
K K/2<br />
N<br />
Figura 2. Variación <strong>de</strong> la tasa <strong>de</strong> crecimiento per cápita (r = dN/(dt*N)) y la tasa <strong>de</strong> reclutamiento neto<br />
(dN/dt) en función <strong>de</strong> N, y <strong>de</strong> N en función <strong>de</strong> t para el mo<strong>de</strong>lo logístico continuo.<br />
K<br />
N<br />
K<br />
t
68<br />
Ecología General<br />
Estimación <strong>de</strong> los parámetros K y R<br />
Para el mo<strong>de</strong>lo logístico discreto estos parámetros se estiman a partir <strong>de</strong> la ecuación (Ec 3) y la<br />
relación lineal entre Nt/Nt+1 (es <strong>de</strong>cir 1/R) y Nt, como se muestra en la Figura 3.<br />
Figura 3: relación lineal <strong>de</strong> la variación <strong>de</strong> la inversa <strong>de</strong>l incremento poblacional (Nt/Nt+1) en función <strong>de</strong><br />
la <strong>de</strong>nsidad (Nt) para el mo<strong>de</strong>lo logístico discreto<br />
Si se pue<strong>de</strong> graficar esta relación para datos experimentales (u observacionales), se pue<strong>de</strong>n<br />
<strong>de</strong>terminar dos puntos fundamentales en la recta formada:<br />
A) es el punto <strong>de</strong> la interceptación <strong>de</strong> la recta con el eje y, y <strong>de</strong>termina el valor <strong>de</strong> 1/Rmax.<br />
B) es el punto don<strong>de</strong> no hay variación en el tamaño poblacional (Nt/Nt+1=1), que es por <strong>de</strong>finición<br />
el que indica la capacidad <strong>de</strong> carga <strong>de</strong>l sistema.<br />
Para una población con crecimiento continuo el rmax pue<strong>de</strong> ser <strong>de</strong>terminado resolviendo mediante<br />
métodos iterativos la fórmula:<br />
∞<br />
∑ e -rmax*x*lx*mx = 1<br />
X = 0<br />
Cuando la población presenta una estructura estable <strong>de</strong> eda<strong>de</strong>s, es posible obtener una estimación<br />
aproximada mediante la relación lnR=r<br />
Bibliografía.<br />
Krebs, Ch. J. 1978. Ecology: The experimental analysis of distribution and abundance. Harper & Row,<br />
Publishers. New York.<br />
Sharov, A. Quantitative Population Ecology. http: www.gypsymoth.ento.vt.edu/sharov
(2) ACTIVIDADES<br />
69<br />
Ecología General<br />
1. Estimación <strong>de</strong> los parámetros <strong>de</strong> la ecuación <strong>de</strong> crecimiento <strong>de</strong> Lemna sp. a partir <strong>de</strong> datos<br />
<strong>de</strong> laboratorio.<br />
Se trabajará con los datos <strong>de</strong> la variación en los números <strong>de</strong> Lemna sp. obtenidos en el trabajo<br />
experimental <strong>de</strong> Recursos y Condiciones <strong>de</strong>l primer cuatrimestre <strong>de</strong> <strong>2012.</strong><br />
Esta especie fue sometida a dos niveles <strong>de</strong> concentración <strong>de</strong> nutrientes (C1 y C2) bajo luz natural,<br />
introduciendo 10 individuos en recipientes conteniendo una <strong>de</strong> las soluciones. Se hicieron 3 réplicas<br />
para cada concentración y se contó el número <strong>de</strong> individuos a intervalos <strong>de</strong> 7 días.<br />
Consi<strong>de</strong>rando los valores promedio obtenidos en cada semana <strong>de</strong> medición (Tablas 1 y 2), para este<br />
trabajo práctico asumiremos que la cantidad total <strong>de</strong> nutrientes disponible se mantiene constante a lo<br />
largo <strong>de</strong>l tiempo (por lo que al aumentar la cantidad <strong>de</strong> individuos totales, la disponibilidad por<br />
individuo disminuye).<br />
Tabla 1: Variación en el número promedio <strong>de</strong> individuos <strong>de</strong> Lemna sp. a lo largo <strong>de</strong>l tiempo para la<br />
concentración 1 <strong>de</strong> nutrientes:<br />
Fecha 29/3 3/4 10/4 17/4 24/4 Unida<strong>de</strong>s<br />
t (días) 0<br />
N<br />
R= N(t+1)/Nt --<br />
1/R<br />
Tabla 2: Variación en el número promedio <strong>de</strong> individuos <strong>de</strong> Lemna sp. a lo largo <strong>de</strong>l tiempo para la<br />
concentración 2 <strong>de</strong> nutrientes<br />
Fecha 29/3 3/4 10/4 17/4 24/4 Unida<strong>de</strong>s<br />
t (días) 0<br />
N<br />
R= N(t+1)/Nt --<br />
1/R<br />
1A. Descripción <strong>de</strong>l crecimiento poblacional y cálculo <strong>de</strong> los parámetros <strong>de</strong> crecimiento.<br />
1. Completar los datos <strong>de</strong> las Tablas.<br />
2. Graficar las variaciones <strong>de</strong> los números <strong>de</strong> individuos <strong>de</strong> Lemna sp. a lo largo <strong>de</strong>l tiempo.<br />
Compare las curvas obtenidas con los mo<strong>de</strong>los exponencial y logístico. ¿a cuál se parecen<br />
los resultados para cada una <strong>de</strong> las concentraciones?<br />
3. Graficar la inversa <strong>de</strong> la tasa <strong>de</strong> incremento poblacional (Nt/Nt+1) en función <strong>de</strong> N. ¿Cuál es el<br />
N que correspon<strong>de</strong> consi<strong>de</strong>rar, Nt o Nt+1? ¿Qué significado biológico tiene la or<strong>de</strong>nada al<br />
origen? ¿Qué significado biológico tiene el valor <strong>de</strong> N cuando 1/R=1?<br />
4. Estimar los valores <strong>de</strong> Rmax y <strong>de</strong> K para ambas concentraciones. ¿A qué atribuye las<br />
diferencias observadas?¿En qué unida<strong>de</strong>s se expresan?<br />
5. ¿Es posible con estos datos obtener una estimación <strong>de</strong> r max para cada caso?<br />
1B: Simulación <strong>de</strong>l crecimiento poblacional hasta t= 100, a partir <strong>de</strong> los parámetros estimados en la<br />
parte anterior.<br />
1. A partir <strong>de</strong> los valores estimados <strong>de</strong> K y el r máximo, estime los valores <strong>de</strong> abundancia<br />
esperados <strong>de</strong> acuerdo a la ecuación logística continua partiendo <strong>de</strong> N = 10. (Use la EC 4<br />
<strong>de</strong> la parte introductoria).<br />
2. Consi<strong>de</strong>rando el valor <strong>de</strong> R máximo estimado, calcule los valores esperados <strong>de</strong><br />
abundancia <strong>de</strong> la población <strong>de</strong> acuerdo al mo<strong>de</strong>lo exponencial continuo. (Use la EC 2 <strong>de</strong> la<br />
parte introductoria).<br />
Suponga una especie hipotética que se reproduce por pulsos cada siete días, y para la cual se<br />
sabe que los parámetros R y K son similares a los obtenidos para Lemna sp. :
70<br />
Ecología General<br />
3. Estime los valores <strong>de</strong> abundancia esperados <strong>de</strong> acuerdo a la ecuación logística discreta<br />
partiendo <strong>de</strong> N = 10. (Use la EC 3 <strong>de</strong> la parte introductoria).<br />
4. Calcule los valores esperados <strong>de</strong> abundancia <strong>de</strong> la población <strong>de</strong> acuerdo al mo<strong>de</strong>lo<br />
exponencial discreto. (Use la EC 1 <strong>de</strong> la parte introductoria)<br />
2. Ejercicios adicionales<br />
2a) Teniendo en cuenta las variaciones en el número promedio <strong>de</strong> individuos <strong>de</strong> Lemna sp.<br />
observadas durante la realización <strong>de</strong>l trabajo práctico <strong>de</strong>l primer cuatrimestre <strong>de</strong> 2008.<br />
Fecha Concentración 1 Concentración 2<br />
27/3 10 10<br />
3/4 22,33 26,66<br />
10/4 40,66 63,66<br />
17/4 49,33 96,33<br />
24/4 52 109,33<br />
• Calcular Rmax y K para estos valores<br />
• Comparar los resultados obtenidos con los <strong>de</strong>l experimento <strong>de</strong> este año<br />
• Discutir posibles explicaciones que permitan explicar las diferencias entre experimentos en caso<br />
<strong>de</strong> que las hubiera<br />
2b) Calcule los tiempos <strong>de</strong> duplicación para las siguientes poblaciones humanas, teniendo en cuenta<br />
que si dN / dt = r N, entonces Nt / N0 = e r t .<br />
País r * t <strong>de</strong> duplicación<br />
Argelia 0.033<br />
Argentina 0.015<br />
Irlanda 0.004<br />
Alemania - 0.002<br />
* extraído <strong>de</strong> Ecología - Krebs CJ 1985<br />
• ¿Qué supuestos se <strong>de</strong>ben hacer para pre<strong>de</strong>cir estos tiempos <strong>de</strong> duplicación?<br />
• Des<strong>de</strong> un punto <strong>de</strong> vista biológico, ¿cómo explicaría que se hayan obtenido distintos valores <strong>de</strong> r<br />
para una misma especie?<br />
• Suponga que una institución <strong>de</strong> asistencia internacional, <strong>de</strong>dicada a fomentar la educación,<br />
necesita conocer dón<strong>de</strong> invertir en la construcción <strong>de</strong> nuevas escuelas. ¿Pue<strong>de</strong> ayudarla a<br />
<strong>de</strong>cidir?<br />
2c) La bacteria Escherichia coli se divi<strong>de</strong> aproximadamente cada 20 minutos bajo condiciones<br />
óptimas.<br />
• Comenzando con una bacteria, ¿cuántas bacterias habrá al cabo <strong>de</strong> 48 horas? nota: N0 = 1, <strong>de</strong>be<br />
calcular el N(t = 48) . La tasa finita <strong>de</strong> crecimiento (R) es 2. Recuer<strong>de</strong> que: Nt = R t N0<br />
• Si el diámetro <strong>de</strong> un individuo <strong>de</strong> E. coli es <strong>de</strong> aproximadamente 10 - 6 m, ¿qué volumen ocupará la<br />
población <strong>de</strong> bacterias estimada en (a)? Recuer<strong>de</strong> que la forma <strong>de</strong> este tipo <strong>de</strong> células pu<strong>de</strong><br />
consi<strong>de</strong>rarse como aproximadamente esférica. El volumen <strong>de</strong> una esfera se calcula como: (4 / 3).<br />
π . R 3 don<strong>de</strong> R es el radio <strong>de</strong> la esfera.<br />
• Compare el valor obtenido en el punto anterio con el volumen <strong>de</strong>l planeta tierra (1.087 . 10 21 m 3 ).<br />
En función <strong>de</strong> lo anterior, ¿pue<strong>de</strong> pensarse un proceso <strong>de</strong> crecimiento <strong>de</strong> tipo exponencial por<br />
largos periodos <strong>de</strong> duración?<br />
Trabajo práctico 7<br />
COMPETENCIA
71<br />
Ecología General<br />
PARTE 1: Bases matemáticas <strong>de</strong>l mo<strong>de</strong>lo <strong>de</strong> Competencia <strong>de</strong> Lotka-<br />
Volterra.<br />
INTRODUCCION<br />
Los mo<strong>de</strong>los <strong>de</strong> crecimiento <strong>de</strong>nso<strong>de</strong>pendiente como la ecuación logística asumen la existencia <strong>de</strong><br />
competencia intraespecífica; los recursos se transforman en limitantes a medida que la población se<br />
incrementa, y la tasa <strong>de</strong> crecimiento per cápita <strong>de</strong>clina. Para representar los efectos interespecíficos<br />
<strong>de</strong>nso<strong>de</strong>pendientes, un término adicional es incorporado a la ecuación logística, y el par <strong>de</strong><br />
expresiones resultantes compren<strong>de</strong>n “las ecuaciones <strong>de</strong> competencia <strong>de</strong> Lotka-Volterra”, las cuales<br />
proveen un vehículo simple e históricamente importante para pensar acerca <strong>de</strong> las interacciones<br />
competitivas.<br />
En las ecuaciones <strong>de</strong> Lotka-Volterra, las <strong>de</strong>nsida<strong>de</strong>s <strong>de</strong> ambas especies son sustraídas<br />
<strong>de</strong>s<strong>de</strong> la capacidad <strong>de</strong> carga para dar un término <strong>de</strong> retroalimentación <strong>de</strong>nso<strong>de</strong>pendiente, y el<br />
número <strong>de</strong> competidores interespecíficos es pesado por el término <strong>de</strong>nominado coeficiente <strong>de</strong><br />
competencia el cual varía con la similitud en requerimientos <strong>de</strong> recursos <strong>de</strong> las especies. Así:<br />
dN1 K1 - N1 - α N2<br />
= r1<br />
N1 dt K1<br />
dN2 K2 - N2 - β N1<br />
= r2<br />
N2 dt K2<br />
Don<strong>de</strong> N1 representa la <strong>de</strong>nsidad <strong>de</strong> la especie 1, K1 es la capacidad <strong>de</strong> carga <strong>de</strong> la especie 1, r1 es<br />
su tasa intrínseca <strong>de</strong> incremento, y α es el coeficiente <strong>de</strong> competencia, una constante <strong>de</strong><br />
proporcionalidad que <strong>de</strong>fine a cuántos individuos <strong>de</strong> la especie 1 equivale un individuo <strong>de</strong> la especie<br />
2. En la segunda expresión, β es el coeficiente análogo que pesa el efecto <strong>de</strong> cada individuo <strong>de</strong> la<br />
especie 1 sobre la especie 2.<br />
Aunque no tenemos una solución <strong>de</strong> forma cerrada para estas ecuaciones, po<strong>de</strong>mos sacar<br />
interesantes observaciones acerca <strong>de</strong> su dinámica cerca <strong>de</strong>l equilibrio cuando dN1 / dt = dN2 / dt = 0.<br />
Hay un equilibrio trivial cuando r ó N son igual a 0; un caso más interesante suce<strong>de</strong> cuando:<br />
N1 = K1 - α N2 y N2 = K2 - β N1<br />
Estas ecuaciones representan líneas rectas en el espacio <strong>de</strong>finido por las coor<strong>de</strong>nadas <strong>de</strong> N2 vs N1,<br />
son las isoclinas <strong>de</strong> crecimiento neto cero <strong>de</strong> cada una <strong>de</strong> las especies, y representan los pares <strong>de</strong><br />
<strong>de</strong>nsida<strong>de</strong>s <strong>de</strong> las dos especies para las cuales el crecimiento es cero (en la ecuación <strong>de</strong> 1, 1 está en<br />
equilibrio, según la ecuación <strong>de</strong> 2, 2 está en equilibrio). Si las isoclinas se cruzan, el punto <strong>de</strong><br />
intersección representa el par <strong>de</strong> <strong>de</strong>nsida<strong>de</strong>s (N1, N2) en las cuales ambas especies están en<br />
equilibrio.<br />
PARTE 2: Competencia. Trabajo experimental.<br />
INTRODUCCION<br />
La competencia interespecífica ha sido <strong>de</strong>finida como la interacción entre individuos <strong>de</strong> dos o más<br />
especies que causa un efecto adverso sobre su crecimiento, supervivencia, ajuste adaptativo (fitness)<br />
o tamaño poblacional (Giller, 1984). Se genera competencia cuando dos o más especies se<br />
superponen en el uso <strong>de</strong> recursos limitantes, produciéndose a través <strong>de</strong> un daño directo<br />
(interferencia), o por la disminución <strong>de</strong> recursos disponibles (explotación).<br />
Las consecuencias <strong>de</strong> la competencia interespecífica pue<strong>de</strong>n manifestarse <strong>de</strong> distintas maneras: en<br />
relaciones numéricas inversas o en relaciones espaciales inversas (distintas preferencias <strong>de</strong> hábitat,<br />
territorialidad interespecífica). A su vez los cambios numéricos <strong>de</strong>bidos a la competencia pue<strong>de</strong>n
72<br />
Ecología General<br />
darse a través <strong>de</strong> distintos mecanismos: variaciones en la mortalidad, natalidad, inmigraciones o<br />
emigraciones.<br />
El constatar la existencia <strong>de</strong> relaciones numéricas o espaciales inversas entre dos especies no<br />
implica necesariamente que estas sean competidoras, pues podría haber otros factores que estén<br />
<strong>de</strong>terminando estas variaciones, como por ejemplo, distintas preferencias <strong>de</strong> hábitat. Sin embargo,<br />
este tipo <strong>de</strong> relaciones representa una evi<strong>de</strong>ncia observacional <strong>de</strong> la competencia, que pue<strong>de</strong> ser<br />
puesta a prueba mediante experiencias <strong>de</strong> remoción.<br />
Existen distintos métodos <strong>de</strong> evaluar la competencia entre especies, muchos <strong>de</strong> ellos <strong>de</strong>rivados <strong>de</strong><br />
los mo<strong>de</strong>los <strong>de</strong> competencia <strong>de</strong> Lotka-Volterra. Este mo<strong>de</strong>lo tiene la limitante <strong>de</strong> que propone<br />
soluciones para sistemas en equilibrio y asume coeficientes <strong>de</strong> competencia constantes<br />
(in<strong>de</strong>pendientes <strong>de</strong> la <strong>de</strong>nsidad) y, por lo tanto, isoclinas rectas.<br />
De acuerdo a este mo<strong>de</strong>lo, cada especie interactuante va a llegar, en el equilibrio, a una <strong>de</strong>nsidad<br />
que correspon<strong>de</strong> a:<br />
N1 (eq)= K1 - α* N2 (eq)<br />
N2 (eq)= K2 - β*N1 (eq)<br />
Don<strong>de</strong>: α es el coeficiente <strong>de</strong> competencia <strong>de</strong> la especie 2 sobre la especie 1 y β es el coeficiente <strong>de</strong><br />
competencia <strong>de</strong> la especie 1 sobre la especie 2.<br />
(Nota: es posible encontrar en las diferentes bibliografías a los coeficientes <strong>de</strong> competencia como:<br />
α = α12; β = α21).<br />
En estas relaciones se basan los métodos <strong>de</strong> regresión para estimar los coeficientes <strong>de</strong> competencia.<br />
Si tenemos datos <strong>de</strong> censos correspondientes a "distintos sitios equivalentes", las relaciones<br />
numéricas entre las dos especies reflejarán las relaciones <strong>de</strong> competencia. Una regresión no<br />
significativa entre sus números significará ausencia <strong>de</strong> interacción, una positiva, interacción positiva, y<br />
una negativa, competencia. La pendiente <strong>de</strong> la regresión constituye un estimador <strong>de</strong>l coeficiente <strong>de</strong><br />
competencia.<br />
También basándonos en el mo<strong>de</strong>lo <strong>de</strong> Lotka-Volterra, po<strong>de</strong>mos estimar los coeficientes <strong>de</strong><br />
competencia mediante experiencias <strong>de</strong> remoción. Al disminuir los números <strong>de</strong> un competidor, la<br />
abundancia <strong>de</strong> una especie aumentará hasta un nuevo valor <strong>de</strong> equilibrio. El aumento relativo en los<br />
números <strong>de</strong> una población al extraer individuos <strong>de</strong> otra permite estimar el coeficiente <strong>de</strong> competencia<br />
(<strong>de</strong> la removida sobre la que respon<strong>de</strong>). El coeficiente <strong>de</strong> competencia se pue<strong>de</strong> interpretar como un<br />
factor <strong>de</strong> conversión <strong>de</strong> individuos <strong>de</strong> una especie en individuos <strong>de</strong> la otra.<br />
DESARROLLO DEL TRABAJO<br />
En el marco <strong>de</strong> la presente temática se realizará un trabajo experimental consi<strong>de</strong>rando dos géneros<br />
