10.08 Echinodermi - Scienze della terra
10.08 Echinodermi - Scienze della terra
10.08 Echinodermi - Scienze della terra

You also want an ePaper? Increase the reach of your titles
YUMPU automatically turns print PDFs into web optimized ePapers that Google loves.
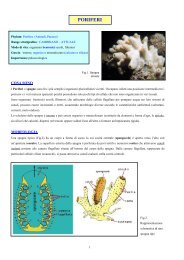
Phylum ECHINODERMATA<br />
Questo phylum, i cui primi rappresentanti, secondo alcuni autori, possono<br />
risalire al Vendiano, è caratterizzato da alcune peculiarità funzionali non presenti<br />
in altri organismi.<br />
Le più esclusiva è forse la presenza di un si isttema acqui ifferro, che funziona<br />
per pressione osmotica. Questo fatto li rende animali stenoalini e li obbliga<br />
a vivere solo in ambiente marino a normale salinità, non potendo sopportarne<br />
forti variazioni, pena la rottura del sistema acquifero, quindi la morte.<br />
Il gruppo comprende una grande varietà di forme sia fossili che attuali. Tra<br />
queste, ci sono famigliari le stelle di mare (Stelleroidea), le ofiure (Ofiuroidea),<br />
cetrioli di mare (Holoturoidea) ed i gigli di mare (Crinoidea).<br />
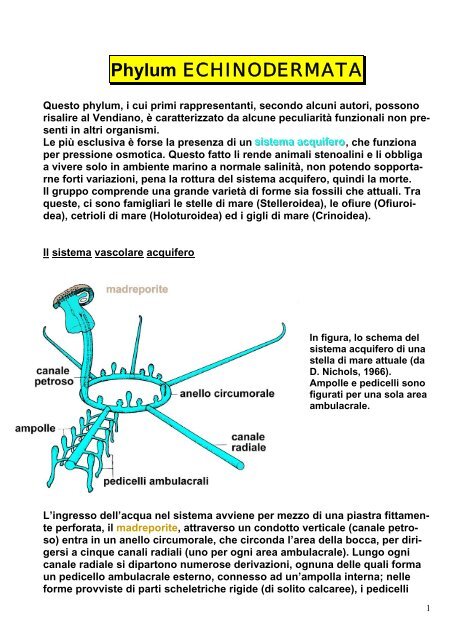
Il sistema vascolare acquifero<br />
In figura, lo schema del<br />
sistema acquifero di una<br />
stella di mare attuale (da<br />
D. Nichols, 1966).<br />
Ampolle e pedicelli sono<br />
figurati per una sola area<br />
ambulacrale.<br />
L’ingresso dell’acqua nel sistema avviene per mezzo di una piastra fittamente<br />
perforata, il madreporite, attraverso un condotto verticale (canale petroso)<br />
entra in un anello circumorale, che circonda l’area <strong>della</strong> bocca, per dirigersi<br />
a cinque canali radiali (uno per ogni area ambulacrale). Lungo ogni<br />
canale radiale si dipartono numerose derivazioni, ognuna delle quali forma<br />
un pedicello ambulacrale esterno, connesso ad un’ampolla interna; nelle<br />
forme provviste di parti scheletriche rigide (di solito calcaree), i pedicelli<br />
1
fuoriescono da aperture semplici o doppie che prendono il nome di “pori<br />
ambulacrali”. Come suggerito dalla terminologia impiegata, il sistema<br />
acquifero svolge una doppia funzione: 1) deambulatoria e 2) di cattura del<br />
particellato alimentare per il sostentamento dell’animale. È intuibile che il<br />
rigonfiamento delle ampolle, provoca l’inturgidimento dei pedicelli e la loro<br />
fuoriuscita dalla parte scheletrica rigida (nelle forme che la possiedono).<br />
In figura, sezioni longitudinali<br />
schematiche di nove rappresentanti<br />
delle numerose classi<br />
viventi e fossili che compongono<br />
il phylum Echinodermata.<br />
Tutti possiedono un si isttema<br />
acqui ifferro con madreporite (m),<br />
ampolle e pedicelli ambulacrali<br />
(p), un apparratto di igerrentte con<br />
bocca (b) ed ano (a), braccia<br />
(br) o brachiole (bch); alcuni<br />
presentano denti nella lanterna<br />
(d), radioli (sp), spiracoli (spr) e<br />
idrospira (i) nei Blastoidi; pori<br />
tecali (pt) nei Cistoidi.<br />
2
Microstruttura dello scheletro<br />
Lo scheletro dei ricci di mare si forma dal mesoderma e durante la vita<br />
è inglobato nei tessuti molli. É composto di calcite alto magnesiaca<br />
o calcite (CaCO3) incorporante il 15% MgCO3. Ciascun frammento o<br />
parte si presenta otticamente come un cristallo singolo. Ad alto ingrandimento<br />
(figura in alto), la parte scheletrica appare come un reticolato<br />
tridimensionale che è chiamato stereoma. Le dimensioni dei<br />
pori e lo spessore dei filamenti formanti lo stereoma varia in funzione<br />
del tessuto molle annesso a questa parte dello scheletro.<br />
Nell’immagine al SEM sopra riportata, si può scorgere il limite tra due<br />
tipi di stereoma, uno dove si attacca un muscolo (in alto a destra) e<br />
l’altro connesso ad un normale epitelio (pelle – in basso a sinistra).<br />
In vita lo spazio tra i pori è riempito di tessuto connettivo che è denominato<br />
stroma. Il disegno scheletrico degli echinidi sembra ideato<br />
per resistere alle tensioni che possono propagarsi fra le placche.<br />
Vedi: Smith, A. B. 1981. The stereom microstructure of the echinoid<br />
test. Special Papers in Palaeontology, 25, 1-85.<br />
Echi. 0a
Classificazione schematica<br />
Phylum Echinodermata<br />
Subphylum Pelmatozoa = forme bentoniche sessili (Cambriano - Attuale)<br />
†Classe Eocrinoidea, crinoidi primitivi (Cambriano m. - Ordoviciano medio)<br />
†Classe Paracrinoidea, crinoidi primitivi (Ordoviciano medio)<br />
†Classe Carpoidea, forme lateralmente compresse (Cambriano medio - Dev. inf.)<br />
†Classe Edrioasteroidea, forme discoidali (Cambriano inf. - Carbonifero inf.)<br />
†Classe Cistoidea, teca irregolare portante pori (Ordoviciano medio - Dev. sup.)<br />
Echi. 01<br />
†Classe Blastoidea, teca simmetrica a bocciolo (Ordoviciano medio - Permiano sup.)<br />
Classe Crinoidea, "gigli di mare" (Ordoviciano inf. - Attuale)<br />
†Sottoclasse Inadunata, braccia libere sotto le placchette radiali (Ord. inf. - Triassico)<br />
†Sottoclasse Flexibilia, placchette del calice mobili (Ord. medio - Permiano sup.)<br />
†Sottoclasse Camerata, placchette radiali rigide nel calice (Ord. - Permiano medio)<br />
Sottoclasse Articulata, la maggior parte dei crinoidi peduncolati (Triassico inf. - Attuale)<br />
Subphylum Eleuterozoa = forme mobili libere (Cambriano medio - Attuale)<br />
Classe Holoturoidea, "cetriolo di mare" (Cambriano medio - Carbonifero - Attuale)<br />
Classe Stelleroidea, "stelle di mare" (Ordoviciano - Attuale)<br />
Sottoclasse Ofiuroidea, "stelle di mare setolose" (Ordoviciano - Attuale)<br />
Classe Echinoidea, eleuterozoi non stellati (Ordoviciano - Attuale)<br />
Sottoclasse Regolari, ricci di mare regolari (Ordoviciano - Attuale)<br />
Sottoclasse Irregolari, ricci di mare a cuore e "dollari delle sabbie" (Giurassico - Attuale)<br />
†Classe Botriocidaroidea, echinidi primitivi simili agli eleuterozoi (Ordoviciano)<br />
† forme estinte<br />
Nota: per quello che riguarda le illustrazioni e la sistematica di innumerevoli generi di<br />
Echinidi si vada alla pagina web: http://www.nhm.ac.uk/researchcuration/projects/echinoid-directory/taxa/search.jsp?search,<br />
forse la pagina più<br />
completa sulla tassonomia degli Echinidi, curata dal Museo di <strong>Scienze</strong> Naturali di Londra.