<strong>de</strong> plantas acuáticas flotantes <strong>de</strong> la familia Lemnaceae, <strong>de</strong>nominadas comúnmente “lentejas <strong>de</strong><br />
agua”: Spiro<strong>de</strong>lla sp. y Lemna sp. El objetivo <strong>de</strong>l experimento es analizar el efecto competitivo <strong>de</strong><br />
Spiro<strong>de</strong>lla sp. sobre el crecimiento (en número <strong>de</strong> fron<strong>de</strong>s) <strong>de</strong> Lemna sp. y viceversa, el efecto<br />
competitivo <strong>de</strong> Lemna sp. sobre el crecimiento <strong>de</strong> Spiro<strong>de</strong>lla sp. Para ello, se realizará un estudio<br />
piloto cultivando cada una <strong>de</strong> las especies en monocultivo y en presencia <strong>de</strong> la otra especie. Se<br />
utilizarán 18 ban<strong>de</strong>jas plásticas conteniendo 350 mL <strong>de</strong> una solución con nutrientes. Se realizarán 6<br />
réplicas conteniendo 10 fron<strong>de</strong>s iniciales <strong>de</strong> Lemna sp., 6 réplicas conteniendo 10 fron<strong>de</strong>s iniciales <strong>de</strong><br />
Spiro<strong>de</strong>lla sp., y 6 réplicas conteniendo 10 fron<strong>de</strong>s <strong>de</strong> cada una <strong>de</strong> las dos especies. Para cada<br />
ban<strong>de</strong>ja se efectuarán recuentos <strong>de</strong>l número <strong>de</strong> fron<strong>de</strong>s vivos una vez por semana, durante cuatro<br />
semanas.<br />
Nota: para las réplicas correspondientes al monocultivo <strong>de</strong> Lemna sp. se utilizarán las mismas<br />
unida<strong>de</strong>s experimentales que para el estudio <strong>de</strong> recursos y condiciones correspondientes al<br />
tratamiento <strong>de</strong> luz y la concentración C1.<br />
En base a los datos obtenidos:
73<br />
Ecología General<br />
1) Calcular y graficar el N promedio semanal <strong>de</strong> fron<strong>de</strong>s <strong>de</strong> Spiro<strong>de</strong>lla sp, y Lemna sp. en monocultivo<br />
y en competencia. Hacer un gráfico para cada especie en ambas situaciones.<br />
2) Consi<strong>de</strong>rando los valores promedio, estimar para cada especie la tasa <strong>de</strong> reproducción per cápita<br />
por unidad <strong>de</strong> tiempo (R) en monocultivo y la capacidad <strong>de</strong> carga (K) en monocultivo y competencia.<br />
¿A qué correspon<strong>de</strong> el K estimado para cada una en la situación <strong>de</strong> competencia?.<br />
3) Consi<strong>de</strong>rando los valores obtenidos para cada especie y el mo<strong>de</strong>lo <strong>de</strong> competencia <strong>de</strong> Lotka-<br />
Volterra: N1(eq) = K1 - α12* N2(eq) y N2(eq) = K2 - α21* N1(eq), estime los coeficientes <strong>de</strong><br />
competencia para cada una respecto <strong>de</strong> la otra.<br />
PARTE 3: Competencia. Ejercicios.<br />
Problema 1<br />
I. G.F.Gause, biólogo ruso, realizó la experiencia <strong>de</strong> hacer crecer a dos especies <strong>de</strong> levadura por<br />
separado y en conjunto, en un medio cultivo idéntico (un azúcar). Cuando crecían separadamente,<br />
observó que el crecimiento cesaba antes <strong>de</strong> agotarse totalmente el azúcar, aproximadamente en el<br />
mismo tiempo. Para la especie 1, estimó r = 0.22, observando que la <strong>de</strong>nsidad se estabilizaba en 13<br />
células por unidad <strong>de</strong> volumen. Para la especie 2, r = 0.6, con una <strong>de</strong>nsidad al final <strong>de</strong> la experiencia<br />
<strong>de</strong> 6 células por unidad <strong>de</strong> volumen. Cuando crecían juntas, observó que se afectaban negativamente<br />
calculando un α = 3.15 para la primer levadura y un β = 0.439 para la segunda.<br />
• Podría Ud <strong>de</strong>scribir qué sucedió con el sistema <strong>de</strong> las dos especies <strong>de</strong> levaduras interactuando.<br />
• Podría Ud interpretar biológicamente los resultados obtenidos.<br />
II. En otro experimento, Gause (1934) trabajó con tres especies <strong>de</strong> Paramecium: P. caudatum, P.<br />
aurelia y P. bursaria. Cuando eran cultivados por separado bajo condiciones idénticas observó que<br />
cada especie crecía bien, alcanzando una <strong>de</strong>nsidad máxima aproximadamente <strong>de</strong>spués <strong>de</strong> unos 10<br />
días <strong>de</strong> cultivo. Los parámetros r y K para cada especie fueron:<br />
especie r K<br />
P. caudatum 0.94 64<br />
P. aurelia 1.24 105<br />
P. bursaria 0.89 74<br />
Luego realizó las siguientes experiencias, don<strong>de</strong> dos especies <strong>de</strong> paramecios crecían juntas.<br />
Experiencia 1:<br />
Cuando crecían conjuntamente P. caudatum (especie 1) con P. aurelia (especie 2), pudo observar<br />
que se afectaban negativamente, calculando: α = 0.7 y β = 0.5.<br />
Experiencia 2:<br />
Cuando crecían conjuntamente P. caudatum (especie 1) con P. bursaria (especie 2), pudo calcular α<br />
= 0.5 y β = 0.5.<br />
• ¿Podría Ud <strong>de</strong>scribir qué sucedió con el sistema <strong>de</strong> las dos especies <strong>de</strong> paramecios interactuando<br />
en cada una <strong>de</strong> las experiencias?<br />
• ¿Podría Ud interpretar biológicamente los resultados obtenidos?<br />
Problema 2<br />
1. Graficar cualitativamente las curvas <strong>de</strong> crecimiento <strong>de</strong> las 2 especies (1 y 2) hasta el equilibrio a<br />
partir <strong>de</strong> las condiciones iniciales señaladas en los gráficos que se muestran y completar los<br />
cuadros vacíos.<br />
2. Indicar los puntos <strong>de</strong> equilibrio inestables o estables según corresponda en cada situación.<br />
3. Indicar según el mo<strong>de</strong>lo <strong>de</strong> L-V, en que situación (I o II) la competencia intraespecífica es mayor<br />
que la interespecífica para ambas especies
74<br />
Ecología General
Trabajo práctico 8<br />
75<br />
Ecología General<br />
INTERACCIÓN PREDADOR-PRESA: ESTUDIO DE LA RESPUESTA<br />
FUNCIONAL<br />
INTRODUCCIÓN<br />
La distribución y abundancia <strong>de</strong> los organismos están <strong>de</strong>terminadas por diversos factores,<br />
por ejemplo la disponibilidad <strong>de</strong> alimento, las condiciones fisicoquímicas, la capacidad <strong>de</strong><br />
dispersión, la competencia y las interacciones predador-presa.<br />
La predación es el consumo <strong>de</strong> un organismo (la presa) por parte <strong>de</strong> otro organismo (el<br />
predador), estando viva la presa cuando el <strong>de</strong>predador ataca por primera vez. La magnitud<br />
<strong>de</strong>l impacto <strong>de</strong> la predación sobre las poblaciones <strong>de</strong> presas <strong>de</strong>pen<strong>de</strong> por un lado <strong>de</strong> la<br />
cantidad <strong>de</strong> predadores, y por el otro <strong>de</strong> la capacidad <strong>de</strong> estos para encontrar y consumir a<br />
las presas.<br />
La predación es consi<strong>de</strong>rada una <strong>de</strong> las interacciones biológicas más importantes en<br />
algunas comunida<strong>de</strong>s, y diversos estudios <strong>de</strong>mostraron que por ejemplo los<br />
macroinvertebrados y los peces cumplen un rol importante en la estructuración <strong>de</strong> las<br />
comunida<strong>de</strong>s acuáticas, afectando significativamente las abundancias <strong>de</strong> las presas.<br />
En el estudio <strong>de</strong> la interacción predador-presa, uno <strong>de</strong> los aspectos más importantes es la<br />
respuesta <strong>de</strong> los predadores a los cambios en la <strong>de</strong>nsidad <strong>de</strong> presas, ya que por regla<br />
general, cuanto mas alta es la <strong>de</strong>nsidad <strong>de</strong>l alimento mayor es el consumo <strong>de</strong>l predador, y a<br />
mayor consumo más se incrementa su supervivencia y/o reproducción. Entonces se pue<strong>de</strong>n<br />
distinguir básicamente dos tipos <strong>de</strong> respuesta <strong>de</strong> los predadores a la <strong>de</strong>nsidad <strong>de</strong> presas: la<br />
respuesta numérica y la respuesta funcional<br />
La respuesta numérica se relaciona con el número <strong>de</strong> predadores, y se <strong>de</strong>fine como la<br />
variación <strong>de</strong> la <strong>de</strong>nsidad <strong>de</strong> predadores <strong>de</strong>bida a las variaciones en la <strong>de</strong>nsidad <strong>de</strong> presas.<br />
Estas variaciones se producen por cambios en la reproducción y/o la supervivencia <strong>de</strong>l<br />
predador frente a cambios en la <strong>de</strong>nsidad <strong>de</strong> presas, y se expresan en una escala <strong>de</strong> tiempo<br />
comparable con el tiempo generacional <strong>de</strong>l predador.<br />
La respuesta funcional se relaciona con el consumo <strong>de</strong>l predador, y se <strong>de</strong>fine como la<br />
variación en la tasa <strong>de</strong> consumo (número <strong>de</strong> presas consumidas por predador por unidad <strong>de</strong><br />
tiempo) en función <strong>de</strong> la <strong>de</strong>nsidad <strong>de</strong> presas. Holling en 1959 diferenció tres tipos <strong>de</strong><br />
respuesta funcional: las respuestas <strong>de</strong> tipo I, II y III.
76<br />
Ecología General<br />
Respuesta <strong>de</strong> tipo I<br />
El número <strong>de</strong> presas consumidas (Nc) por unidad <strong>de</strong> tiempo (T) aumenta en forma lineal con<br />
el número <strong>de</strong> presas disponibles (N), hasta que alcanza un máximo, a partir <strong>de</strong>l cual la tasa<br />
<strong>de</strong> consumo permanece constante (Figura A). En este caso la constante a (la tasa <strong>de</strong><br />
ataque) representa la proporción <strong>de</strong> presas atacadas (comidas) por unidad <strong>de</strong> tiempo (T).<br />
Nc = N* T * a<br />
Un ejemplo <strong>de</strong> este tipo <strong>de</strong> respuesta se pue<strong>de</strong> observar en los organismos filtradores, por<br />
ejemplo la tasa con que Daphnia magna consume células <strong>de</strong> levadura. Esto se <strong>de</strong>be a que<br />
las células <strong>de</strong> levadura son extraídas <strong>de</strong> un volumen constante <strong>de</strong> agua que pasa a través<br />
<strong>de</strong> su aparato filtrador. La cantidad <strong>de</strong> células consumidas se incrementa en forma<br />
constante con la concentración <strong>de</strong> las mismas, hasta que se alcanza el máximo que el<br />
predador pue<strong>de</strong> procesar, y a partir <strong>de</strong> ahí la tasa <strong>de</strong> consumo permanece constante.<br />
Respuesta <strong>de</strong> tipo II<br />
Es la respuesta más frecuente en la naturaleza. La tasa <strong>de</strong> consumo aumenta con la<br />
<strong>de</strong>nsidad <strong>de</strong> la presa, pero la velocidad <strong>de</strong>l aumento disminuye hasta alcanzar una<br />
plataforma en la cual la tasa <strong>de</strong> consumo permanece constante, in<strong>de</strong>pendientemente <strong>de</strong> la<br />
<strong>de</strong>nsidad <strong>de</strong> presa disponible (Figura A). Esta disminución se <strong>de</strong>be a que el predador utiliza<br />
una parte <strong>de</strong>l tiempo total (T) para manipular cada presa que consume (Tm, tiempo <strong>de</strong><br />
manipuleo), que consiste en perseguir, dominar y consumir a la presa y prepararse para la<br />
siguiente búsqueda. A medida que aumenta la <strong>de</strong>nsidad <strong>de</strong> las presas, el tiempo <strong>de</strong><br />
búsqueda disminuye, pero el tiempo <strong>de</strong> manipulación permanece constante y, por lo tanto,<br />
ocupa una proporción cada vez mayor <strong>de</strong>l tiempo <strong>de</strong>l consumidor. Cuando la <strong>de</strong>nsidad <strong>de</strong><br />
las presas es muy alta, el predador se pasa prácticamente todo el tiempo manipulándolas y<br />
su tasa <strong>de</strong> consumo se vuelve constante, <strong>de</strong>terminada por la cantidad <strong>de</strong> presas que un<br />
<strong>de</strong>predador pue<strong>de</strong> manipular durante el tiempo disponible. Este tipo <strong>de</strong> respuesta funcional<br />
ha sido observado frecuentemente en insectos, pero también en algunos vertebrados como<br />
el bisonte. La ecuación que representa este tipo <strong>de</strong> respuesta es la “ecuación <strong>de</strong> los discos<br />
<strong>de</strong> Holling”, y recor<strong>de</strong>mos que a es la constante que representa la tasa <strong>de</strong> ataque.<br />
Nc = N* T * a<br />
1 + a * Tm * N<br />
Respuesta <strong>de</strong> tipo III<br />
En este tipo <strong>de</strong> respuesta existe un incremento inicial en la tasa <strong>de</strong> consumo con el aumento<br />
<strong>de</strong> la <strong>de</strong>nsidad <strong>de</strong> presas hasta un punto <strong>de</strong> inflexión, en la cual comienza una<br />
<strong>de</strong>saceleración hasta alcanzar un plateau similar al que se da en la respuesta <strong>de</strong> tipo II<br />
(Figura A). En conjunto, este tipo <strong>de</strong> respuesta tiene una curva sigmoidal, y se pue<strong>de</strong><br />
producir por ejemplo cuando el incremento en la <strong>de</strong>nsidad <strong>de</strong> alimento lleva a un aumento<br />
en la eficiencia <strong>de</strong> búsqueda <strong>de</strong>l predador o a una disminución <strong>de</strong> su tiempo <strong>de</strong><br />
manipulación. Este tipo <strong>de</strong> respuesta ha sido observado tanto en invertebrados como en<br />
vertebrados, y entre las explicaciones posibles se incluye la heterogeneidad <strong>de</strong>l hábitat<br />
(presentando refugio pero limitado para pocas presas), falta <strong>de</strong> imagen <strong>de</strong> búsqueda a<br />
<strong>de</strong>nsida<strong>de</strong>s <strong>de</strong> presa bajas, y el cambio a presas alternativas cuando la abundancia <strong>de</strong><br />
presas es muy baja. Una <strong>de</strong> las ecuaciones más sencillas que representan este tipo <strong>de</strong><br />
respuesta es la siguiente:<br />
Nc = T * b * N 2
1 + b * N 2 * Tm<br />
77<br />
Ecología General<br />
Esta ecuación es similar a la <strong>de</strong> tipo II, pero a (tasa <strong>de</strong> ataque) no es constante, y se<br />
representa como b * N, siendo b una constante.<br />
40<br />
Número <strong>de</strong> presas<br />
consumidas<br />
Figura A. Respuesta funcional <strong>de</strong> Tipo I, Tipo II y Tipo III, según Holling (1959)<br />
Los distintos tipos <strong>de</strong> respuesta funcional afectan la dinámica <strong>de</strong> las poblaciones <strong>de</strong> los<br />
predadores y <strong>de</strong> las presas. Si el aumento <strong>de</strong> la tasa <strong>de</strong> consumo se <strong>de</strong>sacelera a medida<br />
que se incrementa la <strong>de</strong>nsidad <strong>de</strong>l alimento (respuesta <strong>de</strong> tipo II, respuesta <strong>de</strong> tipo I una vez<br />
alcanzada la plataforma, y respuesta <strong>de</strong>l tipo III a <strong>de</strong>nsida<strong>de</strong>s elevadas), las presas <strong>de</strong> las<br />
poblaciones con alta <strong>de</strong>nsidad tendrán menos probabilida<strong>de</strong>s <strong>de</strong> verse afectadas que las<br />
presas a <strong>de</strong>nsida<strong>de</strong>s bajas. Esto tiene un efecto <strong>de</strong>sestabilizador sobre la dinámica <strong>de</strong> las<br />
poblaciones, llevando a las presas a la extinción o permitiendoles escapar al control <strong>de</strong> los<br />
predadores. Si, por el contrario, el aumento <strong>de</strong> la tasa <strong>de</strong> consumo se acelera a medida que<br />
aumenta la <strong>de</strong>nsidad <strong>de</strong>l alimento (respuesta <strong>de</strong>l tipo III a <strong>de</strong>nsida<strong>de</strong>s bajas), se ven más<br />
afectadas las presas a <strong>de</strong>nsida<strong>de</strong>s altas, lo que tien<strong>de</strong> a estabilizar la dinámica <strong>de</strong> las<br />
presas ya que se produce una mortalidad <strong>de</strong>nso<strong>de</strong>pendiente. Esto es especialmente cierto<br />
en el caso <strong>de</strong> predadores generalistas (Hassell 1978).<br />
Las investigaciones biológicas sobre la respuesta funcional generalmente intentan<br />
respon<strong>de</strong>r las siguientes preguntas:<br />
a) ¿Qué tipo <strong>de</strong> respuesta funcional tiene un predador? La respuesta a esta pregunta es <strong>de</strong><br />
interés cuando se intenta <strong>de</strong>terminar si la predación <strong>de</strong>nso-<strong>de</strong>pendiente es un factor<br />
estabilizador para las poblaciones <strong>de</strong> presas expuestas al predador.<br />
b) ¿Cuáles son los valores estimados para los parámetros? Esta pregunta <strong>de</strong>be ser<br />
respondida para po<strong>de</strong>r utilizar la respuesta funcional en los mo<strong>de</strong>los mecanísticos <strong>de</strong> la<br />
dinámica predador-presa.<br />
Para po<strong>de</strong>r distinguir a partir <strong>de</strong> datos experimentales entre una respuesta <strong>de</strong> tipo II y una <strong>de</strong><br />
tipo III no es suficiente con ajustar los datos a las ecuaciones correspondientes y <strong>de</strong>cidir cuál<br />
ajusta mejor. De hecho las diferencias entre las curvas pue<strong>de</strong>n ser sutiles y difíciles <strong>de</strong><br />
<strong>de</strong>terminar. Una manera <strong>de</strong> po<strong>de</strong>r diferenciar entre ambos mo<strong>de</strong>los es analizar la proporción<br />
<strong>de</strong> presas consumidas a distintas <strong>de</strong>nsida<strong>de</strong>s. Las curvas para cada tipo <strong>de</strong> respuesta<br />
funcional difieren en la pendiente a bajas <strong>de</strong>nsida<strong>de</strong>s (Figura B).<br />
Proporción <strong>de</strong> presas<br />
consumidas<br />
Tipo I<br />
30<br />
Tipo II<br />
0<br />
0<br />
0<br />
0 180 0 Densidad <strong>de</strong> presas 180 0 180<br />
Tipo I Tipo II Tipo III<br />
Densidad <strong>de</strong> presas<br />
50<br />
Tipo III
78<br />
Ecología General<br />
Figura B. Proporción <strong>de</strong> presas consumidas en función <strong>de</strong> la <strong>de</strong>nsidad para los tres tipos <strong>de</strong><br />
respuesta funcional<br />
Con estas curvas resulta mucho más sencillo diferenciar especialmente entre las respuestas<br />
<strong>de</strong> tipo II y <strong>de</strong> tipo III. Para la respuesta funcional <strong>de</strong> tipo II se observa una disminución en la<br />
proporción <strong>de</strong> presas consumidas en función <strong>de</strong> la <strong>de</strong>nsidad <strong>de</strong> presas, mientras que para la<br />
respuesta <strong>de</strong> tipo III a bajas <strong>de</strong>nsida<strong>de</strong>s la proporción <strong>de</strong> presas consumidas se incrementa<br />
hasta alcanzar un máximo, y luego disminuye en forma similar a lo observado para la<br />
respuesta <strong>de</strong> tipo II.<br />
Una vez i<strong>de</strong>ntificada la forma general <strong>de</strong> la curva se utiliza el mo<strong>de</strong>lo correspondiente para<br />
estimar los parámetros <strong>de</strong> la respuesta funcional. Esto se hace mediante una regresión no<br />
lineal <strong>de</strong>l número <strong>de</strong> presas consumidas versus el número <strong>de</strong> presas ofrecidas (Juliano<br />
2001).<br />
DESARROLLO DEL TRABAJO PRÁCTICO<br />
Objetivos<br />
• Estudiar experimentalmente la respuesta funcional <strong>de</strong> un predador.<br />
• Discutir sobre los factores que podrían modificar la respuesta funcional observada.<br />
<strong>Primer</strong>a clase: realización <strong>de</strong>l experimento<br />
Los organismos que se utilizarán en los ensayos serán:<br />
• Predador: Buenoa fuscipennis: es un insecto acuático <strong>de</strong> la familia Notonectidae<br />
(Heteroptera), que es muy frecuente en distintos cuerpos <strong>de</strong> agua <strong>de</strong> la región (lagunas,<br />
charcos temporarios y piletas <strong>de</strong> natación). Su hábito alimentario es predador, y se<br />
alimenta <strong>de</strong> macroinvertebrados <strong>de</strong> pequeño tamaño, como por ejemplo larvas <strong>de</strong><br />
mosquitos, cladóceros, etc.<br />
• Presa: larvas <strong>de</strong> mosquito <strong>de</strong> segundo o tercer estadio: se utilizará la especie Ae<strong>de</strong>s<br />
aegypti <strong>de</strong>bido a la facilidad con que la misma se pue<strong>de</strong> criar en cautiverio, permitiendo<br />
obtener un número suficiente <strong>de</strong> ejemplares <strong>de</strong>l mismo tamaño para realizar el<br />
experimento.<br />
Los ensayos se realizarán en recipientes <strong>de</strong> 4 lts, conteniendo 3 lts <strong>de</strong> agua. En cada uno se<br />
colocará un predador y una <strong>de</strong> seis <strong>de</strong>nsida<strong>de</strong>s crecientes <strong>de</strong> presas. Las <strong>de</strong>nsida<strong>de</strong>s <strong>de</strong><br />
presa que se utilizarán en el ensayo serán: 8, 12, 18, 25, 40 y 75 larvas <strong>de</strong> mosquito. Para<br />
cada <strong>de</strong>nsidad <strong>de</strong> presas se harán nueve réplicas. Debido a que la mortalidad <strong>de</strong> las presas<br />
en condiciones <strong>de</strong> laboratorio es <strong>de</strong>spreciable en el tiempo que dura la experiencia, no se<br />
realizarán controles (sin predador).<br />
Luego <strong>de</strong> dos horas se retirará el predador y se contará el número <strong>de</strong> presas vivas en cada<br />
recipiente. A partir <strong>de</strong> estos datos se calculará el número <strong>de</strong> presas consumidas como el
79<br />
Ecología General<br />
número inicial menos el número final <strong>de</strong> presas vivas. Los resultados serán registrados en la<br />
planilla adjunta en este práctico.<br />
Segunda clase: análisis y discusión <strong>de</strong> los resultados<br />
En forma grupal se realizarán las siguientes activida<strong>de</strong>s, que posteriormente serán<br />
discutidas con todo el curso.<br />
a) Planteo <strong>de</strong> la hipótesis<br />
Cada grupo discutirá y pondrá por escrito la hipótesis y qué resultados espera obtener en<br />
este experimento en función <strong>de</strong> las hipótesis planteadas.<br />
b) Determinación <strong>de</strong>l tipo <strong>de</strong> respuesta funcional <strong>de</strong> Buenoa fuscipennis a partir <strong>de</strong> los<br />
datos experimentales<br />
• Graficar el número <strong>de</strong> presas consumidas en función <strong>de</strong> la <strong>de</strong>nsidad <strong>de</strong> presas. ¿Se<br />
pue<strong>de</strong> establecer el tipo <strong>de</strong> respuesta funcional? ¿Por qué?<br />
• Graficar la proporción <strong>de</strong> presas consumidas en función <strong>de</strong> la <strong>de</strong>nsidad <strong>de</strong> presas y<br />
mirar especialmente la pendiente a <strong>de</strong>nsida<strong>de</strong>s bajas <strong>de</strong> presas. ¿Qué tipo <strong>de</strong><br />
respuesta funcional se observó para este predador?<br />
c) Discusión <strong>de</strong> los resultados en función <strong>de</strong> los siguientes puntos<br />
1. Tipo <strong>de</strong> respuesta funcional:<br />
• ¿Coinci<strong>de</strong>n los resultados obtenidos con los esperados en función <strong>de</strong> la hipótesis<br />
planteada?<br />
• ¿Por qué se analiza la proporción <strong>de</strong> larvas consumidas para <strong>de</strong>terminar el tipo <strong>de</strong><br />
respuesta funcional?<br />
2. Análisis <strong>de</strong> los parámetros:<br />
• ¿Qué significado tienen los parámetros a y Tm (RF tipo II) y los parámetros b y Tm<br />
(RF tipo III)?<br />
• ¿Cómo se pue<strong>de</strong> estimar el máximo número <strong>de</strong> presas que un predador pue<strong>de</strong><br />
capturar en un tiempo <strong>de</strong>terminado?<br />
• Teniendo en cuenta los parámetros obtenidos para los resultados experimentales<br />
(mediante regresiones no lineales por el método <strong>de</strong> cuadrados mínimos), cuál es el
80<br />
Ecología General<br />
máximo número <strong>de</strong> presas que pue<strong>de</strong> consumir Buenoa fuscipennis? ¿A qué<br />
intervalo <strong>de</strong> tiempo se refiere este número?<br />
3. Consecuencias sobre las poblaciones:<br />
• Discutir por qué la respuesta funcional <strong>de</strong> tipo II es <strong>de</strong>sestabilizadora mientras que la<br />
respuesta funcional <strong>de</strong> tipo III es estabilizadora bajo ciertas condiciones<br />
• ¿Cuáles son las condiciones que pue<strong>de</strong>n incrementar la estabilidad <strong>de</strong> las<br />
interacciones predador-presa?<br />
4. Análisis <strong>de</strong> los supuestos <strong>de</strong>l mo<strong>de</strong>lo:<br />
• Discutir para cada uno <strong>de</strong> los siguientes supuestos <strong>de</strong> los mo<strong>de</strong>los <strong>de</strong> respuesta<br />
funcional <strong>de</strong> tipo II y III <strong>de</strong> Holling:<br />
la asíntota <strong>de</strong> la curva está <strong>de</strong>terminada por la limitación <strong>de</strong> tiempo<br />
los predadores capturan y consumen todo lo que pue<strong>de</strong>n en ese tiempo<br />
la tasa <strong>de</strong> encuentro <strong>de</strong> los predadores con las presas se relaciona linealmente<br />
con la <strong>de</strong>nsidad <strong>de</strong> presas<br />
los predadores no hacen capturas adicionales mientras manipulan una presa<br />
los predadores no están hambreados<br />
la <strong>de</strong>nsidad <strong>de</strong> presas permanece constante<br />
¿Se cumplió en el experimento realizado? En caso <strong>de</strong> respuesta negativa, sugiera<br />
los cambios a realizar en el diseño experimental para que se cumpla el supuesto.<br />
• Teniendo en cuenta los resultados <strong>de</strong>l trabajo <strong>de</strong> Ellis y Bor<strong>de</strong>n (1970) para<br />
Notonecta undulata? (ver figura 1), discutir: ¿Cómo influye el grado <strong>de</strong> saciedad <strong>de</strong>l<br />
predador sobre el número <strong>de</strong> larvas consumidas por unidad <strong>de</strong> tiempo? ¿qué<br />
resultados se hubieran obtenido en nuestro experimento si se hubieran analizado por<br />
separado intervalos <strong>de</strong> 30 minutos?<br />
Figura 1. Número medio <strong>de</strong> presas<br />
consumidas durante 10 horas por<br />
Notonecta undulata adultos, que<br />
fueron hambreados por 48 hs antes<br />
<strong>de</strong> ofrecerles cincuenta larvas <strong>de</strong><br />
cuarto estadio <strong>de</strong> Ae<strong>de</strong>s aegypti por<br />
hora. Las barras verticales son<br />
<strong>de</strong>svíos estándar. Extraído <strong>de</strong> Ellis y<br />
Bor<strong>de</strong>n 1970.
81<br />
Ecología General<br />
• Consi<strong>de</strong>rando los resultados <strong>de</strong>l trabajo <strong>de</strong> Chesson (1989), discutir: ¿Cuál es el<br />
efecto que tienen distintas presas alternativas como Daphnia (fig 2a) y Drosophila (fig<br />
2b) sobre la respuesta funcional <strong>de</strong> Notonecta hoffmani?<br />
Figura 2. Efecto <strong>de</strong> (a) Daphnia y <strong>de</strong><br />
(b) Drosophila sobre el número <strong>de</strong><br />
mosquitos consumidos en una hora<br />
por adultos <strong>de</strong> Notonecta hoffmani.<br />
Se muestran las medias ± error<br />
estándar. Extraído <strong>de</strong> Chesson<br />
1989.<br />
BIBLIOGRAFÍA GENERAL<br />
Begon M, Harper JL y Townsend CR (1995) Ecología. Individuos, poblaciones y<br />
comunida<strong>de</strong>s. Ediciones Omega, Barcelona.<br />
Chesson J. 1989. The effect of alternative prey on the functional response of Notonecta<br />
hoffmani. Ecology 70(5): 1227-1235.<br />
Ellis RA y Bor<strong>de</strong>n JH. 1970. Predation by Notonecta undulata (Heteroptera: Notonectidae)<br />
on Larvae of the Yellow-Fewer Mosquito. Annals of the Entomological Society of America<br />
63: 963-973.<br />
Fernan<strong>de</strong>z-Arhex V y Corley JC. 2004. La respuesta funcional: una revisión y guía<br />
experimental. Ecología Austral 14: 83-93.<br />
Hassell MP. 1978. The Dynamics of Arthropod Predator-Prey Systems. Princeton University<br />
Press, Princeton, NJ.<br />
(a)<br />
(b)
82<br />
Ecología General<br />
Juliano SA (2001) Nonlinear curve fitting. Predation and Functional Response Curves. En<br />
Design and Analysis of Ecological Experiments. Second Edition. Scheiner, SM y<br />
Gurevitch, J (eds). Oxford University Press, New York.<br />
Planilla <strong>de</strong> registro <strong>de</strong> los resultados por grupo<br />
Turno Grupo<br />
Número<br />
inicial <strong>de</strong><br />
presas<br />
Número <strong>de</strong><br />
presas vivas<br />
al final<br />
Número <strong>de</strong><br />
presas<br />
consumidas<br />
(inicial-final)<br />
Comentarios y observaciones
Trabajo práctico 9<br />
83<br />
Ecología General<br />
COMPARACION DE LAS TASAS DE DESCOMPOSICION FOLIAR<br />
DE DISTINTAS ESPECIES VEGETALES<br />
por Gabriela Mataloni<br />
Introducción<br />
Dentro <strong>de</strong> un ecosistema, los principales eslabones <strong>de</strong> la trama trófica son los organismos<br />
productores, los consumidores y los <strong>de</strong>scomponedores o <strong>de</strong>tritívoros. Estos últimos son aquellos<br />
organismos que se alimentan <strong>de</strong> restos <strong>de</strong> materia orgánica inerte o muerta (Smith & Smith, 2001).<br />
Los <strong>de</strong>scomponedores son muy abundantes. En sólo 1m 2 <strong>de</strong> suelo forestal <strong>de</strong> clima templado pue<strong>de</strong><br />
haber cerca <strong>de</strong> un millón <strong>de</strong> estos organismos, alre<strong>de</strong>dor <strong>de</strong> un 90% <strong>de</strong> los cuales son bacterias<br />
heterótrofas aeróbicas y hongos microscópicos (Fig. 1)<br />
protozoos<br />
hongos<br />
microscópicos<br />
macroorganismos<br />
hongos<br />
macroscópicos<br />
Fig. 1: Composición <strong>de</strong> la biota <strong>de</strong>tritívora<br />
bacterias<br />
heterótrofas<br />
Debe tenerse en cuenta que estas proporciones se refieren a número <strong>de</strong> individuos y no a su<br />
biomasa. De hecho, hay una relación complementaria entre macro y micro<strong>de</strong>scomponedores. Los<br />
macroorganismos fragmentan el material <strong>de</strong>trítico, facilitando su acceso a los micro<strong>de</strong>tritívoros,<br />
bacterias y hongos. En particular, las bacterias son los principales regeneradores <strong>de</strong> nutrientes en los<br />
ecosistemas terrestres, y concentran la mayor parte <strong>de</strong> la energía en su biomasa.<br />
Cuál es la importancia ecológica <strong>de</strong> los <strong>de</strong>scomponedores? Existen en todo ecosistema dos ca<strong>de</strong>nas<br />
o vías tróficas principales: la <strong>de</strong> los herbívoros, cuya fuente <strong>de</strong> energía es la producción primaria por<br />
fotosíntesis, y la <strong>de</strong> los <strong>de</strong>tritívoros, cuya fuente <strong>de</strong> energía es la materia orgánica muerta. Contra lo<br />
que las apariencias sugieren, es ésta última la principal vía para el flujo <strong>de</strong> energía, ya que sólo una<br />
pequeña parte <strong>de</strong> la producción <strong>de</strong>l ecosistema es utilizada por los herbívoros. Por ejemplo, <strong>de</strong> la<br />
cantidad total <strong>de</strong> energía fijada mediante fotosíntesis (producción bruta) en un bosque caducifolio<br />
templado, aproximadamente el 50% se <strong>de</strong>stina al mantenimiento y respiración; sólo un 13% es<br />
consumido por herbívoros y el 35% entra directamente en la ca<strong>de</strong>na alimentaria <strong>de</strong> los<br />
<strong>de</strong>scomponedores (Smith & Smith, 2001). De esta forma, la <strong>de</strong>scomposición <strong>de</strong>l material vegetal<br />
senescente es un proceso fundamental en el funcionamiento <strong>de</strong>l ecosistema, ya que <strong>de</strong> él <strong>de</strong>pen<strong>de</strong> el<br />
reciclado <strong>de</strong> nutrientes, y por lo tanto la productividad primaria y secundaria (Pérez Harguin<strong>de</strong>guy et<br />
al., 1997).<br />
El proceso <strong>de</strong> <strong>de</strong>scomposición está regulado principalmente por tres factores: las características<br />
físico-químicas <strong>de</strong>l ambiente (temperatura, humedad, pH, etc.), la composición <strong>de</strong> la biota edáfica
84<br />
Ecología General<br />
(Bradford et al., 2002), y la calidad <strong>de</strong>l material vegetal en <strong>de</strong>scomposición (Gallardo & Merino, 1993).<br />
Dado que esta última <strong>de</strong>pen<strong>de</strong>, a su vez, <strong>de</strong> la naturaleza química <strong>de</strong> las hojas, la que varía <strong>de</strong> una<br />
especie a otra, la composición florística <strong>de</strong> una <strong>de</strong>terminada comunidad vegetal influirá en el proceso<br />
<strong>de</strong> <strong>de</strong>scomposición <strong>de</strong>l sistema al que pertenece. Este hecho cobra especial importancia en relación<br />
con la expansión <strong>de</strong> especies invasoras en el contexto <strong>de</strong>l cambio climático global (Allison & Vitousek,<br />
2004).<br />
Objetivos<br />
1) Calcular las tasas <strong>de</strong> <strong>de</strong>scomposición para las hojas <strong>de</strong> tres especies <strong>de</strong> plantas <strong>de</strong> Ciudad<br />
Universitaria.<br />
2) Relacionar los resultados obtenidos con las características fenológicas y formas <strong>de</strong> vida <strong>de</strong> las<br />
especies estudiadas.<br />
3) Discutir la influencia <strong>de</strong> estos parámetros sobre el proceso <strong>de</strong> <strong>de</strong>scomposición para el ecosistema<br />
en general.<br />
Hipótesis<br />
1) Las hojas provenientes <strong>de</strong> plantas <strong>de</strong> diferentes formas <strong>de</strong> vida y características fenológicas tienen<br />
distintas tasas <strong>de</strong> <strong>de</strong>scomposición.<br />
2) La velocidad/tasa <strong>de</strong>scomposición tien<strong>de</strong> a ser mayor en especies herbáceas que en leñosas, y en<br />
especies caducifolias que perennifolias.<br />
Materiales necesarios por grupo<br />
- Un rectángulo <strong>de</strong> 10 x 20 cm <strong>de</strong> tela <strong>de</strong> mosquitero plástica, <strong>de</strong> 3 mm <strong>de</strong> abertura.<br />
- Abrochadora y ganchos metálicos<br />
- 3 trozos <strong>de</strong> goma EVA<br />
- Rotulador<br />
- 1 bolsa <strong>de</strong> papel<br />
- 1 trozo <strong>de</strong> cable <strong>de</strong> unos 30 cm <strong>de</strong> largo<br />
- 1 arco <strong>de</strong> alambre torcido en U<br />
Desarrollo<br />
Cada grupo <strong>de</strong> alumnos trabajará con hojas <strong>de</strong> una especie vegetal perteneciente a uno <strong>de</strong> estos tres<br />
tipos:<br />
1- herbácea<br />
2- árbol leñoso caducifolio<br />
3- árbol leñoso perennifolio<br />
Los docentes proveerán a cada grupo <strong>de</strong> dos bolsas <strong>de</strong> papel, cada una <strong>de</strong> ellas conteniendo hojas<br />
enteras <strong>de</strong> alguna <strong>de</strong> las tres especies, previamente secadas al aire durante 72 hs.<br />
Los alumnos rotularán ambas bolsas con los siguientes datos:<br />
Turno<br />
Grupo<br />
Especie<br />
Bolsa (E=experimental o T=testigo)<br />
y proce<strong>de</strong>rán a pesar el contenido <strong>de</strong> cada bolsa.<br />
Los datos obtenidos se volcarán en una planilla por turno, en la columna <strong>de</strong>nominada "Peso fresco"<br />
(ver Apéndice).<br />
La bolsa A se llevará a estufa a 70 °C durante 48 hs. Transcurrido este tiempo, su contenido se<br />
pesará nuevamente, y se anotará en la columna correspondiente a "Peso seco". En base a estos<br />
datos, se calculará el cociente Peso Seco <strong>de</strong>l Testigo (PS(T))/Peso Fresco testigo (PF(T)) para cada<br />
réplica. Se calculará el valor promedio <strong>de</strong> este cociente para cada tipo <strong>de</strong> hoja. Este valor se<br />
multiplica por el peso fresco experimental para calcular el peso seco inicial experimental.<br />
PSI (E) = PS (T) x PF (E)<br />
PF (T)<br />
don<strong>de</strong>: PS (T) = Peso seco <strong>de</strong>l contenido <strong>de</strong> la bolsa T<br />
PF (E), PF (T) = Peso fresco <strong>de</strong>l contenido <strong>de</strong> las bolsas E y T, respectivamente.