Subphylum BLASTOZOA<br />
Cisto. 1<br />
Questo taxon d’alto livello sistematico comprende gruppi molto eterogenei di pelmatozoi<br />
paleozoici estinti. Fra questi si annoverano importante classi quali Diploporita, Rombifera<br />
(Cistoididea) e Blastoidea. Tutti possiedono un sistema acquifero costituito da aperture (pori<br />
e fessure) che attraversano le placche <strong>della</strong> teca.<br />
Classe Cistoidi<br />
Sulla base dei vari tipi di aperture si distinguono due principali gruppi funzionali: i Diploporita<br />
con pori esotecali ed i Rombifera con aperture a fessura endotecali. Forme come<br />
Haplosphaeronis (diploporide), principalmente noto nell’Ordoviciano dell’Europa settentrionale,<br />
possiede una teca ovoidale di c.a. 3cm con base piatta appoggiata<br />
al substrato. La teca è formata da due serie di placche<br />
poligonali: la serie inferiore di 7 placche laterali una delle<br />
quali con i classici diplopori, e quella superiore, sempre di 7<br />
placche periorali, alla cui sommità si apre una bocca a fessura<br />
dalla quale si dipartono 5 solchi alimentari radiali al termine<br />
dei quali si inseriscono le brachiole. Nella faccia superiore<br />
si apre anche il madreporite. L’ano, di solito sorretto da<br />
una piccola piramide anale, si trova in varie posizioni su una<br />
delle placche <strong>della</strong> teca. Altri cistoidi dell’Ordoviciano, come il<br />
diploporite Arictocystites ed il rombiferide Echinospaerites hanno una teca composta da<br />
numerose placche irregolari; vedi figura.<br />
Per quanto concerne il gruppo dei Rombifera, sono tutti caratterizzati dall’avere aperture endotecali<br />
disposte secondo una geometria rombica di struttura molto particolare e varia. Si<br />
possono infatti distinguere (secondo Paul, 1972) diversi tipi di Pettinirombi fra placche<br />
adiacenti: congiunti, disgiunti e criptorombi (o rombi nascosti), vedi figure.<br />
Echinosphaerites (rombiferide)<br />
con piramide anale<br />
e numerosi criptorombi.<br />
1
Più in dettaglio: i pettinirombi si possono aprire verso l’esterno per tutta la loro lunghezza e<br />
sono detti congiunti, oppure possono essere coperti per un tratto più o meno esteso in prossimità<br />
delle suture tra le placche, e sono detti disgiunti. In alcuni casi possono sporgere<br />
all’esterno solo le loro estremità che formano allineamenti romboidali convergenti verso il<br />
centro delle placche e prendono il nome di criptorombi, come nel gen. Echinosphaerites.<br />
Esistono infine alcuni cistoidi i cui tubi sono internamente ricoperti da un sottile tetto<br />
calcareo chiamato epistereom, e quindi le aperture verso l’esterno sono quelle delle estremità<br />
dei tubi: è il caso dei criptorombi, o rombi nascosti, che si possono vedere solo dopo<br />
una leggera abrasione <strong>della</strong> superficie tecale (evidenziati a volte dall’ erosione meteorica).<br />
Per quello che riguarda la disposizione delle placche tecali, i Rombiferida possiedono molte<br />
placche irregolarmente disposte. Pochi si distinguono per una disposizione regolare e fra<br />
questi possiamo citare Cheirocrinus (Ord. m. e sup., USA), in cui si distinguono 4 circoli di<br />
placche: procedendo dall’alto al basso abbiamo radiali, laterali, infralaterali e basali. Se i circoli<br />
sono solo 3, come in Cystoblastus (Ord. m., Russia), quelle che mancano sono le basali.<br />
Cheirocrinus<br />
Cystoblastus<br />
Un rappresentante specializzato del gruppo è Pleurocystites dell’Ordoviciano medio che possiede<br />
solo 3 pettinirombi, in posizione ben definita sul lato anale, e due robuste brachiole.<br />
Pleurocystites<br />
Glyptocystella<br />
2
Non possiamo concludere la trattazione di questo gruppo di fossili senza accennare ai ritrovamenti<br />
Italiani di Rombifera, (Caryocrinitidae e Carpocystidae). Nell’Ordoviciano sardo questi<br />
fossili, gia noti ai vecchi autori dell’800, si trovano nella Fm. di Portixeddu e sono di<br />
seguito figurate alcune delle forme più significative (da Maccagno, 1965):<br />
Corylocrinus Oocystis Heliocrinites<br />
Dal punto di vista paleoecologico questi blastozoi A) filtratori di particellato alimentare, occupavano<br />
bassi fondali ossigenati ricchi di nutrienti alimentari, condividendo il loro habitat con<br />
B) briozoi, C) brachiopodi e D) edrioasteroidi.<br />
3
Classe Blastoidi<br />
O<br />
Blas. 01 i<br />
I blastoidi comprendono solo pelmatozoi estinti, la cui distribuzione stratigrafica va dallo<br />
Ordoviciano medio?, Siluriano medio al tardo Permiano. Possiedono una teca conica, a forma<br />
di bocciolo o globulare con quattro ordini di placche disposte in simmetria pentamera; cinque<br />
ambulacri (da corti a lunghi) sviluppati in fessure radiali con sottostanti lancette che hanno<br />
lateralmente 2 placchette, ciascuna delle quali regge lunghe brachiole biseriali. Bocca centrale<br />
alla sommità; ano alla sommità all’interno di uno spiracolo più largo, con gonopori nascosti.<br />
Un’idrospira a pieghe è sospesa nella cavità celomatica lungo gli ambulacri; sutura trasversale<br />
dell’idrospira fra le placche radiali e le deltoidi. Otto o dieci di queste strutture (alcune assenti<br />
o ridotte sul lato anale) sono presenti all’interno <strong>della</strong> teca. Le idrospire si aprono sia nel senso<br />
<strong>della</strong> loro lunghezza come fessure sia come paia di pori adiacenti alle placchette laterali ed<br />
inoltre come un grosso poro situato alla loro sommità, lo spiracolo.<br />
In figura le particolarità scheletriche di Pentremites - vista laterale a) e superiore b) del calice a<br />
bocciolo mostrante 4 circoli di placche: 3 basali, 5 radiali, 5 deltoidi e 5 ambulacri complessi;<br />
sezione trasversale <strong>della</strong> teca c) mostrante la disposizione delle lancette sottostanti gli ambulacri<br />
con le faccette laterali lungo le quali si inseriscono le brachiole biseriali; la struttura<br />
dell’idrospira fittamente ripiegata si attacca da un lato alle deltoidi e dall’altro alle radiali; il<br />
particolare dell’ambulacro in sezione tridimensionale prospettica d) mostra le brachiole e la<br />
struttura dell’idrospira.<br />
4
Il peduncolo di solito è lungo e sottile, e si attacca alla teca nella parte distale delle placche<br />
basali tramite faccette peduncolari.<br />
I blastoidi costituiscono la classe più evoluta dei blastozoi, avendo un sistema acquifero molto<br />