85<br />
Ecología General<br />
El contenido <strong>de</strong> la bolsa E se colocará cuidadosamente <strong>de</strong>ntro <strong>de</strong> un sobre <strong>de</strong> 10 x 10 cm <strong>de</strong> tela<br />
plástica <strong>de</strong> mosquitero (bolsa <strong>de</strong> <strong>de</strong>scomposición), el que se cerrará con ganchos metálicos <strong>de</strong><br />
abrochadora. En un rincón <strong>de</strong>l mismo se realizará una perforación a través <strong>de</strong> la que se pasará el<br />
cable, que en su otro extremo llevará una etiqueta hecha con una lámina <strong>de</strong> goma EVA. En esta<br />
etiqueta se colocarán los datos <strong>de</strong>l material (turno, grupo, especie). El dispositivo se completa con un<br />
alambre en forma <strong>de</strong> U con el que se lo fijará al suelo.<br />
El aspecto general es el que se muestra en la figura:<br />
Todos los dispositivos se trasladarán DENTRO DE LAS BOLSAS DE PAPEL RESPECTIVAS al<br />
campo experimental <strong>de</strong> la Ciudad Universitaria para su instalación.<br />
Allí habrá un área previamente <strong>de</strong>marcada, <strong>de</strong>ntro <strong>de</strong> la cual se instalarán todos los dispositivos a lo<br />
largo <strong>de</strong> líneas paralelas (una por turno), a intervalos <strong>de</strong> 30 cm <strong>de</strong> distancia entre sí. Dentro <strong>de</strong> cada<br />
turno los diferentes dispositivos (1 por grupo) se dispondrán al azar, sorteando su ubicación.<br />
ES IMPORTANTE QUE CADA TURNO REALICE UN ESQUEMA DE LA UBICACIÓN DE SUS<br />
DISPOSITIVOS EN EL CAMPO. Una vez <strong>de</strong>finidos los lugares, se levantará cuidadosamente el<br />
mantillo <strong>de</strong>l suelo (capa <strong>de</strong> materia orgánica en <strong>de</strong>scomposición <strong>de</strong> origen aún reconocible), se<br />
apoyará la bolsa <strong>de</strong> red sobre la superficie y se cubrirá nuevamente con el mantillo. Todo el<br />
dispositivo se asegurará mediante el alambre, y se dispondrán las etiquetas <strong>de</strong> goma EVA <strong>de</strong> forma<br />
bien visible, ya que cumplen la doble función <strong>de</strong> rótulo y marca en el terreno para recuperar el<br />
material.<br />
Luego <strong>de</strong> 10 a 12 semanas, se recuperarán cuidadosamente los dispositivos y trasladarán al<br />
laboratorio <strong>de</strong>ntro <strong>de</strong> las bolsas <strong>de</strong> papel. Se abrirán <strong>de</strong>ntro <strong>de</strong> ban<strong>de</strong>jas plásticas y con pinza y pincel<br />
se separarán los remanentes <strong>de</strong> las hojas utilizadas en el experimento <strong>de</strong> todo otro tipo <strong>de</strong> material.<br />
Los restos <strong>de</strong> hojas se colocarán en bolsas <strong>de</strong> papel pequeñas, las que se rotularán cuidadosamente<br />
y llevarán a estufa durante 48 hs. Después <strong>de</strong> este tiempo, se medirá el peso seco (PSF) <strong>de</strong> la<br />
hojarasca contenida en cada una <strong>de</strong> ellas.<br />
La tasa promedio <strong>de</strong> <strong>de</strong>scomposición diaria se calculará como:<br />
D= PSI - PSF x 100 .<br />
PSI N° <strong>de</strong> días<br />
don<strong>de</strong>: D= tasa promedio <strong>de</strong> <strong>de</strong>scomposición porcentual diaria<br />
PSI= peso seco inicial<br />
PSF= peso seco final<br />
Unida<strong>de</strong>s <strong>de</strong> D: % <strong>de</strong> peso seco / día<br />
Nota: Por razones <strong>de</strong> comodidad, calculamos una tasa diaria promedio, pero esto no significa que la<br />
tasa <strong>de</strong> <strong>de</strong>scomposición sea constante a lo largo <strong>de</strong>l tiempo. ¿Cómo diseñaría un experimento para<br />
comprobar esto?<br />
Análisis <strong>de</strong> los datos
86<br />
Ecología General<br />
Se consignarán los datos <strong>de</strong> cada turno en la siguiente tabla, reuniéndose luego los datos <strong>de</strong> todos<br />
los turnos para su análisis:<br />
Grupo:<br />
Grupo:<br />
Grupo:<br />
Grupo:<br />
Grupo:<br />
Grupo:<br />
Grupo:<br />
Grupo:<br />
Grupo:<br />
Grupo:<br />
D herbácea D árbol caducifolio D árbol<br />
perennifolio<br />
Se calculará la media y la <strong>de</strong>sviación estandar <strong>de</strong> la tasa <strong>de</strong> <strong>de</strong>scomposición <strong>de</strong> cada especie, las<br />
que se graficarán en un histograma.<br />
Se realizará un ANOVA <strong>de</strong> un factor (previa transformación <strong>de</strong> la variable en caso <strong>de</strong> no cumplir con<br />
los supuestos) para comprobar si hay diferencias significativas entre las tasas <strong>de</strong> <strong>de</strong>scomposición.<br />
Cuál es, en este caso, el factor? Y los niveles?<br />
En caso <strong>de</strong> rechazarse:<br />
H0= todas las medias <strong>de</strong> las tasas <strong>de</strong> <strong>de</strong>scomposición son iguales<br />
se realizarán contrastes a posteriori para <strong>de</strong>terminar cuál o cuáles son las especies con tasas <strong>de</strong><br />
<strong>de</strong>scomposición significativamente diferentes.<br />
De no cumplirse los supuestos para realizar un ANOVA, las diferencias entre tasas <strong>de</strong><br />
<strong>de</strong>scomposición se analizarán mediante un test no paramétrico <strong>de</strong> Kruskal-Wallis, con contrastes a<br />
posteriori por el método <strong>de</strong> Dunn (Zar, 1996).<br />
De acuerdo a los resultados obtenidos, discuta:<br />
- Se relacionan las tasas <strong>de</strong> <strong>de</strong>scomposición con la fenología y/o con la forma <strong>de</strong> vida <strong>de</strong> las<br />
especies?<br />
- Según su respuesta anterior, cómo piensa Ud. que influiría un cambio en la composición <strong>de</strong><br />
especies en el funcionamiento <strong>de</strong>l ecosistema?<br />
Bibliografía consultada
87<br />
Ecología General<br />
Allison, S.D. & P.M. Vitousek, 2004: Rapid nutrient cycling in leaf litter from invasive plants in Hawai'i.<br />
Oecologia 141: 612-619.<br />
Bradford, M.A., G.M. Tordoff, T. Elgers, T. Hefin Jones & J.E. Newington, 2002: Microbiota, fauna and<br />
mesh size interactions in litter <strong>de</strong>composition. Oikos 99: 3117-323.<br />
Cornelissen, J.H.C., 1996: An experimental comparison of leaf <strong>de</strong>composition rates in a wi<strong>de</strong> range of<br />
temperate plant species and types. Journal of Ecology, 84: 573-582.<br />
Gallardo, A. & J. Merino, 1993: Leaf <strong>de</strong>composition in two mediterranean ecosystems of southwest<br />
Spain: Influence of substrate quality. Ecology, 74(1): 152-161.<br />
Pérez Harguin<strong>de</strong>guy, N., S. Díaz, J.H.C. Cornelissen & M. Cabido, 1997: Comparación experimental<br />
<strong>de</strong> la tasa <strong>de</strong> <strong>de</strong>scomposición foliar <strong>de</strong> especies vegetales <strong>de</strong>l centro-oeste <strong>de</strong> Argentina. Ecología<br />
Austral 7: 87-94.<br />
Smith, R.L. & T. M. Smith, 2001: Ecología. 4ta. Edición. Pearson Educación, Madrid. 664 pp.<br />
Zar, J.H., 1996: Biostatistical Analysis. Prentice-Hall, New Jersey. 662 pp.
Apéndice: Tabla 1<br />
Debe ser entregada por los docentes <strong>de</strong> cada turno al coordinador <strong>de</strong>l TP<br />
88<br />
Ecología General<br />
TURNO Peso fresco Peso fresco Peso seco Peso seco<br />
bolsa A bolsa B bolsa A bolsa B<br />
Grupo:<br />
Especie:<br />
Grupo:<br />
Especie:<br />
Grupo:<br />
Especie:<br />
Grupo:<br />
Especie:<br />
Grupo:<br />
Especie:<br />
Grupo:<br />
Especie:<br />
Grupo:<br />
Especie:<br />
Grupo:<br />
Especie:<br />
Grupo:<br />
Especie:<br />
Grupo:<br />
Especie:
Trabajo práctico 10<br />
ATRIBUTOS DE LAS COMUNIDADES<br />
INTRODUCCIÓN<br />
89<br />
Ecología General<br />
En los libros <strong>de</strong> texto <strong>de</strong> ecología se pue<strong>de</strong>n encontrar varias <strong>de</strong>finiciones <strong>de</strong> “comunidad”, que<br />
cubren un rango consi<strong>de</strong>rable <strong>de</strong> significados. Algunos consi<strong>de</strong>ran a la comunidad como “un<br />
ensamble <strong>de</strong> poblaciones <strong>de</strong> plantas, animales, bacterias y hongos que viven en un ambiente y que<br />
interactúan unos con otros, formando juntos un sistema viviente distintivo con su propia composición,<br />
estructura, relaciones ambientales, <strong>de</strong>sarrollo y función” (Whittaker 1975). En el otro extremo, se la ha<br />
consi<strong>de</strong>rado como “cualquier conjunto <strong>de</strong> organismos que viven cerca unos <strong>de</strong> otros y acerca <strong>de</strong> los<br />
cuales es interesante hablar” (MacArthur 1971). Todas las <strong>de</strong>finiciones, no obstante, concuerdan en<br />
que las comunida<strong>de</strong>s son conjuntos <strong>de</strong> individuos <strong>de</strong> distintas especies que aparecen juntos en<br />
tiempo y espacio, y la mayoría <strong>de</strong>staca la importancia <strong>de</strong> las interacciones entre esas poblaciones.<br />
Varios autores, por otra parte, señalan la existencia <strong>de</strong> propieda<strong>de</strong>s emergentes <strong>de</strong> las comunida<strong>de</strong>s,<br />
atributos <strong>de</strong> estructura (e.g., la composición <strong>de</strong> especies) o <strong>de</strong> funcionamiento (e.g., el flujo <strong>de</strong><br />
energía) que son característicos <strong>de</strong> este nivel <strong>de</strong> organización.<br />
En alguna medida, las diferentes <strong>de</strong>finiciones <strong>de</strong> comunidad son consecuencia <strong>de</strong> los distintos<br />
objetivos <strong>de</strong> los investigadores que las propusieron. Los ecólogos <strong>de</strong> plantas, que tratan con<br />
ensambles espacialmente fijos, a menudo enfatizan la <strong>de</strong>scripción <strong>de</strong> tales asociaciones y sus<br />
cambios en el tiempo; los ecólogos <strong>de</strong> animales, confrontados con organismos móviles y activos, le<br />
dan más importancia a las interacciones y a las relaciones funcionales entre las especies. Algunos<br />
<strong>de</strong>finen a la comunidad en términos <strong>de</strong> unida<strong>de</strong>s <strong>de</strong> hábitat (e.g., las comunida<strong>de</strong>s <strong>de</strong>l intermareal),<br />
otros por categorías <strong>de</strong> formas <strong>de</strong> vida (e.g., comunida<strong>de</strong>s herbáceas) o por taxonomía (e.g.,<br />
comunida<strong>de</strong>s <strong>de</strong> aves). Lo común a todas estas formas <strong>de</strong> <strong>de</strong>finir a una comunidad es su valor<br />
operativo: todas se centran en una parte <strong>de</strong>l conjunto total <strong>de</strong> especies que coexisten, pues es<br />
prácticamente imposible trabajar con el concepto original <strong>de</strong> comunidad (i.e., el conjunto <strong>de</strong> todos los<br />
individuos <strong>de</strong> todas las especies que viven en un <strong>de</strong>terminado lugar). Es muy claro que la noción <strong>de</strong><br />
comunidad, aún cuando se utilicen solo formas “operativas”, ha contribuido notablemente al <strong>de</strong>sarrollo<br />
<strong>de</strong> nuestro entendimiento <strong>de</strong> la naturaleza (Wiens 1989).<br />
Los atributos comunitarios más comúnmente utilizados por los ecólogos son los siguientes:<br />
• Diversidad específica: es una función <strong>de</strong> la riqueza específica (número <strong>de</strong> especies presentes) y<br />
<strong>de</strong> la equitatividad (grado <strong>de</strong> uniformidad <strong>de</strong> las abundancias relativas <strong>de</strong> las especies). La<br />
variación conjunta <strong>de</strong> ambos componentes <strong>de</strong>termina los cambios en la diversidad.<br />
• Dominancia: no todas las especies tienen la misma influencia sobre la comunidad; las dominantes<br />
pue<strong>de</strong>n ejercer un mayor control sobre la estructura comunitaria. La dominancia pue<strong>de</strong> estar dada<br />
por su abundancia, tamaño o actividad.<br />
• Abundancia relativa: las abundancias relativas entre las distintas especies también permiten<br />
<strong>de</strong>scribir a las comunida<strong>de</strong>s.<br />
• Estructura trófica: las relaciones alimentarias entre las especies <strong>de</strong> la comunidad <strong>de</strong>terminan el<br />
flujo <strong>de</strong> materia y energía.<br />
• Interacciones entre especies: una <strong>de</strong> las i<strong>de</strong>as implícitas en el concepto <strong>de</strong> comunidad es que<br />
existen <strong>de</strong>terminadas asociaciones <strong>de</strong> especies (i.e., que éstas aparecen juntas más a menudo<br />
que lo que uno esperaría por azar). Estas asociaciones pue<strong>de</strong>n <strong>de</strong>berse a las interacciones entre<br />
ellas, como en el caso <strong>de</strong> las mutualistas, o ser consecuencia <strong>de</strong> afinida<strong>de</strong>s <strong>de</strong> su biología (e.g.,<br />
requerimientos <strong>de</strong> hábitat similares).
90<br />
Ecología General<br />
De estos atributos comunitarios, uno <strong>de</strong> los más estudiados históricamente por los ecólogos ha<br />
sido la diversidad. El concepto <strong>de</strong> diversidad ha provisto un marco teórico importante para el<br />
<strong>de</strong>sarrollo <strong>de</strong> muchas especulaciones acerca <strong>de</strong> la estructura y el funcionamiento <strong>de</strong> las comunida<strong>de</strong>s<br />
(Magurran 1988). Al mismo tiempo, el interés por este tema se ha incrementado <strong>de</strong>bido a la creciente<br />
necesidad <strong>de</strong> compren<strong>de</strong>r los factores que gobiernan los patrones globales <strong>de</strong> biodiversidad (Sala et<br />
al. 2000).<br />
OBJETIVOS<br />
El objetivo general <strong>de</strong>l trabajo práctico es familiarizarse con el cálculo <strong>de</strong> algunos atributos<br />
comunitarios, con algunos aspectos <strong>de</strong>l muestreo <strong>de</strong> comunida<strong>de</strong>s y con la <strong>de</strong>terminación <strong>de</strong> la<br />
diversidad y sus componentes.<br />
Los objetivos específicos son:<br />
(1) Determinar el área mínima necesaria para estimar la riqueza específica <strong>de</strong> una comunidad.<br />
(2) Calcular la riqueza específica, la diversidad y la equitatividad <strong>de</strong> una comunidad.<br />
(3) Construir una curva rango-abundancia para una comunidad.<br />
(4) Evaluar el grado <strong>de</strong> similitud entre distintas comunida<strong>de</strong>s.<br />
(5) Estimar similitu<strong>de</strong>s y diferencias en los patrones <strong>de</strong> diversidad <strong>de</strong> varias comunida<strong>de</strong>s y explorar<br />
las posibles causas.<br />
DESARROLLO<br />
Se utilizará un “mapa” <strong>de</strong> una comunidad imaginaria. En el mapa se encuentran indicados (con<br />
distintas letras) los individuos <strong>de</strong> las distintas especies. El grillado facilita el muestreo, al mismo<br />
tiempo que cada cuadrado constituye una unidad muestreal mínima.<br />
1.- Determinación <strong>de</strong>l área mínima y <strong>de</strong> la riqueza específica <strong>de</strong> la comunidad<br />
Para evaluar el área mínima <strong>de</strong> muestreo necesaria para estimar la riqueza <strong>de</strong> la comunidad hay que<br />
realizar muestreos sucesivos <strong>de</strong> la riqueza en unida<strong>de</strong>s <strong>de</strong> tamaño cada vez mayor (véase figura 1).<br />
En cada paso se duplica la superficie <strong>de</strong> muestreo, extendiendo la unidad anterior (Matteucci y Colma<br />
1982). Las unida<strong>de</strong>s muestrales sucesivas son: un cuadrado, dos cuadrados, cuatro cuadrados (2x2),<br />
ocho cuadrados, 16 cuadrados (4x4), 32 cuadrados, 64 cuadrados (8x8), etc. Comenzar el muestreo<br />
en alguna <strong>de</strong> las esquinas <strong>de</strong>l bor<strong>de</strong> izquierdo <strong>de</strong>l mapa. Graficar el número <strong>de</strong> especies en función<br />
<strong>de</strong>l tamaño <strong>de</strong>l cuadrado. ¿Cuál es el tamaño que usted emplearía para <strong>de</strong>terminar la riqueza <strong>de</strong> la<br />
comunidad y por qué? Examine cómo se modifica su resultado si hubiera comenzado el muestreo en<br />
alguna <strong>de</strong> las esquinas <strong>de</strong>l bor<strong>de</strong> <strong>de</strong>recho <strong>de</strong>l mapa. ¿A qué lo atribuye? ¿Qué implicancias tiene esto<br />
para el muestreo <strong>de</strong> comunida<strong>de</strong>s?
Figura 1: Tamaños sucesivos <strong>de</strong> muestreador para la evaluación <strong>de</strong>l área mínima.<br />
57 58 59 60 61 62 63 64<br />
49 50 51 52 53 54 55 56<br />
41 42 43 44 45 46 47 48<br />
33 34 35 36 37 38 39 40<br />
25 26 27 28 29 30 31 32<br />
17 18 19 20 21 22 23 24<br />
9 10 11 12 13 14 15 16<br />
1 2 3 4 5 6 7 8<br />
2.- Estimación <strong>de</strong> la diversidad y <strong>de</strong> la equitatividad<br />
91<br />
Ecología General<br />
Para la diversidad, se utilizarán el índice <strong>de</strong> Shannon-Wiener (H) y el <strong>de</strong> Simpson (D). Utilizando un<br />
muestreador <strong>de</strong> 8 x 8, tomar 10 muestras al azar (elegir coor<strong>de</strong>nadas x e y al azar, y colocar el<br />
extremo superior izquierdo <strong>de</strong>l cuadrado muestral sobre la celda correspondiente; <strong>de</strong>scartar el punto<br />
si no cabe el muestreador entero). En cada uno <strong>de</strong> los 10 casos, contar el número <strong>de</strong> individuos <strong>de</strong><br />
cada una <strong>de</strong> las especies presentes, completando la tabla 1.<br />
Tabla 1. Número <strong>de</strong> individuos <strong>de</strong> cada especie por muestra (M), y suma <strong>de</strong> abundancias para cada<br />
especie.<br />
spp. A B C D E F G H I J K L M N<br />
M1<br />
M2<br />
M3<br />
M4<br />
M5<br />
M6<br />
M7<br />
M8<br />
M9<br />
M10<br />
Suma<br />
Sobre la base <strong>de</strong> los datos promedio correspondientes a las 10 muestras, calcular los siguientes<br />
índices, utilizando la tabla 2:<br />
(a) Índice <strong>de</strong> Shannon-Wiener. Basado en la teoría <strong>de</strong> la información, predice cuál es la probabilidad<br />
<strong>de</strong> que un individuo en una muestra sea <strong>de</strong> la misma especie que el <strong>de</strong> la muestra anterior.
S<br />
H = - ∑ (pi)(ln pi)<br />
i=1<br />
92<br />
Ecología General<br />
don<strong>de</strong> H = contenido <strong>de</strong> información <strong>de</strong> la muestra (diversidad); S = riqueza específica; pi =<br />
proporción <strong>de</strong> individuos <strong>de</strong> la especie i respecto al total <strong>de</strong> individuos. Varía entre un valor mínimo <strong>de</strong><br />
0 y un máximo que <strong>de</strong>pen<strong>de</strong> <strong>de</strong> la riqueza específica (véase más abajo).<br />
(b) Índice <strong>de</strong> Simpson. Basado en la teoría <strong>de</strong> probabilida<strong>de</strong>s. ¿Cuál es la probabilidad <strong>de</strong> que dos<br />
individuos tomados al azar pertenezcan a una misma especie? Si pi es la proporción <strong>de</strong> individuos <strong>de</strong><br />
la especie i respecto al total <strong>de</strong> individuos, entonces en una muestra <strong>de</strong> dos individuos, la<br />
2<br />
probabilidad <strong>de</strong> que sean los dos <strong>de</strong> la misma especie es pi * pi , o sea pi . Si se suman las<br />
probabilida<strong>de</strong>s para todas las especies presentes, se obtiene el índice <strong>de</strong> Simpson:<br />
S<br />
D = 1 - ∑ (pi) 2<br />
i=1<br />
Este índice otorga mayor peso a las especies abundantes que a las raras. Varía entre un valor<br />
mínimo <strong>de</strong> 0 (cuando todos los individuos pertenecen a la misma especie) y un máximo <strong>de</strong> (1 - 1/S)<br />
cuando los individuos se reparten equitativamente entre especies.<br />
(c) Equitatividad. El valor máximo <strong>de</strong> diversidad varía con el número <strong>de</strong> especies presentes. Usando<br />
el índice <strong>de</strong> Shannon-Wiener, para un S dado, el H será máximo cuando los individuos se distribuyan<br />
equitativamente entre las especies (i.e., cuando todos los pi sean iguales entre sí e iguales a 1/S).<br />
Reemplazando en la fórmula <strong>de</strong> H:<br />
S<br />
Hmáx = - ∑ (1/S)(ln 1/S) = - S(1/S)(ln(1/S)) = ln S<br />
i=1<br />
Equitatividad = H/Hmáx = H/ln S<br />
Tabla 2. Valores utilizados para el cálculo <strong>de</strong> la diversidad y la equitatividad.<br />
Suma pi ln pi (pi)(ln pi) (pi)<br />
spp.<br />
2<br />
A<br />
B<br />
C<br />
D<br />
E<br />
F<br />
G<br />
H<br />
I<br />
J<br />
K<br />
L<br />
M<br />
N<br />
∑
3.- Construcción <strong>de</strong> la curva <strong>de</strong> rango-abundancia<br />
93<br />
Ecología General<br />
Utilizando los datos <strong>de</strong> abundancia promedio <strong>de</strong> cada una <strong>de</strong> las especies (tabla 2), se <strong>de</strong>be<br />
or<strong>de</strong>narlas en or<strong>de</strong>n <strong>de</strong>creciente <strong>de</strong> abundancia, asignándole rango 1 a la más abundante, 2 a la<br />
segunda, y así sucesivamente. Volcar los datos <strong>de</strong>l número <strong>de</strong> individuos (abundancia) en función <strong>de</strong>l<br />
rango en la figura 2.<br />
Figura 2. Curva <strong>de</strong> rango-abundancia.<br />
Abundancia<br />
4.- Estimación <strong>de</strong> la similitud entre comunida<strong>de</strong>s<br />
Rango<br />
Existen distintos índices que permiten comparar comunida<strong>de</strong>s <strong>de</strong> a pares (Matteucci y Colma 1982,<br />
Crisci y López Armengol 1983). Estos índices pue<strong>de</strong>n ser cualitativos o cuantitativos. Los primeros se<br />
basan solo en la presencia o ausencia <strong>de</strong> las distintas especies en las dos comunida<strong>de</strong>s que se<br />
comparan, mientras que los cuantitativos utilizan información <strong>de</strong> la abundancia relativa <strong>de</strong> las<br />
especies.<br />
(a) Índice <strong>de</strong> Jaccard. Este índice cualitativo tiene en cuenta la relación entre el número <strong>de</strong> especies<br />
comunes a las dos comunida<strong>de</strong>s que se comparan y el total <strong>de</strong> las especies en ambas comunida<strong>de</strong>s.<br />
En la tabla 3 se muestra un esquema <strong>de</strong> los valores utilizados. Los valores oscilan entre 0 y 1. El<br />
índice es:<br />
Similitud = a / (a + b + c)<br />
(b) Índice <strong>de</strong> Sorensen. Este índice cualitativo conce<strong>de</strong> mayor significación a las presencias<br />
conjuntas. Los valores oscilan entre 0 y 1. El índice es:<br />
Similitud = (2a) / (2a + b + c)<br />
Tabla 3. Esquema <strong>de</strong> los valores utilizados para calcular la similitud cualitativa entre dos<br />
comunida<strong>de</strong>s A y B. En la tabla, a es el número <strong>de</strong> especies comunes a A y B, b es el número <strong>de</strong><br />
especies exclusivas <strong>de</strong> B, c es el número <strong>de</strong> especies exclusivas <strong>de</strong> A, y d es el número <strong>de</strong> especies<br />
ausentes en ambas muestras simultáneamente.<br />
Comunidad B<br />
Comunidad A<br />
Presente Ausente<br />
Presente a b<br />
Ausente c d
94<br />
Ecología General<br />
(c) Índice <strong>de</strong> Czekanowski. Este índice cuantitativo está basado en la menor abundancia <strong>de</strong> cada<br />
especie en las comunida<strong>de</strong>s que se comparan. Los valores oscilan entre 0 y 1. El índice es:<br />
S<br />
Similitud = ∑ mín (pi1, pi2)<br />
i=1<br />
don<strong>de</strong> pi1 = proporción <strong>de</strong> individuos <strong>de</strong> la especie i respecto al total <strong>de</strong> individuos en la comunidad 1;<br />
pi2 = proporción <strong>de</strong> individuos <strong>de</strong> la especie i respecto al total <strong>de</strong> individuos en la comunidad 2.<br />
Tabla 4. Abundancia <strong>de</strong> las especies <strong>de</strong> aves acuáticas en la Reserva Costanera Sur (C.S.) durante<br />
primavera, verano, otoño e invierno, y <strong>de</strong> las especies <strong>de</strong> aves acuáticas en humedales cercanos a<br />
Chascomús, Chasicó (al oeste <strong>de</strong> Bahía Blanca) y Mar Chiquita (en la costa, al norte <strong>de</strong> Mar <strong>de</strong>l<br />
Plata). Los valores correspon<strong>de</strong>n al número <strong>de</strong> individuos observados en censos estandarizados.<br />
C.S. C.S. C.S. C.S. Chascomús Chasicó Mar<br />
spp.<br />
primav verano otoño inv<br />
Chiquita<br />
Podiceps rolland 140 128 322 45 77<br />
Podiceps major 69 63 28 23<br />
Phalacrocorax olivaceus 29 19 43 12 36<br />
Egretta thula 25 3 1 7 12 5<br />
Bubulcus ibis 27 1 2 7 7 18 12<br />
Plegadis chihi 19 1 6 25 12<br />
Coscoroba coscoroba 25 14 21<br />
Cynus melancoryphus 16 32 5 7<br />
Anas georgica 10 2 11 23 12<br />
Anas flavirostris 12 3 32 11 13<br />
Anas platalea 9 2 12 2 17<br />
Anas cyanoptera 8 2 8 1<br />
Anas versicolor 9 3 25 15 33<br />
Dendrocygna bicolor 15 5 13 32<br />
Dendrocygna viduata 2 1 5 2 14<br />
Netta peposaca 14 8 31 46<br />
Heteronetta atricapilla 32 26 16 24<br />
Oxyura vittata 23 24<br />
Rallus sanguinolentus 1 5 1 1 6 2<br />
Fulica rufifrons 25 26 17 44 124 87<br />
Fulica leucoptera 125 145 98 65 155 107<br />
Fulica armillata 168 58 529 426 198 98<br />
Gallinula chloropus 18 10 6 35<br />
Jacana jacana 7 17 1 12 23<br />
Himantopus melanurus 6 20 1 9 15 45<br />
Vanellus chilensis 5 12 2 4 2 8<br />
Larus maculipennis 1 1 14 21 67<br />
Larus dominicanus 1 1 48<br />
Sterna tru<strong>de</strong>aui 3 1 19<br />
Podiceps occipitalis 3<br />
Anas sibilatrix 1<br />
Anas bahamensis 3<br />
Aramus guarauna 1<br />
Charadrius falklandicus 25<br />
Zonibyx mo<strong>de</strong>stus 12<br />
Gelochelidon nilotica 1<br />
Tringa flavipes 7 38<br />
Charadrius collaris 5 25
95<br />
Ecología General<br />
En la tabla 4 se presentan datos correspondientes a las abundancias <strong>de</strong> especies <strong>de</strong> aves acuáticas<br />
en la Reserva Costanera Sur (Buenos Aires) en distintas épocas <strong>de</strong>l ciclo anual y en otros humedales<br />
<strong>de</strong> la provincia <strong>de</strong> Buenos Aires. Usando dichos datos, calcule la similitud (con los tres índices<br />
<strong>de</strong>scriptos arriba) entre la comunidad <strong>de</strong> aves acuáticas <strong>de</strong> Costanera Sur y las otras tres<br />
comunida<strong>de</strong>s (para Costanera Sur utilice solamente los valores invernales, pues los datos <strong>de</strong> las<br />
comunida<strong>de</strong>s <strong>de</strong> la provincia <strong>de</strong> Buenos Aires fueron tomados en dicha estación). Comparar los<br />
resultados obtenidos con los distintos índices. ¿Cuáles podrían ser las causas <strong>de</strong>l grado <strong>de</strong> similitud<br />
observado entre las comunida<strong>de</strong>s.<br />
5.- Patrones <strong>de</strong> diversidad<br />
Los datos <strong>de</strong> Costanera Sur <strong>de</strong> la tabla 4 correspon<strong>de</strong>n a cuatro períodos <strong>de</strong>l ciclo anual, y fueron<br />
tomados en un año en el cual se produjo una inusual sequía. Los niveles <strong>de</strong> agua <strong>de</strong> las lagunas <strong>de</strong><br />
la reserva (que <strong>de</strong>pen<strong>de</strong>n <strong>de</strong>l régimen <strong>de</strong> precipitaciones y <strong>de</strong> las temperaturas) son máximos durante<br />
el invierno, y van disminuyendo hasta su mínimo durante el verano y el otoño subsiguiente. Durante el<br />
verano estudiado casi la totalidad <strong>de</strong> las lagunas estaban secas, <strong>de</strong>jando solo gran<strong>de</strong>s extensiones<br />
barrosas. Para el otoño, aunque el nivel <strong>de</strong>l agua era bajo, ya había una superficie anegada<br />
consi<strong>de</strong>rable. Calcular la diversidad (usando el índice <strong>de</strong> Shannon-Wiener), la riqueza <strong>de</strong> especies y<br />
la equitatividad correspondiente a los cuatro muestreos. Examine comparativamente los valores<br />
obtenidos. ¿De qué manera afectan la riqueza y la equitatividad a las estimaciones <strong>de</strong> diversidad?<br />
Construya la curva <strong>de</strong> rango-abundancia <strong>de</strong> cada muestreo. ¿Qué conclusiones pue<strong>de</strong> alcanzar<br />
comparando las cuatro curvas? ¿Cómo se relaciona la forma <strong>de</strong> las curvas con los valores <strong>de</strong><br />
diversidad y sus componentes? ¿Cuáles podrían ser las causas <strong>de</strong> los patrones observados?<br />
REFERENCIAS<br />
Crisci JV y López Armengol MF (1983) Introducción a la teoría y práctica <strong>de</strong> la taxonomía numérica.<br />
Monografía 26, Serie <strong>de</strong> Biología. OEA, Washington DC.<br />
MacArthur RH (1971) Patterns of terrestrial bird communities. Pp. 189-221 en: Avian biology. Farner<br />
DS y King JR (eds). Aca<strong>de</strong>mic Press, New York.<br />
Magurran AE (1988) Ecological diversity and its measurement. Princeton University Press, Princeton.<br />
Matteucci SD y Colma A (1982) Metodología para el estudio <strong>de</strong> la vegetación. Monografía 22, Serie<br />
<strong>de</strong> Biología. OEA, Washington DC.<br />
Sala OE, Chapin III FS, Armesto JJ, Berlow E, Bloomfield J, Dirzo R, Huber-Sanwald E, Huenneke LF,<br />
Jackson RB, Kinzig A, Leemans R, Lodge DM, Mooney HA, Oesterheld M, LeRoy Poff N, Sykes<br />
MT, Walker BT, Walker M & Wall DH (2000) Global biodiversity scenarios for the year 2100.<br />
Science 287:1770-1774.<br />
Whittaker RH (1975) Communities and ecosystems. Second edition. Macmillan, New York.<br />
Wiens JA (1989) The ecology of bird communities. Volume 1. Foundations and patterns. Cambridge<br />
University Press, Cambridge.