avanzato costituito da idrospire. Dal punto di vista evolutivo, si ha nel tempo ad un allungamento<br />
delle strutture ambulacrali e dello sviluppo di una teca da conica a globulare, come è<br />
mostrato nella figura sotto riportata:<br />
A destra Pentremites elegans con pinnule conservate<br />
sopra l’ambulacro.<br />
A - Vista superiore e laterale di<br />
Heteroschisma (Dev. M., Canada)<br />
si notino i corti ambulacri e le<br />
aperture a fessura dell’idrospira.<br />
B - idem di Timoroblastus (Perm.<br />
sup., Timor) mostrante un insolito<br />
aspetto <strong>della</strong> teca e corti ambulacri.<br />
C - Pentremites in vista laterale<br />
(Carbonifero medio).<br />
D - Placoblastus (Dev. m., USA).<br />
E - Drophocrinus (Carb. inf., Irlanda).<br />
F - Deltablastus (Perm. sup.,<br />
Timor) si notino le strie di<br />
accrescimento.<br />
G - Lophoblastus (Carb. inf., Timor).<br />
H - Schizoblastus (Carb. inf., USA)<br />
5
Welcome to<br />
THE ECHINOID DIRECTORY<br />
Designed and created by<br />
Dr. Andrew B. Smith, The Natural History<br />
Museum, London<br />
Morfologia scheletrica degli Echinoidea<br />
Echinidi Regolari<br />
Dollaro delle sabbie ed<br />
affini<br />
Echinidi a cuore o Irregolari<br />
Click su uno dei tipi illustrati per un’introduzione sulla loro<br />
morfologia in Inglese (solo se si è collegati in rete Internet)<br />
Contact and enquiries Accessibility Site map Terms of use © The Natural History Museum, London 2006. All Rights Reserved<br />
Echi. 02
Morfologia schel. Echinidi regolari<br />
I ricci di mare regolari possiedono un guscio a simmetria pentaradiale<br />
composto di dieci doppie file di placche, cinque interambulacrali e cinque<br />
ambulacrali, che formano la corona. Le aree ambulacrali sono costituite<br />
da placche perforate da un singolo o doppio per i pedicelli ambulacrali.<br />
Queste paia di pori sono poste lungo il margine esterno<br />
(adambulacrale) delle aree ambulacrali. La linea di sutura centrale tra<br />
le due file di placche di ciascun’area ambulacrale rappresenta la linea di<br />
sutura periradiale. Le placche interambulacrali si congiungono alle<br />
ambulacrali lungo la sutura adambulacrale, e la linea mediana tra le<br />
due file delle placche interambulacrali è la sutura in<strong>terra</strong>diale.<br />
Echi. 02a<br />
La corona porta due principali aperture, il peristoma e il periprocto. Il<br />
peristoma è circolare e centrale sulla superficie inferiore ed è li che si<br />
apre la bocca. In vita molta parte di questa è coperta da una pelle spessa<br />
che ingloba placchette (nell’esemplare illustrato le placchette inglobate<br />
embricate sono ancora in posto). Tutte le aree ambulacrali ed interambulacrali<br />
convergono attorno alla bocca.<br />
All’apice <strong>della</strong> teca, nel punto dove so originano le aree ambulacrali, è<br />
posto il disco apicale. Questo anello di dieci placche contorna l’apertura<br />
1
anale, ed è sempre posto sulla superficie aborale. Il disco apicale si compone<br />
di cinque placche genitali e cinque placche oculari che contornano<br />
una membrana flessibile con placche contenente l’ano. Questa membrana<br />
flessibile è chiamata periprocto. La disposizione delle placche del disco<br />
apicale forma un importante carattere tassonomico.<br />
L’esatta disposizione sul guscio delle placche ambulacrali è molto<br />
variabile ed estremamente importante. Ulteriori informazioni su questo<br />
carattere sono fornite in disposizione delle placche ambulacrali.<br />
Tutte le placche portano radioli, che si attaccano e si articolano sui tubercoli.<br />
Nell’esemplare qui illustrato c’é un singolo tubercolo primario su ciascuna<br />
placca interambulacrale, e numerosi piccoli tubercoli e granuli lungo<br />
i margini esterni delle placche interambulacrali e l’area periradiale<br />
delle placche ambulacrali. I tubercoli possono essere perforati o imperforati,<br />
e crenulati o lisci: ulteriori informazioni sono disponibili in struttura<br />
dei tubercoli. I radioli, con una gran varietà di forme, servono per<br />
difesa e/o locomozione.<br />
All’interno <strong>della</strong> teca si trova un complesso apparato chiamato lanterna<br />
d’Aristotele. Questa funziona come le fauci degli animali, comprende 50<br />
elementi scheletrici e 60 muscoli. La morfologia <strong>della</strong> lanterna è molto<br />
importante per la tassonomia delle categorie superiori di questo gruppo.<br />
Un’altra struttura interna importante è costituita dalla flangia peristomale.<br />
Questa è una struttura composta dalle placche immediatamente<br />
attorno al peristoma e forma le zone di ancoraggio per i muscoli che<br />
fanno funzionare la lanterna di Aristotele. Anche le modalità di come si<br />
compone e si muove quest’ultima forniscono utili caratteri tassonomici.<br />
Da: http://www.nhm.ac.uk/research-curation/projects/echinoid-directory/morphology/regulars/intro.html e siti correlati,<br />
(trad. M. Gnoli)<br />
2
Placchette ambulacrali<br />
La disposizione delle placchette ambulacrali degli echinidi regolari costituisce uno dei più<br />
importanti caratteri per la loro classificazione. Le aree ambulacrali sono formate da una doppia<br />
serie di placchette, ciascuna perforata da un paio di pori. Ciascun paio connette il pedicello<br />
ambulacrale esterno all’ampolla interna del sistema acquifero radiale. Il paio di pori è posto<br />
verso il margine esterno di ciascuna placchetta ambulacrale, vicino alla sutura radiale <strong>della</strong><br />
placchetta. La sutura radiale segna la giunzione fra le placchette ambulacrali e quelle interambulacrali,<br />
mentre la sutura periradiale è la linea centrale che separa le due colonne delle<br />
placche ambulacrali fra loro. Il margine superiore <strong>della</strong> placca ambulacrale è la sutura adapicale,<br />
mentre quello inferiore è la sutura adorale.<br />
Nel più semplice dei casi, ciascun paio di pori è posto su una placchetta allungata che si estende<br />
al periradio. Questa ha uno o più tubercoli che non si sovrappongono mai alla placchetta<br />
adiacente. Questa disposizione, mostrata nella fotografia in alto a sinistra, prende il nome<br />
d’ambulacro semplice. La disposizione delle placche ambulacrali è detta essere unigeminata.<br />
Nella gran maggioranza dei tipi di placche ambulacrali, comunque, si associano assieme a<br />
formare placche ambulacrali composte e complesse. Nell’immagine in alto a destra, ci sono tre<br />