DEJAR ESTA HOJA EN BLANCO<br />
96<br />
Ecología General
97<br />
Ecología General<br />
arriba-izquierda]<br />
A B C A C G A J A A J A A<br />
B B E B B<br />
A B B G F E A<br />
A K E D I J B B I H A A<br />
G E B<br />
A B M A B A D A C L B<br />
F B E L B I A F<br />
E A N D B M B C B A A C<br />
B G C A<br />
C A D B F E A L B C H C<br />
A F D G C A A C C H<br />
F A C C A C G I<br />
G A A A J A B N J L C A<br />
A B A B C J N L C<br />
E L H I B D E H E C L D<br />
D D E I B C E A M<br />
C G G N A F J D<br />
I C J G C C<br />
I G J D J C A C C B<br />
F D J B J C G G<br />
F C M G M C<br />
D H H C C C B C B J D D C G<br />
I E J B C A<br />
H A B A H A B<br />
F G B G J B A<br />
A B B A A A A<br />
A L A G J G<br />
I J J D K A A<br />
A A<br />
D N A B A J B A G<br />
A G B A A G B<br />
F G J L A<br />
H B D H<br />
i
DEJAR ESTA HOJA EN BLANCO<br />
98<br />
Ecología General
99<br />
Ecología General<br />
[arriba-<strong>de</strong>recha]<br />
B A J A A<br />
D D B B B B<br />
B A B A A<br />
A G C<br />
G B A B A<br />
A A E B E D A A C<br />
A B A D C A I<br />
B A I A I A B A B A A<br />
A C B E A<br />
D A A E A B B B<br />
A B C J A C C C<br />
C A D K B B A C<br />
A C B N E E<br />
C B A C B A A L<br />
A D A B I A B B A B<br />
A C B A A A A C N<br />
D B D C A C<br />
C J J D G C A C C<br />
C C C C C C C J B J C C B C<br />
A A C C C B G C B<br />
C C A C M B<br />
C C E E A E E J D C D<br />
C C B B J<br />
D A K A J B A<br />
C C A B J B B<br />
A B B B A A A<br />
C C C C E A G B A<br />
C C J C K J<br />
C C F A G A A<br />
A C K B A J B B<br />
C A A G<br />
C C J C H B<br />
A C H D<br />
ii
DEJAR ESTA HOJA EN BLANCO<br />
100<br />
Ecología General
101<br />
Ecología General<br />
i<br />
G A B G A B<br />
A A B B A C A D J<br />
A A H F A B<br />
H A B B J L F<br />
L M A B D D L G I<br />
G G A B J A<br />
A B A B A F A<br />
B E D F J D A A<br />
H D E A I J E A E A<br />
I G<br />
D J E<br />
A A A M F E<br />
M D L F N B A B<br />
A B A B C G H A K F<br />
E L F<br />
A B B I A A I H A A<br />
A B D A I M F<br />
E A<br />
A B A F A F G B D<br />
D D B E B A E H<br />
E A B B N I A<br />
A B M E<br />
M I M N N F A B I<br />
A B D B I B K F E F<br />
M A B D C B I A E F<br />
F E E F<br />
A B B A A A A<br />
G G B A B J L B D<br />
B J A B A A<br />
A I L B E A A<br />
H E A I J E F E A D<br />
G G H B B I<br />
[abajo-izquierda]
DEJAR ESTA HOJA EN BLANCO<br />
102<br />
Ecología General
103<br />
Ecología General<br />
ii<br />
B J J A B<br />
J<br />
A B J A B<br />
A D J F<br />
B L A L<br />
A A A A B J A B B<br />
A A A<br />
A A A D A<br />
A I J E F E E E<br />
J<br />
A<br />
A M A M<br />
A A B<br />
A A A N F<br />
E E<br />
A A F G E<br />
I F F<br />
A A I E F K A<br />
M E F E<br />
F A A B B F E A<br />
A M M<br />
A B<br />
A A I I L F<br />
I A F<br />
A E E E<br />
A A A<br />
A A G A B J<br />
A A<br />
A A A A<br />
I E J E E E<br />
M<br />
[abajo-<strong>de</strong>recha]
DEJAR ESTA HOJA EN BLANCO<br />
104<br />
Ecología General
Trabajo práctico 11<br />
105<br />
Ecología General<br />
COMPARACIÓN DE LA COMPOSICIÓN DE LA COMUNIDAD DE INVERTEBRADOS CAMINADORES<br />
EN DOS AMBIENTES DIFERENTES DE CIUDAD UNIVERSITARIA.<br />
Las comunida<strong>de</strong>s biológicas <strong>de</strong>pen<strong>de</strong>n ampliamente <strong>de</strong> las interacciones entre organismos<br />
pequeños – mayormente invertebrados- plantas y microbios. Estudios <strong>de</strong> producción secundaria en<br />
insectos e invertebrados muestran que los insectos son gran<strong>de</strong>s productores <strong>de</strong> biomasa y<br />
conductores <strong>de</strong> energía a través <strong>de</strong> las comunida<strong>de</strong>s. Estudios llevados a campo en ambientes <strong>de</strong><br />
clima templado, indican que los insectos son los herbívoros más importantes, no sólo porque<br />
consumen una mayor proporción <strong>de</strong> producción primaria que las aves y roedores sino también porque<br />
son mucho más eficientes en convertirla en biomasa. Muchos invertebrados inconspicuos también<br />
<strong>de</strong>sarrollan papeles importantes para el funcionamiento <strong>de</strong>l ecosistema por su importancia en la<br />
aireación y drenaje <strong>de</strong>l suelo, <strong>de</strong>scomposición <strong>de</strong> materia orgánica, ciclo <strong>de</strong> nutrientes, distribución<br />
<strong>de</strong> semillas, herbivoría y como predadores y fuente <strong>de</strong> alimento para los predadores.<br />
La actividad <strong>de</strong>l hombre produce cambios en el medio ambiente que repercuten en forma directa<br />
o indirecta sobre diversos grupos vivientes, tanto animales como vegetales. Los efectos pue<strong>de</strong>n darse<br />
<strong>de</strong>s<strong>de</strong> el nivel individual, poblacional, comunitario, o afectar el flujo <strong>de</strong> materia y energía en el<br />
ecosistema. Muchos invertebrados están caracterizados por presentar distribuciones y movimientos<br />
restringidos y asociaciones con un único hábitat. La estructura <strong>de</strong> la vegetación refleja las condiciones<br />
ambientales y ejerce una fuerte influencia sobre la composición y funcionamiento <strong>de</strong> la comunidad.<br />
Por lo tanto, entre ambientes con distinto grado <strong>de</strong> intervención antrópica es <strong>de</strong> esperar encontrar<br />
diferencias cuali y/o cuantitativas en la composición entre las comunida<strong>de</strong>s comparadas.<br />
Objetivo: Comparar la diversidad <strong>de</strong> invertebrados caminadores en dos ambientes fisonómicamente<br />
diferentes <strong>de</strong>l campo <strong>de</strong> Ciudad Universitaria.<br />
Desarrollo:<br />
Se seleccionaron dos ambientes <strong>de</strong>ntro <strong>de</strong>l predio <strong>de</strong> la Ciudad Universitaria: un área<br />
enmalezada (campo experimental próxima al ingreso a Ciudad Universitaria) y un área don<strong>de</strong> las<br />
propieda<strong>de</strong>s fisico-quimicas <strong>de</strong>l suelo han sido modificadas al establecerse un camino <strong>de</strong> conchillas<br />
(zona <strong>de</strong> Mantenimiento <strong>de</strong> vehículos <strong>de</strong> la Ciudad Universitaria). En ambos ambientes los alumnos<br />
divididos en 6 grupos realizarán las siguientes activida<strong>de</strong>s:<br />
1. Estimación <strong>de</strong> la abundancia y composición <strong>de</strong> las comunida<strong>de</strong>s <strong>de</strong> artrópodos caminadores: En<br />
cada ambiente se instalarán líneas transectas <strong>de</strong> 2 trampas <strong>de</strong> caída para artrópodos<br />
caminadores por estación. Dentro <strong>de</strong> cada estación las trampas estarán separadas por 1 metro y<br />
entre cada estación la distancia será <strong>de</strong> 3 metros. Las trampas consisten en recipientes plásticos<br />
<strong>de</strong> ¼ l. que <strong>de</strong>ben ser enterrados <strong>de</strong> manera que la boca que<strong>de</strong> a ras <strong>de</strong>l suelo. La tapa se coloca<br />
unos cm por encima sostenida por alambre. En el interior se coloca alcohol al 80% para la<br />
conservación <strong>de</strong> los insectos. Se <strong>de</strong>jarán funcionar durante 5 días, al cabo <strong>de</strong> los cuales se<br />
retirarán y se llevarán al laboratorio para la estimación <strong>de</strong> abundancia, composición <strong>de</strong> especies,<br />
diversidad y equitatividad.<br />
2. Estimación <strong>de</strong> la cobertura <strong>de</strong> las especies vegetales: En cada ambiente se estimará la cobertura<br />
vegetal usando el método <strong>de</strong> interceptación lineal. Cada grupo consi<strong>de</strong>rará como línea transecta<br />
la línea <strong>de</strong> 1 metro que separa a las dos trampas <strong>de</strong> caída <strong>de</strong> invertebrados. En cada transecta se<br />
registrará la longitud interceptada por vegetación (herbácea latifoliada o herbácea graminiforme) y<br />
suelo <strong>de</strong>snudo. El registro se hará al ras <strong>de</strong>l suelo, a los 20 cm y aproximadamente al metro <strong>de</strong><br />
distancia <strong>de</strong>l suelo.<br />
Trabajo <strong>de</strong> laboratorio y análisis <strong>de</strong> datos:<br />
1. Estimación <strong>de</strong> la abundancia y composición <strong>de</strong> las comunida<strong>de</strong>s <strong>de</strong> artrópodos caminadores: Los<br />
invertebrados serán clasificados bajo lupa a nivel <strong>de</strong> clase y or<strong>de</strong>n (sólo para la clase Insecta).
<strong>Guía</strong> para clasificar los ejemplares más probables a ser capturados:<br />
106<br />
Ecología General<br />
Phylum Arthropoda<br />
• Clase Arachnida<br />
• Clase Crustacea, Sub- clase malacostraceos: Or<strong>de</strong>n Isopoda (ej. bicho bolita)<br />
Clase Insecta: Or<strong>de</strong>n Colémbolos, Or<strong>de</strong>n Odonatos (ej. libélulas); Or<strong>de</strong>n Ortópteros (ej. langosta,<br />
grillo, etc); Or<strong>de</strong>n Hemípteros (heterópteros: ej. chinche <strong>de</strong> campo); Or<strong>de</strong>n Coleópteros (ej.<br />
escarabajos); Or<strong>de</strong>n Lepidópteros (ej. mariposas o polillas, larvas o adultos); Or<strong>de</strong>n Dípteros<br />
(moscas); Or<strong>de</strong>n Himenóptera (hormigas, abejas, avispas).<br />
• Clase Chilopoda (ej. ciempiés)<br />
Invertebrados no artrópodos:<br />
• Phylum Mollusca<br />
• Phylum Nemato<strong>de</strong>s<br />
• Phylum Annelida, etc.<br />
En cada muestra se calculará el número <strong>de</strong> individuos <strong>de</strong> cada clase u or<strong>de</strong>n (para cada par <strong>de</strong><br />
muestras se suman las abundancias <strong>de</strong> los ejemplares <strong>de</strong>terminados). Se calculará el promedio para<br />
cada clase u or<strong>de</strong>n. Sobre esa base <strong>de</strong> los datos promedios correspondientes a las 30 muestras (total<br />
<strong>de</strong> todos los turnos) se calculará en cada ambiente la diversidad, mediante los índices <strong>de</strong> Shannon-<br />
Wiener y Simpson, y la equitatividad. Los índices <strong>de</strong> diversidad <strong>de</strong> Shannon-Wiener calculados para<br />
cada ambiente se compararán mediante el test <strong>de</strong> t <strong>de</strong> Hutcheson (Magurran 1988)<br />
Se graficará la curva <strong>de</strong> rangos <strong>de</strong> abundancia para ambas comunida<strong>de</strong>s.<br />
Se compararán cuali y cuantitativamente ambas comunida<strong>de</strong>s usando los índices <strong>de</strong> Jaccard,<br />
Sorensen y Czekanowski respectivamente.<br />
Test <strong>de</strong> t <strong>de</strong> Hutcheson (Magurran 1983). En cada tipo <strong>de</strong> ambiente calcular el H, así se obtiene<br />
el H’1 para el ambiente 1 y el H’2 para el ambiente 2. Luego calcular la varianza <strong>de</strong> cada una <strong>de</strong> esas H:<br />
Para el ambiente 1:<br />
Var H’1 = Σpi (ln pi) 2 2 +<br />
- (Σpi ln pi)<br />
N1<br />
(y aplica la misma formulita con los datos <strong>de</strong>l ambiente 2 para calcular Var H’2). Después se<br />
<strong>de</strong>termina el valor <strong>de</strong> t:<br />
/ H’1 - H’2 /<br />
(Var H’1 + Var H’2) 1/2<br />
t =<br />
GL (Grados <strong>de</strong> libertad) =<br />
S1 -1<br />
2N1 2<br />
(Var H’1 + Var H’2) 2<br />
(Var H’1) 2 /N1 + (Var H’2) 2 /N2<br />
En general, los grados <strong>de</strong> libertad son > 120, y el valor crítico correspondiente es 1.96.<br />
2. Estimación <strong>de</strong> la cobertura <strong>de</strong> las especies vegetales: La cobertura se estimará como el<br />
porcentaje <strong>de</strong> la longitud interceptada con respecto a la longitud total <strong>de</strong> la línea. La cobertura<br />
media y el <strong>de</strong>svío se estimará usando la información <strong>de</strong> todos los turnos.<br />
Discusión: Estará orientada en analizar la relación entre la composición <strong>de</strong> la comunidad <strong>de</strong>
107<br />
Ecología General<br />
invertebrados observada y esperada con la estructura y altura <strong>de</strong> la vegetación <strong>de</strong> cada ambiente.<br />
Materiales necesarios (aportados por la cátedra):<br />
• Trampas <strong>de</strong> caída para invertebrados caminadores (120)<br />
• Bolsas <strong>de</strong> nylon (120)<br />
• Alcohol al 80%<br />
• Sacabocados para muestras <strong>de</strong> suelo (1 por equipo ~ 18)<br />
• Guantes <strong>de</strong>scartables<br />
Materiales que <strong>de</strong>ben traer cada grupo <strong>de</strong> alumnos:<br />
• 2 frascos <strong>de</strong> vidrio con tapa (ej. mermelada o mayonesa)<br />
• Marcador in<strong>de</strong>leble<br />
• 1 cinta métrica<br />
• Repelente<br />
• Palita<br />
BIBLIOGRAFÍA<br />
Magurran A (1988) Ecological diversity and its measurement. Chapman and Hall, London.
Trabajo Práctico 12<br />
SEMINARIO INVASIONES BIOLOGICAS<br />
108<br />
Ecología General
Trabajo práctico 13<br />
PATRONES ESPACIALES Y ESCALAS<br />
109<br />
Ecología General<br />
OBJETIVO GENERAL<br />
Analizar la estructura espacial <strong>de</strong> la heterogeneidad y el efecto <strong>de</strong> la escala sobre la percepción <strong>de</strong> la<br />
heterogeneidad en un gradiente urbano-suburbano.<br />
INTRODUCCIÓN<br />
Los sistemas naturales son heterogéneos, y esta heterogeneidad pue<strong>de</strong> darse a distintas<br />
escalas espaciales. A una escala dada, la heterogeneidad pue<strong>de</strong> formar distintos patrones: si el<br />
ambiente va cambiando en forma continua y gradual a lo largo <strong>de</strong>l espacio hablamos <strong>de</strong> gradiente,<br />
cuando las porciones distintas se ubican en forma contigua, <strong>de</strong> mosaico ambiental, mientras que si<br />
po<strong>de</strong>mos distinguir porciones distintas <strong>de</strong>ntro <strong>de</strong> un ambiente predominante, se habla <strong>de</strong> una<br />
estructura <strong>de</strong> parche- matriz (Figura 1). Esta estructura espacial, es <strong>de</strong>cir, cómo se distribuye la<br />
heterogeneidad en el espacio, está <strong>de</strong>terminada principalmente por la heterogeneidad <strong>de</strong>l sustrato<br />
(por ejemplo, un gradiente <strong>de</strong> altura, o distintos tipos <strong>de</strong> suelo), por disturbios naturales (por ejemplo,<br />
inundaciones periódicas, incendios) o por las activida<strong>de</strong>s humanas (uso <strong>de</strong> la tierra, urbanizaciones).<br />
a b<br />
Figura 1. Esquemas <strong>de</strong> patrones <strong>de</strong> heterogeneidad: (a) parches <strong>de</strong>ntro <strong>de</strong> una matriz, (b) mosaico ambiental<br />
La ecología <strong>de</strong>l paisaje es la rama <strong>de</strong> la ecología que estudia cómo la estructura espacial afecta<br />
los procesos ecológicos y, a su vez, cómo estos procesos <strong>de</strong>terminan la estructura espacial. Un<br />
paisaje está formado por un conjunto <strong>de</strong> ecosistemas que se repiten en el espacio (a una escala <strong>de</strong><br />
entre 10 y 100 km <strong>de</strong> diámetro), y pue<strong>de</strong> ser <strong>de</strong>scripto por sus elementos: tipos, forma y tamaño <strong>de</strong><br />
parches, forma, longitud y ancho <strong>de</strong> corredores, y tipo <strong>de</strong> matriz. La matriz es el tipo <strong>de</strong> parche más<br />
frecuente, y pue<strong>de</strong> ser más o menos extensa, dispersa o agregada, y estar más o menos perforada.<br />
Por otro lado, un paisaje también se caracteriza por el arreglo espacial <strong>de</strong> los elementos (grado <strong>de</strong><br />
agregación) y el grado <strong>de</strong> diferencia entre los distintos tipos <strong>de</strong> parches (contraste), Figura 2.<br />
Un conjunto <strong>de</strong> paisajes forma una región, que es un área geográfica mayor <strong>de</strong>terminada por sus<br />
características geológicas, el macroclima y una actividad humana común, sus distintas partes están<br />
conectadas por transporte, comunicación y cultura. Tanto los paisajes como las regiones son escalas<br />
espaciales “humanas”, no correspon<strong>de</strong>n a niveles <strong>de</strong> organización biológica. Como hemos dicho<br />
antes, las características y los procesos que suce<strong>de</strong>n a escala <strong>de</strong> paisaje están <strong>de</strong>terminados por<br />
sus elementos, es <strong>de</strong>cir, por las características y procesos a una escala menor. Pero también están<br />
influidas por las características <strong>de</strong> la región don<strong>de</strong> están inmersos los paisajes. Es <strong>de</strong>cir, lo que<br />
suce<strong>de</strong> a una escala dada, está <strong>de</strong>terminada por los niveles superiores e inferiores.