paia di pori per ciascun tubercolo ambulacrale principale. Questa è la disposizione composta.<br />
Il modo col quale le placchette si uniscono per formare le placche composte è molto vario e<br />
molto importante dal punto di vista tassonomico. Alcuni dei più comuni tipi di disposizione<br />
nelle connessioni delle placchette sono illustrati nella prossima pagina.<br />
Per ultimo, ciascun paio di pori stesso può apparire piccolo e circolare con ciascun poro separato<br />
dall’altro da una piccola partizione (chiamato non-congiunto; come nella foto in alto a<br />
destra) o allungato con i due pori connessi da un basso solco (chiamato congiunto; come<br />
nell’immagine in alto a sinistra).<br />
Echi. 03
Tipi di placchette ambulacrali<br />
Nelle vere placchette ambulacrali composte gli elementi sono fusi fra loro a formare<br />
una singola placchetta composta con margine periradiale a “V”. Quando il<br />
guscio si “disarticola” le placchette composte non si disgregano nei loro elementi<br />
individuali. Al contrario le placchette semi-composte, conservano l’aspetto delle<br />
placchette semplici, possedendo la sutura periradiale a “V”, ma due o più di queste<br />
mostrano la sovracrescita di un tubercolo esterno che le tiene unite. Le placchette<br />
pseudocomposte differiscono per avere uno solo dei due elementi fortemente<br />
ingrandito che regge i tubercoli, essendo l’altro ridotto ad una minuscola placchetta<br />
poco più grande di un paio di pori. Queste placchette si dissociano rapidamente<br />
e non sono fuse.<br />
Placchette con due paia di pori ambulacrali su ogni tubercolo primario sono dette<br />
bigeminate, e quelle con tre paia di pori su una placchetta composta sono dette<br />
trigeminate. Quando ci sono più di tre paia di pori su una placchetta composta, la<br />
placchetta è detta essere poligeminata. Paia di pori possono essere disposte su una<br />
singola fila (uniseriali), su due file (biseriali), tre file (triseriali), o ad arco più o<br />
meno obliquo alla sutura verticale.<br />
Semplici<br />
(unigeminate)<br />
Bigeminata<br />
Trigeminata<br />
(Diadematoide)<br />
Pseudocomposta<br />
Semicomposta<br />
Poligeminata<br />
(Diadematoide)<br />
Arbacioide<br />
Goniopigide<br />
Trigeminata<br />
(Echinoide)<br />
Phymosomatide<br />
Stomopneustide<br />
Poligeminata<br />
(Echinoide)<br />
Da: http://www.nhm.ac.uk/research-curation/projects/echinoid-directory/morphology/regulars/regamb1.html<br />
Echi. 04
Tubercoli dei regolari<br />
Echi. 04a<br />
I tubercoli costituiscono il punto d’attacco dei radioli alla teca. Si presentano<br />
in varie fogge, ma tutti sono costituiti nello stesso modo. Al<br />
centro c’é una protuberanza emisferica, il mamellone (m), che rappresenta<br />
la superficie d’articolazione del radiolo. Si presenta liscio e lucido<br />
essendo ricoperto di solida calcite. Può essere sia perforato (come<br />
nella fotografia sopra) sia imperforato (sotto a destra), la perforazione<br />
alloggia il legamento che va dalla placca al radiolo. Il mamellone<br />
è circondato da un sottile parapetto che nell’esemplare illustrato è<br />
liscio, ma è crenulato nell’esemplare fotografato in basso a sinistra.<br />
Le crenulazioni si accoppiano a crenulazioni simili alla base del radiolo<br />
che aiutano a tenerlo in posto. Il fatto che i tubercoli siano perforati o<br />
imperforati, crenulati o non crenulati, costituisce un importante carattere<br />
tassonomico. Il mamellone è posto all’apice di una protuberanza<br />
conica, il bottone (b). Questo è il luogo dove si ancora il tessuto connettivo<br />
(impiegato per tenere il radiolo in posizione di difesa). Per ultima,<br />
la zona spesso depressa, che è definita come l’areola (ar) è l’area<br />
dove si ancora il muscolo impiegato per muovere il radiolo.<br />
In ogni placca può esserci un solo tubercolo principale, o parecchi. Di<br />
solito c’é una varietà di tubercoli e granuli più piccoli. Nell’esemplare<br />
sopra illustrato c’é un distinto anello di tubercoli secondari che circonda<br />
l’areola del tubercolo principale. Questi sono i tubercoli scrobicolari o<br />
scrobicoli che formano l’anello scrobicolare attorno al tubercolo<br />
principale.<br />
I tubercoli illustrati di seguito sono di un diadematoide (crenulato e<br />
perforato) e di un camarodonte (non crenulato ed imperforato); questo<br />
1
Da: http://www.nhm.ac.uk/research-curation/projects/echinoid-directory/morphology/regulars/regtub.html<br />
2
Radioli dei regolari<br />
I radioli sono costituiti da calcite ed impiegati dagli echinidi regolari<br />
per la difesa e la locomozione. Ogni radiolo si articola su un tubercolo<br />
del guscio. Nel cidaride sotto figurato vi sono grossi radioli<br />
primari (in bianco) ed un gran numero di radioli (spine) più piccoli<br />
secondari attorno alla loro base. I cidaridi sono insoliti per avere<br />
un guscio esterno di calcite, il cortex, che rapidamente comincia ad<br />
essere ricoperto da alghe che provvedono a mimetizzare l’animale.<br />
In tutti gli altri echinidi, un tessuto cutaneo vivo copre i radioli.<br />
Questi sono uniti al guscio da tessuto molle e sono subito persi<br />
dopo la morte, quindi raramente conservati fossili in situ.<br />
I radioli mostrano un’ampia gamma di forme e dimensioni (vedi<br />
sotto). Quelli di Diadema (sotto a sinistra) sono lunghi come aghi<br />
e vengono di solito usati per difesa contro i predatori. In Podophora<br />
(sotto a destra) gli articoli aborali sono modificati a formare<br />
una volta intrecciata, un adattamento per vivere nell’ambiente<br />
di scoglio ad alta energia. Alcuni cidaridi sviluppano grossi<br />
radioli glandiformi.<br />
La struttura dei radioli è molto variabile e fornisce un importante<br />
carattere tassonomico. Il radiolo può essere liscio o presentare<br />
un’ornamentazione a coste, bottoni e granuli. In sezione trasversale<br />
possono essere cavi o massicci.<br />
Psammechinus Diadema Tylocidaris Podophora<br />
Da: http://www.nhm.ac.uk/research-curation/projects/echinoid-directory/morphology/regulars/spine.html<br />
Echi. 05
Struttura dei radioli<br />
In un singolo radiolo si possono distinguere tre parti, una lunga asta,<br />
un corto collare ed una base. La superficie concava d’articolazione che<br />
si attacca al mamellone del tubercolo, è detta acetabulum, e può essere<br />
sia perforata che imperforata. Muscoli e dispositivo di tenuta connettono<br />
la base del radiolo al tubercolo e ne permettono il movimento<br />
individuale. In molti echinidi regolari l’asta è liscia ed appare finemente<br />
striata sotto ingrandimento. In pochi gruppi, specialmente negli<br />
arbacioidi, l’asta ha un aspetto vitreo ed è ricoperta da una sottile<br />
cuticola. Nei cidaridi c’é uno spesso strato di “cortex” e l’asta è generalmente<br />
ornata da solchi, spine o granuli. Nei diademoidi è visibile<br />
una struttura molto caratteristica dove l’asta è costituita da spire ed<br />
appare segmentata, ed il radiolo è detto verticillato).<br />
I radiali sono solitamente colorati da un pigmento incorporato nella<br />
calcite dello scheletro.<br />
In sezione trasversale (microfotografie al SEM sotto) i radioli possono<br />
essere cavi o massicci. Lo spazio vuoto di un radiolo è detto lumen.<br />
Quando il centro è riempito da calcite porosa disorganizzata, il “tappo”<br />
centrale è chiamato midollo.<br />
Da: http://www.nhm.ac.uk/research-curation/projects/echinoid-directory/morphology/regulars/spine2.html<br />
Echi. 06
Peristoma dei regolari<br />
Il peristoma è l’apertura del guscio associata alla bocca. Nel vivente<br />
è coperta da una membrana di tessuto connettivo flessibile. Nei<br />
cidaroidi (in alto a sinistra) ed echinoturioidi le placche sono disposte<br />
come serie embricate e si estendono sopra il peristoma (a),<br />
ciascuna è perforata da un paio di pori dove nel vivente escono i pedicelli.<br />
Ci può anche essere una sola o doppia serie di placche nonambulacrali<br />
(n) in posizione interambulacrale. In tutti gli altri tipi di<br />
membrane peristomali ci sono cinque paia di placchette boccali,<br />
che sono placchette ambulacrali ovali portanti ciascuna un singolo<br />
pedicello. La parte rimanente <strong>della</strong> membrana è piuttosto poco<br />
sclerotizzata, avendo solo piccole placchette.<br />
Echinidi diversi dai cidaroidi, possiedono sacche di tessuto molle che<br />
sono delle estensioni esterne del celoma interno. Queste sono necessarie<br />
per compensare le variazioni di volume all’interno del guscio<br />
rigido <strong>della</strong> lanterna quando questa si muove durante l’assunzione di<br />
cibo. Queste “sacchette boccali” emergono dall’orlo esterno dell’apertura<br />
peristomale immediatamente adiacenti alla sutura radiale<br />
delle aree ambulacrali. La loro posizione è evidenziata da distinte<br />
tacche sull’orlo del peristoma (indicate dalla freccia nell’immagine a<br />
destra). Queste sono le tacche boccali.<br />
Da: http://www.nhm.ac.uk/research-curation/projects/echinoid-directory/morphology/regulars/peristome.html<br />
Echi. 07
Flangia (cintura) peristomale<br />
La flangia peristomale si trova sulla parte interna <strong>della</strong> “corona” al<br />
margine del peristoma. È composta di proiezioni calcaree che agiscono<br />
da piattaforma per il sostegno e l’attacco dei muscoli per l’estrusione<br />
ed il ritiro <strong>della</strong> lanterna. Nei cidaroidi (in alto a destra) queste<br />
proiezioni sono costituite interamente dalle placche ambulacrali, e<br />
sono chiamate apofisi (ap). I muscoli espulsori si connettono vicino<br />
alla base centralmente, mentre i muscoli retrattori sono connessi<br />
alle estremità superiori. Negli echinidi regolari non cidaroidi (in alto a<br />
sinistra) ci sono di solito entrambi gli elementi ambulacrali ed interambulacrali.<br />
Quelli ambulacrali sono strutture rilevate digitiformi<br />
dette auricole (au) ed è qui che si connettono i muscoli retrattori. I<br />
muscoli espulsori si attaccano alle basse pareti interambulacrali che<br />
si alternano alle prime.<br />
Da: http://www.nhm.ac.uk/research-curation/projects/echinoid-directory/morphology/regulars/girdle.html<br />
Echi. 08
Disco apicale e Periprocto Echi. 09<br />
Il disco apicale di un echinide regolare è composto di cinque placche oculari (indicate I-V) e cinque<br />
placche genitali (indicate 1-5). Le placche oculari sono microscopicamente perforate e alloggiano la<br />
sommità del sistema acquifero; una placca oculare è posta alla sommità di ogni ambulacro. Le<br />
placche genitali sono di solito più larghe e perforate da un poro più grosso, il gonoporo. È da<br />
queste aperture che l’animale rilascia nell’acqua i prodotti sessuali (uova e sperma) per la<br />
riproduzione. Una di queste placche, G2, è più grande delle altre e fittamente perforata (visibile al<br />
microscopio), il madreporite. Questo cerchio di placche circonda una membrana flessibile<br />
chiamata periprocto, al centro del quale c’é l’ano. Il margine esterno del periprocto può essere<br />
liscio ed ovale, sub-circolare o (come nell’esemplare sopra) angolato. Il sistema di placche del disco<br />
apicale può essere fermamente unito alla “corona” (è così che si presenta nei fossili), embricato o<br />
solo appoggiato allo scheletro (conseguentemente molto raro come fossile).<br />
Nell’esemplare sopra figurato il cerchio di placche genitali è parzialmente interrotto dalle<br />
placche oculari posteriori (V e I separate dalle adiacenti placche genitali). Questa disposizione<br />
viene detta emiciclica. Quando tutte le cinque placche oculari formano un anello continuo<br />
con le placche oculari separate dalla disposizione delle placche del periprocto, il disco è<br />
detto diciclico. Dove tutte cinque le placche oculari giacciono a contatto del periprocto e<br />
sono separate dalle placche genitali, il disco è chiamato monociclico. In alcuni taxa esiste<br />
una distinta placca periproctale ingrandita, la placca sopra-anale. Questa può essere saldamente<br />
connessa al circolo delle placche genitali, o essere presente solo negli stadi giovanili.<br />
Importanti variazioni nella disposizione delle placche sono di seguito figurate (placche oculari in<br />
verde, placche genitali in giallo e periprocto in nero. Quella perforata è il madreporite).<br />
Monociclico Diciclico Emiciclico Placca sopra-anale<br />
Da: http://www.nhm.ac.uk/research-curation/projects/echinoid-directory/morphology/regulars/girdle.html
Lanterna d’Aristotele<br />
La lanterna è usata per mordere e triturare ed è composta di 50 elementi<br />
scheletrici mossi da 60 muscoli. È quasi completamente interna, sebbene<br />
le punte dei denti si estendano all’esterno <strong>della</strong> bocca. Ci sono due gruppi<br />
principali di muscoli che connettono la lanterna al guscio e questi si ancorano<br />
alla flangia peristomale. Gli espulsori che fanno uscire la lanterna,<br />