Poca agregación Mucha agregación<br />
Figura 2. Componentes <strong>de</strong> la heterogeneidad: contraste y agregación<br />
110<br />
Poco<br />
contraste<br />
Mucho<br />
contraste<br />
Ecología General<br />
Según la escala en que estemos trabajando, vamos a po<strong>de</strong>r distinguir la heterogeneidad a<br />
distintos niveles, como cuando se cambia el aumento <strong>de</strong> un microscopio cambian los <strong>de</strong>talles <strong>de</strong><br />
algunas estructuras, o cuando un avión se va acercando a tierra va cambiando el nivel <strong>de</strong> <strong>de</strong>talle con<br />
que uno ve los objetos, pero <strong>de</strong>jamos <strong>de</strong> percibir otros niveles <strong>de</strong> heterogeneidad a mayor escala. Por<br />
otro lado, según el ambiente, la heterogeneidad pue<strong>de</strong> variar según la escala <strong>de</strong> observación <strong>de</strong>bido<br />
al tamaño <strong>de</strong> los parches. Si los parches son muy gran<strong>de</strong>s (grano grueso, Figura 3l trabajar en una<br />
escala pequeña (<strong>de</strong> <strong>de</strong>talle) no los percibimos <strong>de</strong>bido a que abarcamos un solo tipo <strong>de</strong> parche,<br />
mientras que si los parches son pequeños (grano fino, Figura 3) al trabajar a escalas gran<strong>de</strong>s no son<br />
distinguibles.<br />
Grano grueso Grano fino<br />
Figura 3. Grano <strong>de</strong> un paisaje <strong>de</strong> acuerdo al tamaño promedio <strong>de</strong> los parches
111<br />
Ecología General<br />
Es importante diferenciar entre la escala ecológica y la escala cartográfica. Esta última se refiere<br />
a la relación entre la distancia que hay entre dos puntos dados en su representación en un mapa<br />
respecto a su distancia real (Por ejemplo 1:50000, 1:250000). La escala ecológica se refiere a la<br />
dimensión espacial y/o temporal <strong>de</strong> un objeto o proceso. Un proceso <strong>de</strong> gran escala abarca un área<br />
gran<strong>de</strong> y/o dura un tiempo largo (por ejemplo, las glaciaciones ocuparon gran<strong>de</strong>s áreas y se<br />
produjeron durante largos períodos <strong>de</strong> tiempo). Por otro lado, un proceso a pequeña escala ocurre en<br />
dimensiones limitadas o en tiempos relativamente cortos (por ejemplo, perturbación producida por la<br />
caída <strong>de</strong> un árbol).<br />
La escala se distingue por su grano y su extensión. El grano es el tamaño <strong>de</strong> la mínima unidad<br />
distinguible (como el tamaño <strong>de</strong> los granos <strong>de</strong> arena o la rugosidad <strong>de</strong> una lija) y la extensión es el<br />
tamaño total abarcado (Figura 4). Si uno está parado en un punto, hasta dón<strong>de</strong> pue<strong>de</strong> abarcar con la<br />
vista sería la extensión y los <strong>de</strong>talles que pue<strong>de</strong> percibir son el grano. Entre ambos pue<strong>de</strong> haber un<br />
número variable <strong>de</strong> niveles intermedios. El grano y la extensión <strong>de</strong>pen<strong>de</strong>n <strong>de</strong> las características <strong>de</strong><br />
cada especie, principalmente su tamaño y movilidad. Las especies más gran<strong>de</strong>s y/o que recorren<br />
mayores distancias tien<strong>de</strong>n a tener una extensión mayor que las especies pequeñas y se<strong>de</strong>ntarias.<br />
Figura 4: Para un organismo dado, la extensión es el mayor tamaño al que pue<strong>de</strong> distinguir la heterogeneidad, el<br />
grano el menor tamaño. En la figura se distinguen 3 niveles, los parches más chicos representan el grano y el<br />
área más gran<strong>de</strong> representa la extensión.<br />
DESARROLLO<br />
Grano<br />
Extensión<br />
Área <strong>de</strong> estudio:<br />
En este trabajo práctico vamos a analizar el efecto <strong>de</strong> los cambios <strong>de</strong> escala en la percepción <strong>de</strong> la<br />
heterogeneidad en la región metropolitana <strong>de</strong> Buenos Aires y alre<strong>de</strong>dores, hasta la zona <strong>de</strong>l Río<br />
Luján- Otamendi, Partido <strong>de</strong> Campana. Se tratará <strong>de</strong> <strong>de</strong>terminar a cada escala los factores que<br />
<strong>de</strong>terminan la heterogeneidad, teniendo en cuenta que ésta es el resultado y la expresión <strong>de</strong><br />
procesos que actúan a distintas escalas y son <strong>de</strong> distinto origen, en particular en la región estudiada,<br />
la heterogeneidad observada resulta <strong>de</strong> la interacción entre procesos naturales y las<br />
transformaciones que introduce el hombre al hacer uso <strong>de</strong>l suelo.<br />
El área estudiada compren<strong>de</strong> un mosaico <strong>de</strong> comunida<strong>de</strong>s don<strong>de</strong> están representadas tres<br />
ecorregiones <strong>de</strong>terminadas principalmente por el clima y la topografía: la ecorregión Delta e Islas <strong>de</strong>l<br />
Paraná, representada por el bosque ribereño, la región Pampeana, representada por los pastizales<br />
<strong>de</strong> la zona alta, y la región <strong>de</strong>l Espinal, representada por los talares que se <strong>de</strong>sarrollan sobre<br />
albardones <strong>de</strong> conchilla en la barranca. La ecorregión Delta e Islas <strong>de</strong>l Paraná correspon<strong>de</strong> a los<br />
valles <strong>de</strong> inundación <strong>de</strong> los trayectos medios e inferiores <strong>de</strong> los río Paraná y Paraguay, e incluye al
112<br />
Ecología General<br />
Delta <strong>de</strong>l Paraná. Se trata <strong>de</strong> un paisaje <strong>de</strong> islas bajas e inundables. La ecorregión <strong>de</strong> la Pampa<br />
constituye el sistema <strong>de</strong> pra<strong>de</strong>ras más importante <strong>de</strong> la Argentina. Su relieve es relativamente plano y<br />
está expuesta a anegamientos permanentes o cíclicos. Las pra<strong>de</strong>ras estuvieron originalmente<br />
dominadas por gramíneas entre las que predominaron las <strong>de</strong>l género Poa, Stipa, Piptochaetiun y<br />
Aristida. Es la ecorregión que más transformación ha sufrido <strong>de</strong>bido a procesos <strong>de</strong> urbanización y a la<br />
implementación <strong>de</strong> la agricultura y gana<strong>de</strong>ría extensiva. El Espinal <strong>de</strong>s<strong>de</strong> el punto <strong>de</strong> vista arbóreo<br />
está caracterizado por la presencia <strong>de</strong>l género Prosopis (algarrobos, ñandubay y caldén). En la<br />
provincia <strong>de</strong> Buenos Aires los talares son los representantes típicos <strong>de</strong>l espinal.<br />
Pamp<br />
Pampa<br />
Delta<br />
AMBA<br />
Costa<br />
Figura 5. Unida<strong>de</strong>s <strong>de</strong>l paisaje en la región metropolitana <strong>de</strong> Buenos Aires<br />
Las características propias <strong>de</strong> cada ecorregión han sido modificadas por las activida<strong>de</strong>s<br />
humanas, dando lugar en el área a cinco gran<strong>de</strong>s unida<strong>de</strong>s <strong>de</strong> paisaje en la Región Metropolitana<br />
(Atlas Ambiental <strong>de</strong> Buenos Aires; Figura 5):<br />
• La Planicie Pampeana y la Franja Costera ocupada por la urbanización, a la que<br />
se <strong>de</strong>nomina Área Metropolitana <strong>de</strong> Buenos Aires (AMBA)<br />
• La Planicie Pampeana no ocupada por la urbanización, a la que se <strong>de</strong>nomina<br />
Pampa<br />
• La Franja Costera no ocupada por la urbanización, a la que se <strong>de</strong>nomina Costa<br />
• El Bajo Delta <strong>de</strong>l Río Paraná, que se <strong>de</strong>nomina Delta<br />
• El Estuario <strong>de</strong>l Río <strong>de</strong> la Plata que se <strong>de</strong>nomina Río<br />
La división entre el AMBA y la Pampa es dinámica y arbitraria, y entre ellas se presenta una<br />
interfase, comúnmente <strong>de</strong>nominada “periurbano”, en la cual se registran simultáneamente<br />
manifestaciones urbanas y rurales.<br />
Durante los últimos años, la Región Metropolitana <strong>de</strong> Buenos Aires ha experimentado una intensa<br />
transformación <strong>de</strong>l territorio y <strong>de</strong> los usos dominantes. Por ejemplo, en algunos sectores <strong>de</strong> los valles<br />
<strong>de</strong> inundación <strong>de</strong> los ríos Luján, Reconquista y Paraná <strong>de</strong> las Palmas pue<strong>de</strong> observarse que amplios<br />
sectores <strong>de</strong> la franja periurbana y <strong>de</strong> numerosos espacios intersticiales han pasado <strong>de</strong> un uso rural a<br />
otro urbano, especialmente resi<strong>de</strong>ncial, recreativo y comercial.<br />
Extendiéndose más hacia el norte <strong>de</strong> la Región Metropolitana, don<strong>de</strong> disminuye la urbanización,<br />
se pue<strong>de</strong> distinguir la transición <strong>de</strong> los pastizales en la zona alta hacia los talares <strong>de</strong> la barranca, los<br />
Río<br />
Costa
113<br />
Ecología General<br />
humedales en la zona baja hasta la selva en galería a lo largo <strong>de</strong> arroyos y <strong>de</strong>l río Paraná. En esta<br />
zona tenemos representadas comunida<strong>de</strong>s vegetales asociadas tanto a ambientes terrestres como a<br />
ambientes acuáticos. Po<strong>de</strong>mos encontrar diferentes comunida<strong>de</strong>s <strong>de</strong> herbáceas, bosques, ríos,<br />
arroyos y canales. A pequeña escala no es el clima el que <strong>de</strong>termina el <strong>de</strong>sarrollo <strong>de</strong> diferentes<br />
comunida<strong>de</strong>s vegetales, sino que los factores principales son la hidrología y el tipo <strong>de</strong> suelo, ambos<br />
asociados al relieve (presencia <strong>de</strong> barrancas, zonas <strong>de</strong> inundación) y microrelieve. Las comunida<strong>de</strong>s<br />
terrestres que se <strong>de</strong>sarrollan en cada zona <strong>de</strong>pen<strong>de</strong>n <strong>de</strong>l relieve y las características físico–químicas<br />
<strong>de</strong>l suelo (principalmente la salinidad y la humedad), nivel <strong>de</strong> la napa freática, frecuencia <strong>de</strong><br />
inundaciones y período durante el cual el suelo permanece inundado. Pue<strong>de</strong> observarse, entonces,<br />
un gradiente <strong>de</strong> las comunida<strong>de</strong>s vegetales asociado al relieve y a la hidrología (Figura 6).<br />
Figura 6. Esquema <strong>de</strong> localización <strong>de</strong> las diferentes comunida<strong>de</strong>s según su<br />
posición en la barranca <strong>de</strong> río<br />
Desarrollo <strong>de</strong>l Trabajo Práctico:<br />
El trabajo práctico constará <strong>de</strong> tres partes:<br />
A. Trabajo con el Google Earth: Describir la heterogeneidad espacial a distintas<br />
escalas cartográficas e i<strong>de</strong>ntificar los distintos elementos que se pue<strong>de</strong>n distinguir en<br />
cada caso.<br />
B. En terreno: Describir las características <strong>de</strong>l paisaje sobre distintos puntos <strong>de</strong>l<br />
gradiente <strong>de</strong>s<strong>de</strong> ambientes urbanos <strong>de</strong>l Partido <strong>de</strong> Vicente López cercanos a la<br />
General Paz hasta los suburbanos y rurales <strong>de</strong>l Partido <strong>de</strong> Campana.<br />
C. En terreno: Describir y caracterizar las comunida<strong>de</strong>s vegetales asociadas a las<br />
unida<strong>de</strong>s <strong>de</strong> paisaje Delta, Pampa y Espinal en la zona <strong>de</strong> Río Luján y Otamendi.<br />
PARTE A: Trabajo en laboratorio con GoogleEarth<br />
Google Earth es un programa informático que existe bajo este nombre <strong>de</strong>s<strong>de</strong> mayo 2005 y que<br />
permite visualizar el planeta entero a través <strong>de</strong> un mosaico <strong>de</strong> imágenes <strong>de</strong> satélite o fotografías<br />
aéreas. El programa permite visualizar la superficie terrestre <strong>de</strong>s<strong>de</strong> diferentes alturas, lo que implica<br />
diferentes escalas <strong>de</strong> la imagen que se obtiene.<br />
Activida<strong>de</strong>s:<br />
Parte baja<br />
<strong>de</strong>l humedal<br />
Talar Pastizal<br />
1) Descripción <strong>de</strong> la estructura <strong>de</strong>l paisaje a distintas escalas en tres puntos ubicados a distancia<br />
creciente <strong>de</strong> la ciudad <strong>de</strong> Buenos Aires:<br />
Se trabajará con los puntos geográficos 1, 3 y 5 especificados en la Tabla 1. Para ubicarlos en el<br />
mapa <strong>de</strong>be escribir las coor<strong>de</strong>nadas geográficas en el recuadro que dice “volar a” <strong>de</strong>l Google Earth<br />
(recordar que latitud Sur y longitud Oeste se representan con valores negativos). En cada punto se<br />
trabajará a tres escalas diferentes. Para ello se ubicará el punto en el centro <strong>de</strong> la pantalla y se<br />
buscará que la escala que muestra el Google Earth en el extremo inferior izquierdo, coincida con 4<br />
km, 1 km y 100 m.
114<br />
Ecología General<br />
Para cada punto y a cada escala especificar: Tipos <strong>de</strong> unida<strong>de</strong>s distintas que se distinguen,<br />
tamaño promedio <strong>de</strong> las unida<strong>de</strong>s, proporción <strong>de</strong>l área con cada unidad y contraste entre<br />
unida<strong>de</strong>s. Comparar los distintos puntos en cuanto a la heterogeneidad, y a cómo varía ésta según<br />
la escala. Los criterios a usar para i<strong>de</strong>ntificar las unida<strong>de</strong>s <strong>de</strong> paisaje, así como los distintos<br />
ambientes son: la forma, el color y la textura <strong>de</strong> la imagen.<br />
2) Descripción <strong>de</strong> la estructura <strong>de</strong>l paisaje a lo largo <strong>de</strong> una transecta que va <strong>de</strong>s<strong>de</strong> la ciudad <strong>de</strong><br />
Buenos Aires hacia el norte, abarcando unos 50 km. hasta la zona <strong>de</strong>l río Luján:<br />
Los cinco puntos especificados en la Tabla 1 se encuentran a lo largo <strong>de</strong> una transecta. Se<br />
ubicarán esos puntos en el mapa a partir <strong>de</strong> sus coor<strong>de</strong>nadas y se visualizará la transecta. Luego<br />
se trabajará con cada uno <strong>de</strong> ellos a una escala 1:1000, analizando una superficie <strong>de</strong><br />
aproximadamente 4 Km 2 (2 X 2 Km). Para cada punto se cuantificará sobre la imagen <strong>de</strong> Google<br />
Earth la composición relativa <strong>de</strong> distintos componentes <strong>de</strong>l paisaje, y los resultados serán<br />
volcados en la planilla adjunta (Tabla 2).<br />
Tabla 1. Localización <strong>de</strong> los puntos a observar durante el trabajo práctico (coor<strong>de</strong>nadas y manera <strong>de</strong><br />
localizarlos en el campo durante la salida)<br />
Punto 1<br />
Punto 2<br />
Punto 3<br />
Punto 4<br />
Punto 5<br />
Punto 6<br />
Localización en el campo (durante la salida)<br />
Tercer puente <strong>de</strong>spués <strong>de</strong> tomar Panamericana (Para mirar se<br />
consi<strong>de</strong>ra <strong>de</strong>s<strong>de</strong> la bajada durante un minuto y medio).<br />
Latitud -34.533176° Longitud -58.502494°<br />
Después <strong>de</strong> la bifurcación <strong>de</strong>l ramal tigre, justo pasando el<br />
primer puente.(Se empieza a mirar a partir <strong>de</strong> la bifurcación <strong>de</strong>l<br />
ramal tigre por un minuto y medio).<br />
Latitud -34.488707° Longitud -58.570718°<br />
El punto está en Panamericana y Ruta 197 (se empieza a mirar<br />
<strong>de</strong>s<strong>de</strong> el peaje, pasando por el cruce con 197 y medio minuto<br />
mas <strong>de</strong>spués <strong>de</strong> este último)<br />
Latitud -34.474717° Longitud -58.661775°<br />
El punto esta en la Av. Benavi<strong>de</strong>z (Ruta 27). (Se empieza a<br />
mirar a partir <strong>de</strong>l cruce <strong>de</strong>spués <strong>de</strong> la FORD durante un minuto)<br />
Latitud -34.420476° Longitud -58.717830°<br />
El punto esta pasando el primer puente peatonal <strong>de</strong>spués <strong>de</strong>l<br />
arroyo escobar. (Se empieza a mirar <strong>de</strong>s<strong>de</strong> el arroyo por un<br />
minuto)<br />
Latitud -34.367515° Longitud -58.774746°<br />
Cruzando el Río Luján, a partir <strong>de</strong>l cartel Reserva Los Cardales,<br />
aproximadamente km 60<br />
Latitud -34.295539° Longitud -58.884219°<br />
Indicaciones para<br />
observaciones (Parte B)<br />
Mirar durante un minuto y<br />
medio.<br />
Mirar durante un minuto y medio<br />
Mirar durante un minuto y medio<br />
Se empieza a mirar <strong>de</strong>s<strong>de</strong> el<br />
arroyo por un minuto y medio.<br />
Mirar durante un minuto y medio<br />
Mirar durante un minuto y medio
115<br />
Ecología General<br />
Tabla 2. Planilla para <strong>de</strong>scribir la estructura <strong>de</strong>l paisaje en los 5 puntos a lo largo <strong>de</strong> la transecta<br />
Buenos Aires–Río Luján.<br />
Punto 1 Punto 2 Punto 3 Punto 4 Punto 5<br />
Tipos <strong>de</strong> uso <strong>de</strong> la tierra<br />
a) % Resi<strong>de</strong>ncial (total)<br />
a1) % Viviendas multifamiliares<br />
(edificios y <strong>de</strong>partamentos)<br />
a2) % Viviendas unifamiliares<br />
(casas y ph)<br />
a3) % Barrios cerrados,<br />
countries<br />
b) % Industrial/Comercial<br />
(incluye: escuelas, cementerios, etc)<br />
c) % Espacios ver<strong>de</strong>s no naturales/<br />
parques (no pertenecientes a countries)<br />
d) % <strong>de</strong> terreno agrícola<br />
e) % <strong>de</strong> terreno natural o no agrícola<br />
(pasturas, pastizales naturales)<br />
f) % <strong>de</strong> rutas o autopistas<br />
Otros indicadores<br />
% total <strong>de</strong> la superficie construida o<br />
pavimentada (sin incluir calles o rutas)<br />
% total <strong>de</strong> la superficie con vegetación<br />
N° <strong>de</strong> cuadrados con rutas o caminos<br />
3) Exploración visual <strong>de</strong> las comunida<strong>de</strong>s vegetales que se visitarán durante la salida <strong>de</strong> campo.<br />
Solo para el punto 5, se intentará diferenciar –agrandando la escala– un conjunto <strong>de</strong><br />
comunida<strong>de</strong>s que, posteriormente, se observarán en campo. Para ello se usarán las<br />
coor<strong>de</strong>nadas provistas en la Tabla 3 para ubicar los distintos ambientes en el Google Earth. Se<br />
observarán las distintas texturas, estructuras y colores que puedan ser usados para<br />
reconocerlos. Se discutirá hasta qué punto es posible distinguir estas comunida<strong>de</strong>s <strong>de</strong>s<strong>de</strong> las<br />
imágenes aéreas y la importancia <strong>de</strong> la validación en el terreno.<br />
Tabla 3. Coor<strong>de</strong>nadas <strong>de</strong> los distintos ambientes que pue<strong>de</strong>n encontrarse en la zona <strong>de</strong>l Río Luján y<br />
Otamendi (Punto 5). El or<strong>de</strong>n <strong>de</strong> presentación se relaciona con una disminución <strong>de</strong> la permanencia <strong>de</strong> agua<br />
estancada y la frecuencia <strong>de</strong> inundación en el área.<br />
Ambientes Latitud Longitud<br />
Terrestres<br />
Pastizal Pampeano 34 0 ´13´ 26.56´´ S 58 0 54´10.79” O<br />
Campo aledaño Reserva 34 0 ´13´ 24.52´´ S 58 0 54´10.40” O<br />
Talar 34 0 ´13´ 19.82´´ S 58 0 53´54.64” O<br />
Humedales<br />
Pastizal salino 34 0 ´13´ 14.10´´ S 58 0 53´45.15” O<br />
Selva en Galería 34 0 ´10´ 54.29´´ S 58 0 52´14.59” O<br />
Albardón 34 0 ´10´ 28.81´´ S 58 0 52´10.89” O<br />
Pajonal 34 0 ´09´ 46.89´´ S 58 0 51´52.99” O<br />
Acuáticos<br />
Río Paraná <strong>de</strong> las Palmas 34 0 ´52´ 16.94´´ S 58 0 51´47.97” O<br />
Arroyo 34 0 ´09´ 31.67´´ S 58 0 52´16.94” O
PARTE B: Salida <strong>de</strong> Campo. Observación durante la recorrida<br />
1) Verificación <strong>de</strong> los datos obtenido en el Google Earth en la Parte A <strong>de</strong>l trabajo práctico.<br />
116<br />
Ecología General<br />
Se ubicará en el campo cada uno <strong>de</strong> los puntos observados en el Google Earth según las<br />
indicaciones <strong>de</strong> la Tabla 1. A partir <strong>de</strong> cada punto se observará durante un minuto y medio hacia<br />
cada lado <strong>de</strong> la ruta y se registrarán las estructuras observadas, completando la Tabla 4 con los<br />
porcentajes estimados.<br />
Tabla 4. Observaciones a campo para cada punto.<br />
Izquierda Derecha<br />
% total <strong>de</strong> la superficie construida<br />
o pavimentada<br />
Descripción:<br />
viviendas (unifamiliares o<br />
<strong>de</strong>partamentos.), oficinas o<br />
comercios, industria,<br />
construcciones rurales, otras<br />
% <strong>de</strong> la superficie con espacios<br />
ver<strong>de</strong>s<br />
Tipo <strong>de</strong> espacios ver<strong>de</strong>s:<br />
jardines particulares, arbolado<br />
urbano, parques <strong>de</strong> uso público,<br />
cultivos, campos <strong>de</strong> pastoreo,<br />
pastizales sin uso, otros<br />
Tipo <strong>de</strong> vegetación Árboles,<br />
arbustos, hierbas (gramíneas o<br />
dicotiledóneas)<br />
Principal uso <strong>de</strong> la tierra<br />
resi<strong>de</strong>ncial urbano, industrial,<br />
agrícola<br />
Principal (%):<br />
Secundario (%):<br />
Terciario (%):<br />
Dominante:<br />
Acompañante<br />
Principal (%):<br />
Secundario (%):<br />
Terciario (%):<br />
Dominante:<br />
Acompañante<br />
PARTE C. Salida <strong>de</strong> Campo. Observación <strong>de</strong> las comunida<strong>de</strong>s locales en la zona <strong>de</strong> Río Luján y<br />
Otamendi.<br />
1) Se observarán las principales comunida<strong>de</strong>s vegetales que se encuentran en la zona, prestando<br />
particular atención a los elementos que les permitan distinguirlas. Los datos recopilados se volcarán<br />
en las correspondientes planillas (Tablas 5 y 6).