mentre i muscoli retrattori la sollevano. Potenti muscoli fra le emipiramidi<br />
chiudono i denti e provvedono ad una forte azione masticatoria.<br />
Struttura <strong>della</strong><br />
lanterna<br />
Tipi di Lanterne Tipi di Lanterne<br />
Da: http://www.nhm.ac.uk/research-curation/projects/echinoid-directory/morphology/regulars/girdle.html<br />
Echi. 09a
Struttura <strong>della</strong> “Lanterna”<br />
La lanterna é composta di 50 elementi scheletrici e circonda<br />
l’ingresso dello stomaco. Qui é mostrata la lanterna di un cidaroide<br />
in veduta posteriore e laterale. Gli elementi più grossi<br />
sono le emipiramidi (h). Queste sono piastre a forma di<br />
zeppa saldamente unite a formare le cinque piramidi <strong>della</strong><br />
lanterna. C’è una tacca fra le due emipiramidi nella parte terminale<br />
adorale <strong>della</strong> loro sutura, questa è il foramen magnum<br />
(fm). Questo è piccolo nei cidaridi (t sopra), ma è tipicamente<br />
molto profondo ed a forma di “V” negli altri echinidi<br />
regolari. L’elemento adapicale “a spalla” di ogni emipiramide<br />
ospita un piccolo elemento assiale, l’epifisi (e). In posizione<br />
radiale c’é un singolo elemento linguiforme, la rotula (r), che<br />
si pone direttamente fra due epifisi, una da ciascuna adiacente<br />
piramide. La rotula funziona da elemento d’unione delle<br />
due adiacenti emipiramidi. Un singolo dente (t) giace addossato<br />
all’interno di ogni emipiramide, ed è visibile dal foramen<br />
magnum. Per ultimi ci sono dei minuti elementi spatoliformi, i<br />
compassi (c), che sovrastano ciascuna rotula. I compassi<br />
servono per alzare ed abbassare le pareti celomatiche <strong>della</strong><br />
lanterna, quando questa si muove su e giù, per masticare il<br />
cibo. Le piattaforme d’attacco dei muscoli che formano le<br />
flangie peristomali (g) attorno alla bocca, sono visibili alla<br />
base.<br />
Da: http://www.nhm.ac.uk/research-curation/projects/echinoid-directory/morphology/regulars/lantern2.html<br />
Echi. 10
Echi. 11<br />
Principali tipi di denti – vista esterna<br />
Quattro sono i tipi di lanterne degli echinidi regolari, variando nella<br />
profondità del foramen magnum, sezione trasversale dei denti e nello<br />
sviluppo delle epifisi. Qui sono illustrati i segmenti di lanterna di<br />
ciascun tipo, visti dall’esterno.<br />
Cidaroide. Lanterna minuta con un minuscolo foramen magnum. Epifisi<br />
piccole che non si proiettano e denti di sezione ad “U”. Caratteristica<br />
di tutti i cidaridi.<br />
Aulodonte. Lanterna con foramen magnum a forma di profonda “V”,<br />
con epifisi che tendono ad avere piccole proiezioni libere digitiformi.<br />
Denti con sezione a forma di “U”. Caratteristica degli echinoturioidi,<br />
diadematoidi e pedinoidi.<br />
Stirodonte. In sostanza come l’aulodonte, ma con denti con sezione a<br />
“T”. Caratteristica di arbacioidi, salenioidi e stomopneustidi.<br />
Camarodonte. Foramen magnum come la lanterna Aulodonte, ma le<br />
epifisi sono più grosse e s’incontrano superiormente al foramen magnum<br />
formando un arco continuo. Denti con sezione a “T”. Caratteristica<br />
di temnopleuridi ed echinidi.<br />
Da: http://www.nhm.ac.uk/research-curation/projects/echinoid-directory/morphology/regulars/lanterntypes1.html
Echi. 12<br />
Principali tipi di denti - vista interna<br />
Quattro sono i tipi di denti nella lanterna degli echinidi regolari, variando<br />
nella profondità del foramen magnum, sezione trasversale dei denti<br />
e nello sviluppo delle epifisi. Qui sono illustrati i segmenti di lanterna di<br />
ciascun tipo, visti dall’interno.<br />
Cidaroide. Lanterna minuta con un minuscolo foramen magnum. Epifisi<br />
piccole che non si proiettano e denti di sezione ad “U”. Caratteristica di<br />
tutti i cidaridi.<br />
Aulodonte. Lanterna con foramen magnum a forma di profonda “V”,<br />
con epifisi che tendono ad avere piccole proiezioni libere digitiformi.<br />
Denti con sezione a forma di “U”. Caratteristica degli echinoturioidi,<br />
diadematoidi e pedinoidi.<br />
Stirodonte. In sostanza come l’aulodonte, ma con denti con sezione a<br />
“T”. Caratteristica di arbacioidi, salenioidi e stomopneustidi.<br />
Camarodonte. Foramen magnum come la lanterna Aulodonte, ma le<br />
epifisi sono più grosse e s’incontrano superiormente al foramen magnum<br />
formando un arco continuo. Denti con sezione a “T”. Caratteristica<br />
di temnopleuridi ed echinidi.<br />
Da: http://www.nhm.ac.uk/research-curation/projects/echinoid-directory/morphology/regulars/lanterntypes1.html
Chiave ai principali gruppi di Echinoidea<br />
1a. Teca composta da più di 20 file di<br />
placche (più di 2 file di ambulacri o interambulacrali<br />
in ciascuna area)<br />
Gruppo principale di echinidi Paleozoici<br />
2a. Teca a simmetria bilaterale sovrapposta<br />
ad una pentamera; periprocto posto<br />
posteriormente perlopiù esterno al disco<br />
apicale. Irregolari<br />
3a. Flangia peristomale rudimentale o<br />
assente.<br />
Gruppo evoluto Echinidi, gruppi<br />
Cidaroidea o Euechinoidea<br />
1b. Teca composta da 20 file di placche;<br />
aree ambulacrali ed interambulacrali<br />
ciascuna composta da 2 file di placche<br />
Vai alla 2<br />
2b. Simmetria pentaradiale; periprocto<br />
circondato da placchette del disco apicale<br />
Vai alla 3<br />
3b. Flangia peristomale ben sviluppata,<br />
composta di auricole ambulacrali e/o<br />
apofisi interambulacrali.<br />
Vai alla 4 (Echinidi regolari)<br />
Echi. 12a
4a. Flangia peristomale sempre presente<br />
4b. Flangia peristomale composta da<br />
e composta solo da apofisi interambulacrali<br />
auricole ambulacrali (au). Ambulacri<br />
(ap). Placche ambulacrali semplici che si<br />
sempre composti.<br />
estendendono nel peristoma.<br />
Cidaroidea<br />
Euechinoidea<br />
Da: http://www.nhm.ac.uk/research-curation/projects/echinoid-directory/taxa/key.jsp?id=388
Gruppo principale (stem group) degli Echinidi paleozoici<br />
[= Perischoechinoidea McCoy, 1849. Include ordini Bothriocidaroida Zittel, 1879, Echinocystitoida<br />
Jackson 1912, Megalopoda MacBride & Spencer, 1938, Palaechinoida Haeckel,<br />
1866]<br />
Diagnosi<br />
Echinidi con:<br />
• Teca composta da più di due file di placche ambulacrali e/o<br />
interambulacrali in ciascuna area;<br />
• No flangia peristomale.<br />
Echi. 12b<br />
Distrib. Ordoviciano medio - Permiano superione; cosmopoliti.<br />
Tutti gli Echinidi viventi possiedono una teca dove si alternano aree di<br />
placche ambulacrali ed interambulacrali, ciascun’area è costituita solo da<br />
due file di placche. Questa è una sinapomorfia per il gruppo corona (crown<br />
group) degli Echinoidea, ed è anche vero per tutti i gruppi di echinidi post-<br />
Paleozoici, Tiarechinus, ne hanno solo una. Contrariamente, tutti gli echinidi<br />
paleozoici possiedono una teca che si discosta da questo modello base;<br />
essendo Miocidaris il solo taxon paleozoico a seguire il modello del gruppo<br />
corona. I Bothriocidaridi hanno una teca composta esclusivamente da placche<br />
ambulacrali, mentre molti altri gruppi possiedono sia placche ambulacrali<br />
che interambulacrali. Questi gruppi ricadono nel gruppo base (stem group)<br />
degli Echinoidea.<br />
Note<br />
Una analisi filogenetica preliminare degli echinidi del Paleozoico fu pubblicata<br />
da Smith (1984). In ogni caso, è necessaria una più rigorosa analisi per<br />
stabilire un consistente bagaglio di conoscenze per ordinare questi gruppi<br />
molto vari. In questa sede, il gruppo base (stem group) viene diviso in un<br />
numero di taxa, che, in alcuni casi, probabilmente rappresenta piuttosto delle<br />
gradazioni invece che dei gruppi monofiletici (cladi).<br />
Un resoconto <strong>della</strong> differenziazione morfologica degli echinidi del Paleozoico<br />
ci è fornita da Kier (1965) e da Jackson (1912) e rimane la più esauriente per<br />
la maggior parte dei taxa.<br />
1
I seguenti gruppi rimangono non stabiliti (indeterminati) ed omessi dalle<br />
moderne chiavi di classificazione e discussione:<br />
Lanternarius Regnell, 1956<br />
Kongielechinus Jesionek-Szymanska, 1979<br />
Protocidaris Whidborne, 1898<br />
Rhoechinus Keeping, 1876<br />
Silurocidaris Regnell, 1956<br />
Tornquistellus Berg, 1899<br />
Jackson, R. T. 1912 Boston Society for Natural History Memoirs 7, p. 296, pl.<br />
24, fig. 4, pl. 25, figs 1,2.<br />
Kier, P. M. 1965. Evolutionary trends in Paleozoic echinoids. Journal of<br />
Paleontology 39, 436-465, pls 55-60.<br />
Kier, P. M. 1966. Noncidaroid Paleozoic echinoids. Pp. U298-312 in Moore, R.<br />
C. (ed.). Treatise on Invertebrate Paleontology. University of Kansas Press &<br />
Geological Society of America, Boulder.<br />
Smith, A. B. 1984. Echinoid Palaeobiology. George Allen & Unwin, London<br />
Da: http://www.nhm.ac.uk/research-curation/projects/echinoid-directory/taxa/taxon.jsp?id=1<br />
2
Gruppi principali (stem group) Euechinoidea, Cidaroidea<br />
e/o gruppo stelo evoluto Echinoidea<br />
Diagnosi<br />
Euechinoidi con:<br />
• Placche ambulacrali semplici o al massimo tendenti a pseudocomposte;<br />
• Flangia peristomale assente, o con deboli punti di atacco muscolare ai<br />
margini mediani e radiali delle aree interambulacrali.<br />
Echi. 13<br />
Distrib. Triassico medio - sup. Europa, Nord e Sud America.<br />
Il gruppo corona Euechinoidea viene distinto dai Cidaroidea dalla loro flangia<br />
peristomale, che è composta da proiezioni ambulacrali radiali dette auricole. I<br />
Cidaroidea al contrario non possiedono componenti ambulacrali nella flangia<br />
peristomale, che è costituita interamente da apofisi. Il gruppo principale (stem<br />
group) Echinoidea non ha flangia peristomale, ma la loro teca è composta da più<br />
di 20 file di placche. Nel Triassico, in ogni caso, un piccolo gruppo di echinidi<br />
possiede la struttura <strong>della</strong> teca del “gruppo corona” (quella composta da solo 20<br />
file di placche), ma non hanno niente di più d’una flangia peristomale molto<br />
rudimentale.<br />
Note<br />
Nei Triadocidaridae e Paurocidaridae le placche ambulacrali più adorali mostrano<br />
spesso ispessimenti marginali che Kier (1984) interpreta come abbozzi di<br />
auricole. Allo stesso modo, i Serpianotiaridae possiedono nelle loro placche più<br />
interambulacrali delle sporgenze radiali angolate verso gli ambulacri. Queste<br />
differiscono dalle più consistenti apofisi dei cidaroidi. Kier (1984) e Smith (1994)<br />
considerano entrambe queste famiglie come gruppi stelo Euechinoidea.<br />
Kier (1984) realizzò per primo il significato filogenetico <strong>della</strong> struttura <strong>della</strong><br />
flangia peristomale in questo gruppo. La loro analisi filogenetica fu fatta da Smith<br />
(1994).<br />
1
I seguenti taxa sono basati su frammenti o radioli e sono poco conosciuti per<br />
essere classificati. Sono stati omessi dalle chiavi identificative e discussioni:<br />
Batheritiaris Vadet, 2001<br />
Braunechinus Vadet, 2001<br />
Kieritiaris Vadet, 2001<br />
Thielicidaris Vadet, 2001<br />
Wissmannechinus Vadet, 2001<br />
Zardinitiaris Vadet, 2001<br />
Kier, P.M. 1977. Triassic echinoids. Smithsonian Contributions to Paleobiology<br />
30, 1-88.<br />
Kier, P. M. 1984. Echinoids from the Triassic (St Cassian) of Italy, their lantern<br />
supports, and a revised phylogeny of Triassic echinoids. Smithsonian<br />
Contributions to Paleobiology 56, 1-41.<br />
Smith, A. B. 1994. Triassic echinoids from Peru. Palaeontographica A 233, 177-<br />
202.<br />
Da: http://www.nhm.ac.uk/research-curation/projects/echinoid-directory/taxa/taxon.jsp?id=860 e siti<br />
correlati (trad. M. Gnoli)<br />
2
Morfologia schel. degli irregolari<br />
I ricci a cuore (spatangidi) ed i loro parenti (holasteroidi, disasteroidi)<br />
possiedono un guscio a simmetria bilaterale formato da 10 doppie<br />
file di placche, cinque file interambulacrali (siglate 1-5) e cinque<br />
file ambulacrali (siglate I-V). Le placche ambulacrali sono perforate<br />
da pori singoli o doppi per i pedicelli ambulacrali. L’ambulacro<br />
anteriore è l’ambulacro III. Nell’esemplare illustrato le placche che<br />
formano la corona sono facilmente identificabili; I margini delle placche<br />
sono chiari ed il loro centro scuro.<br />
Nel guscio ci sono due aperture principali, il peristoma ed il periprocto.<br />
L’apertura ovale o a forma di “D” nella parte inferiore costituisce<br />
il peristoma, ed è lì che è posta la bocca. Tutti i cinque ambulacri convergono<br />
verso il peristoma, e perlopiù tutti sono situati prossimi al<br />
margine anteriore. Il periprocto ospita l’apertura anale ed è sempre<br />
posto verso la parte posteriore. Secondo la specie, può aprirsi leggermente<br />
sopra l’ambito tanto da essere visibile dall’alto, o sotto, ed è<br />