117<br />
Ecología General<br />
Tabla 5. Planilla a completar con las comunida<strong>de</strong>s terrestres encontradas en la zona <strong>de</strong> Río Luján-<br />
Otamendi<br />
Ecorregión<br />
Altura (tomada con GPS)<br />
Vegetación dominante<br />
>75 %<br />
(árboles, arbustos,<br />
herbáceas graminiformes,<br />
herbáceas latifoliadas)<br />
Vegetación<br />
acompañante frecuente<br />
75-50 %<br />
(árboles, arbustos,<br />
herbáceas graminiformes,<br />
herbáceas latifoliadas)<br />
Vegetación<br />
acompañante ocasional<br />
2m, 1–2m,<br />
30cm–1m,
Ecorregión<br />
Altura (tomada con GPS)<br />
Vegetación dominante<br />
>75 %<br />
(árboles, arbustos,<br />
herbáceas graminiformes,<br />
herbáceas latifoliadas)<br />
Vegetación<br />
acompañante frecuente<br />
75-50 %<br />
(árboles, arbustos,<br />
herbáceas graminiformes,<br />
herbáceas latifoliadas)<br />
Vegetación<br />
acompañante ocasional<br />
2m, 1–2m,<br />
30cm–1m,
119<br />
Ecología General<br />
Tabla 6. Planilla a completar con las características <strong>de</strong> las comunida<strong>de</strong>s acuáticas encontradas en la<br />
zona <strong>de</strong> Río Luján- Otamendi<br />
1. Arroyo al lado <strong>de</strong>l camino isleño 2. Río Paraná<br />
Río / arroyo<br />
Vegetación orilla<br />
Vegetación arraigada<br />
Vegetación flotante<br />
% agua libre<br />
Ancho<br />
Velocidad <strong>de</strong> la corriente<br />
A modo <strong>de</strong> guía, se ofrece una breve <strong>de</strong>scripción <strong>de</strong> las principales comunida<strong>de</strong>s vegetales que<br />
se pue<strong>de</strong>n distinguir en el área <strong>de</strong> Otamendi:<br />
Comunida<strong>de</strong>s <strong>de</strong> humedal y pastizal:<br />
Zona <strong>de</strong> bajo, planicie aluvial y planicie <strong>de</strong> inundación. Por lo general, sus suelos se encuentran<br />
inundados permanentemente o semipermanentemente). Características <strong>de</strong>l bajo <strong>de</strong>l Rió <strong>de</strong> la Plata.<br />
Pajonal<br />
Ubicados don<strong>de</strong> la napa freática está cerca <strong>de</strong> la superficie o don<strong>de</strong> hay agua estancada.<br />
Básicamente hay dos tipos <strong>de</strong> pajonales: <strong>de</strong> Schoenoplectus californicus o <strong>de</strong> Scirpus americanus. En<br />
los lugares don<strong>de</strong> el agua permanece estancada durante la mayor parte <strong>de</strong>l año encontramos<br />
tamb8ién Typha latifolia (totora) y Juncus acutus (juncos).<br />
Espartillar<br />
La especie más abundante es la Spartina <strong>de</strong>nsiflora. Esta comunidad se ubica en suelos<br />
relativamente bajos, salobres, arcillosos y pantanosos (pero con menor cantidad <strong>de</strong> agua anegada<br />
que en los pajonales), y en los meses <strong>de</strong> verano el suelo está prácticamente seco.<br />
Junquillar <strong>de</strong> Juncus acutus (juncos)<br />
Ubicado en zonas húmedas suceptibles a inundaciones por lluvias o por <strong>de</strong>sbor<strong>de</strong> <strong>de</strong> los arroyuelos.<br />
Corta<strong>de</strong>ral <strong>de</strong> Corta<strong>de</strong>ria selloana<br />
Esta comunidad se encuentra paralela a los arroyuelos interiores (en los márgenes enlomados), las<br />
canaletas, las vías <strong>de</strong>l ferrocarril y el terraplén <strong>de</strong>l camino. También se la observa en zonas elevadas<br />
pero no libres <strong>de</strong> inundaciones periódicas y don<strong>de</strong> la napa freática está a muy poca profundidad.<br />
Pue<strong>de</strong> haber renovales <strong>de</strong> Erythrina crista-galli (ceibo) y las especies más frecuentes son: Baccharis<br />
saliscifolia (chilca) y Eryngium cabrerae (serrucheta).<br />
Pastizal <strong>de</strong> Dystichlis spicata<br />
Se ubica sobre las partes elevadas <strong>de</strong>l microrelieve, en zonas bajas sin anegamiento permanente o<br />
inundaciones periódicas. Al estar ubicado en suelos arcillosos salinos existe un predominio <strong>de</strong><br />
gramíneas halófitas <strong>de</strong> poca altura. La especie dominante es D. spicata, pero la acompañan Phyla<br />
canescens, Senecio geniculata y Chaetotropis chilensis. Se integran a este pastizal especies que<br />
revelan zona húmedas y salinas como Aster squamatus. También hay especies que se usan como<br />
forrajeras Paspalum dilatatum (pasto miel) y Trifolium repens (trébol). En las zonas más elevadas hay<br />
Cynodon dactylon (pata <strong>de</strong> perdiz).
120<br />
Ecología General<br />
Pastizal <strong>de</strong> Sporobolus pyramidatus y Xanthium cavanillesii<br />
En los albardones <strong>de</strong>l río Luján existe esta comunidad en la cual existen dos especies codominantes:<br />
Sporobolus pyramidatus y Xanthium cavanillesii (abrojo).<br />
Comunida<strong>de</strong>s <strong>de</strong> bosque:<br />
Se encuentran en las partes altas y en la media loma <strong>de</strong>l perfil. Po<strong>de</strong>mos distinguir entre bosques<br />
xéricos (el talar) y los bosques ribereños, asociados a cursos <strong>de</strong> agua o zonas con buena<br />
disponibilidad <strong>de</strong> agua (ceibal y selva en galería).<br />
Bosque <strong>de</strong> Celtis tala (tala)<br />
Los talares son bosques xéricos <strong>de</strong> tala (Celtis tala) ubicados en la barranca (entre la terraza alta y<br />
baja, con pendiente pronunciada. Se <strong>de</strong>sarrollan sobre suelos compactos con tosca, médanos<br />
muertos y <strong>de</strong>pósitos <strong>de</strong> conchilla. Es una formación típica <strong>de</strong>l espinal <strong>de</strong>l norte bonaerense que ha<br />
sido intensamente utilizada para la extracción <strong>de</strong> ma<strong>de</strong>ra y leña y actualmente subsiste como<br />
manchones cada vez más pequeños.<br />
Ceibal (Erythrrina crista-galli)<br />
En los suelos pantanosos se <strong>de</strong>sarrolla el ceibal, cerca <strong>de</strong> la orilla <strong>de</strong>l río Paraná <strong>de</strong> Las Palmas y a<br />
continuación <strong>de</strong>l albardón <strong>de</strong>l mismo río. Es un bosque abierto que pue<strong>de</strong> tener herbáceas<br />
dominantes <strong>de</strong> los pajonales y algunos elementos <strong>de</strong> la selva en galería por ejemplo enreda<strong>de</strong>ras.<br />
Selva en galería<br />
Esta comunidad también recibe el nombre <strong>de</strong> monte blanco (Kandus et al. 2006) o bosque <strong>de</strong> ribera.<br />
Se <strong>de</strong>sarrolla principalmente en los albardones <strong>de</strong> las islas y ríos (zonas altas <strong>de</strong>l terreno). Su<br />
formación se ve favorecida por una oferta hídrica uniforme a lo largo <strong>de</strong>l año sin llegar al<br />
anegamiento. Se encuentran representadas especies <strong>de</strong> la ecorregión paranaense, entre las que se<br />
<strong>de</strong>stacan numerosas herbáceas, enreda<strong>de</strong>ras, epifitas, arbustos y árboles. Se pue<strong>de</strong> observar a la<br />
palmera pindó, sauces, ceibos y bromeliáceas.<br />
Comunida<strong>de</strong>s acuáticas:<br />
Se encuentran en terrenos permanentemente inundados y con aguas estancadas. Representada<br />
principalmente por Azolla filiculoi<strong>de</strong>s y Pistia stratiotes, aunque hay especies acompañantes como ser<br />
Lemna minima (lenteja <strong>de</strong> agua) y Salvinia rotundifolia (helechito <strong>de</strong> agua). El <strong>de</strong>sarrollo <strong>de</strong> esta<br />
comunidad varía si se trata <strong>de</strong> un cuerpo <strong>de</strong> agua lótico (río) o léntico (lagos, arroyos <strong>de</strong> curso lento).<br />
Pudiendo, en el caso <strong>de</strong> los ríos, estar representada principalmente por juncos y totoras.<br />
Plantas invasoras<br />
Algunas comunida<strong>de</strong>s están siendo invadidas por plantas invasoras (plantas exóticas naturalizadas<br />
que se dispersan naturalmente y producen gran<strong>de</strong>s transformaciones en los ecosistemas) o por<br />
plantas nativas que no correspon<strong>de</strong>n a esa comunidad. Estas introducciones se <strong>de</strong>ben principalmente<br />
a la construcción y mantenimiento <strong>de</strong> caminos, pero también <strong>de</strong>bido a los cambios en el uso <strong>de</strong> la<br />
tierra que ocurrieron en las últimas décadas. Por ejemplo, en la Reserva Natural Otamendi, la<br />
eliminación <strong>de</strong> la gana<strong>de</strong>ría condujo a la invasión <strong>de</strong> Dipsacus fullanum (la carda). Esta planta invadió<br />
la barranca y, en la última década, las comunida<strong>de</strong>s <strong>de</strong>l bajo, incluyendo las comunida<strong>de</strong>s <strong>de</strong> suelos<br />
salino. Otro ejemplo son el ligustro, la ligustrina y las acacias, que han invadido el bosque <strong>de</strong> tala.<br />
2) Trabajo <strong>de</strong> laboratorio:<br />
I. Describir la estructura <strong>de</strong>l paisaje a lo largo la transecta (Buenos Aires–Río Luján). Graficar los<br />
valores <strong>de</strong> las variables medidas para cada sitio y analizar su variación a lo largo <strong>de</strong>l gradiente.<br />
Comparar lo obtenido a campo con lo que se observó usando las imágenes <strong>de</strong> Google Earth.<br />
II. Para las comunida<strong>de</strong>s <strong>de</strong> cada tipo <strong>de</strong> ambiente, comparar las observaciones realizadas a campo<br />
con lo observado en el Google Earth. ¿Qué atributos a campo le parece que <strong>de</strong>terminan la textura y<br />
el color observados en la imagen <strong>de</strong> Google Earth?
121<br />
Ecología General<br />
III. Describir y caracterizar las comunida<strong>de</strong>s observadas en cada ambiente en el campo teniendo en<br />
cuenta las variables bióticas y abióticas que se registraron a campo. ¿Cómo se relacionan las<br />
variables bióticas y abióticas en cada comunidad?<br />
IV. En base a sus datos y a la bibliografía grafique un perfil topográfico <strong>de</strong>l área recorrida y ubique las<br />
comunida<strong>de</strong>s i<strong>de</strong>ntificadas. Analice dicho perfil en función <strong>de</strong> las variables ambientales.<br />
BIBLIOGRAFÍA<br />
Atlas Ambiental <strong>de</strong> Buenos Aires. http://www.atlas<strong>de</strong>buenosaires.gov.ar/aaba<br />
Begon, M., Harper, J.L.& Towsend, C.R. 1987. Ecología: Individuos, poblaciones y comunida<strong>de</strong>s. De.<br />
Omega, Barcelona.<br />
Bonfilds, C. G. 1962. Los suelos <strong>de</strong>l Delta <strong>de</strong>l río Paraná. Factores generadores, clasificación y uso.<br />
INTA (RIA). Num. 16 (3) 370 pp. Buenos Aires.<br />
Bonfilds, C. G. 1962. Los suelos <strong>de</strong>l Delta <strong>de</strong>l río Paraná. Factores generadores, clasificación y uso.<br />
INTA (RIA). Num. 16 (3) 370 pp. Buenos Aires.<br />
Brown A., Martínez Ortíz U., Acerbi M., Corcuera J., 2005. La situación ambiental argentina 2005.<br />
Fundación vida silvestre argentina.<br />
Chichizola S., 1993. Las comunida<strong>de</strong>s vegetales <strong>de</strong> la Reserva Natural Estricta Otamendi y sus<br />
relaciones con el ambiente. Parodiana 8 (2): 227-263.<br />
Forman RTT, (1999) Landscape ecology, the growing foundation in landuse planning and naturalresource<br />
management. In: Kovar P (Ed.) Nature and culture in landscape<br />
Iriondo MH y Scotta E. 1978. The evolution of the Paraná River Delta. Proceedings of the International<br />
Symposium on coastal Evolution in the Quaternary: 405-418. INQUA,San Pablo, Brasil<br />
Kandus P., Malvárez A.I. y N. Madanes. 2003. Estudio <strong>de</strong> las comunida<strong>de</strong>s <strong>de</strong> Plantas Naturales <strong>de</strong><br />
las Islas <strong>de</strong>l Bajo Delta <strong>de</strong>l Río Paraná. (Argentina). Darwiniana 41 (1-4): 1-16. Nacional.ISSN 0011-<br />
6793<br />
Kandus P., Quintana R D., Bó R., 2006. Patrones <strong>de</strong> paisaje y biodiversidad <strong>de</strong>l bajo <strong>de</strong>lta <strong>de</strong>l río<br />
Paraná. http://www.ambiente.gov.ar/<strong>de</strong>fault.asp?IdArticulo=5505<br />
Krebs, Charles, J. 1986. Ecología. Análisis experimental <strong>de</strong> la distribución y abundancia. Ed.<br />
Pirámi<strong>de</strong>. Madrid.<br />
Madanes, N. 2008. Humedales <strong>de</strong> la Reserva Natural Otamendi. En <strong>Guía</strong> <strong>de</strong> la Flora <strong>de</strong> la Reserva<br />
Natural Otamendi. Editora Liliana Goveto. Publicación <strong>de</strong> la Administración <strong>de</strong> Parques Nacionales.<br />
En prensa<br />
Malvárez, A.I y R.F. Bó. 2002. Cambios ecológicos en el Delta Medio <strong>de</strong>l Río Paraná <strong>de</strong>bidos al<br />
evento <strong>de</strong> El Niño 1982-1983”. Publicación especial <strong>de</strong>l Taller “El Niño, sus impactos en el Plata y<br />
en la Región Pampeana. J.A. Schnack (Ed.). La Plata, Buenos Aires.<br />
Marchetti B, Ruiz L., Madanes N., Sartori G. y P. Cichero. 1988. Relevamiento <strong>de</strong>l medio natural y una<br />
propuesta <strong>de</strong> plan <strong>de</strong> manejo para la futura Area Natural Protegida " Ing. Rómulo Otamendi".<br />
Informe Interno Administración Parques Nacionales: 1-53. Biblioteca Administración <strong>de</strong> Parques<br />
Nacionales<br />
Matteucci, S. y Colma A. 1982. Metodología para el estudio <strong>de</strong> la vegetación. O.E.A. Serie <strong>de</strong><br />
Biología.<br />
Matteucci, Silvia D.; Jorge Morello; Gustavo D. Buzai; Claudia A. Baxendale; Mariana Silva; Nora<br />
Mendoza; Walter Pengue y Andrea Rodríguez. 2006. Crecimiento urbano y sus consecuencias<br />
sobre el entorno rural. Orientación Gráfica Editora. Buenos Aires. (350 páginas).ISBN 978-987-<br />
9260-45-6


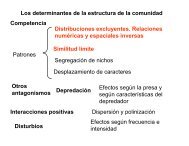





![¿Cómo escribir un trabajo científico? [PDF]](https://img.yumpu.com/14415288/1/184x260/como-escribir-un-trabajo-cientifico-pdf.jpg?quality=85)