visibile in veduta orale. È circondato dalle placche dell’interambulacro<br />
5.<br />
All’apice <strong>della</strong> teca, nel punto d’origine delle aree ambulacrali, sta il<br />
disco apicale. Questo è composto di un piccolo numero di placche che<br />
Echi. 14<br />
1
sono le prime a generarsi nella metamorfosi. Si compone di una a<br />
quattro placche genitali perforate dai gonopori che connettono le<br />
gonadi all’esterno, e da cinque placche oculari alle estremità d’accrescimento<br />
di ciascun ambulacro. La disposizione delle placche del<br />
disco apicale fornisce un importante numero di caratteri tassonomici.<br />
Le porzioni apicali degli ambulacri si sviluppano variamente in petali,<br />
aree specializzate di paia di pori dai quali fuoriescono i pedicelli respiratori.<br />
La forma e lo sviluppo dei petali sono pure importanti dal<br />
punto di vista tassonomico. In alcuni taxa tutti i cinque ambulacri<br />
sono identici. Gli spatangidi possiedono i petali tipicamente infossati,<br />
mentre altri gruppi possiedono tutti gli ambulacri a livello <strong>della</strong> superficie.<br />
Spesso l’ambulacro anteriore (ambulacro III) si differenzia<br />
dagli altri, come nella forma illustrata. Il grado di differenziazione<br />
dell’ambulacro frontale è importante: varia dall’essere a livello a<br />
profondamente infossato, e lo sviluppo delle paia di pori in quest’ambulacro<br />
varia anch’esso considerevolmente. I pedicelli ambulacrali di<br />
quest’area <strong>della</strong> teca sono responsabili <strong>della</strong> creazione del tunnel<br />
respiratorio nelle specie infossate. Nelle aree ambulacrali possono<br />
essere presenti <strong>della</strong> paia di pori ingranditi nel retro del guscio per la<br />
costruzione di un tunnel per l’espulsione dei rifiuti.<br />
Sulla superficie orale, la disposizione delle placche dell’interambulacro<br />
posteriore (sotto il peristoma) è tassonomicamente importantissima.<br />
Questa area prende il nome di piastrone, ed è spesso la parte<br />
più fittamente tubercolata <strong>della</strong> teca.<br />
Radioli e tubercoli sono generalmente piuttosto piccoli, sebbene<br />
alcuni taxa possiedono grossi tubercoli primari sulla loro superficie<br />
orale che reggono lunghe spine acuminate. I tubercoli aborali sono<br />
sempre più sottili e più fitti di quelli orali. I tubercoli sono sempre<br />
perforati e tipicamente crenulati, sebbene questo gruppo mostri<br />
moltissime specializzazioni nella struttura di entrambi tubercoli e<br />
spine. Le spine sono lunghe e sottili, spesso con punte a spatola. I<br />
ricci a cuore dell’infauna possiedono delle zone con spine molto fini<br />
che sono chiamate fasciole, che permettono loro di vivere in sedimenti<br />
fini. Forma e posizione dettagliata di queste fasciole è molto<br />
importante dal punto di vista tassonomico.<br />
I ricci a cuore sono depositivori usando i loro pedicelli ambulacrali<br />
orali per ammassare il detrito. Non possiedono lanterna (anche nella<br />
prima parte dell’ontogenesi), e neanche la flangia peristomale.<br />
Da: http://www.nhm.ac.uk/research-curation/projects/echinoid-directory/morphology/hearts/intro.html e siti correlati,<br />
(trad. M. Gnoli)<br />
2
Piastrone<br />
Echi. 15<br />
Piastrone è il nome dato alla zona ingrandita dell’area interambulacrale<br />
<strong>della</strong> superficie inferiore dietro la bocca. La disposizione delle placche in<br />
quest’area fornisce importanti caratteri tassonomici. Una prima placca<br />
interambulacrale è quella al margine del peristoma, il labbro (L). La<br />
sua morfologia è estremamente variabile, da una mezza luna a laminare.<br />
Le prossime due placche possono essere ingrandite come sternali. In<br />
molti spatangidi i due sternali sono circa ugualmente sviluppati e formano,<br />
come sopra, virtualmente l’intero piastrone. Il piastrone è quindi<br />
chiamato amfisterno. A volte, le adiacenti placche ambulacrali possono<br />
unirsi centralmente così da separare il labbro dagli sternali successivi: il<br />
piastrone è chiamato disgiunto. Ci possono essere anche delle altre<br />
seconde piastre dietro le sternali, le episternali. Infine, in alcuni<br />
olasteridi e spatangidi una singola placca interambulacrale può trovarsi<br />
dietro (posteriore) il piastrone, la placca rostrale.<br />
Il piastrone può essere completamente coperto da tubercoli e spine,<br />
oppure questi possono essere presenti solo nella sua parte posteriore.<br />
Alcuni dei più importanti tipi di piastroni sono mostrati nella pagina<br />
seguente.<br />
Da: http://www.nhm.ac.uk/research-curation/projects/echinoid-directory/morphology/hearts/plastron1.html
Tipi di Piastrone<br />
Protosterno Meridosterno Amfisterno<br />
Metasterno Disgiunto (Amfisterno)<br />
Da: http://www.nhm.ac.uk/research-curation/projects/echinoid-directory/morphology/hearts/plastron2.html<br />
Echi. 15a
Docce e rigonfiamenti<br />
Nei cassidulidi, la struttura del guscio che circonda il peristoma<br />
è particolarmente importante per la classificazione. Gli ambulacri<br />
in quest’area sono tipicamente ingranditi, i pori diventano<br />
molto fitti e da questi fuoriescono specializzati tubercoli per<br />
l’alimentazione. I pori possono essere singoli o doppi e si dispongono<br />
in una singola linea irregolare, su due colonne, o su<br />
una colonna esterna con pori separati interni come sopra.<br />
Queste zone allargate di pori sono chiamate docce. I due pori<br />
più larghi vicini al peristoma ospitano tubercoli alimentari<br />
sensoriali che sono leggermente più piccoli degli altri. Questi<br />
sono i pori boccali (bp). Le aree interambulacrali adiacenti al<br />
peristoma possono essere indifferenziate, o essere ingrandite e<br />
rigonfie a formare distinte proiezioni a forma rilevata o spine.<br />
Questi sono i rigonfiamenti e le loro pareti adorali sono<br />
coperte da piccole spine che servono per dirigere le particelle<br />
alimentari dentro la bocca.<br />
Echi. 16
Tubercoli<br />
La superficie di un riccio a cuore è ricoperta da una quantità di tubercoli<br />
che servono per l’articolazione dei radioli e pedicellarie. Nell’esemplare<br />
figurato i tubercoli sono di varie dimensioni, ed i più grossi leggermente<br />
infossati. Tutti sono costruiti nello stesso modo. La base del radiolo sta e<br />
si articola su di un mamellone emisferico centrale. Questo è composto<br />
di calcite policristallina d’aspetto vitreo, liscio, ed è sempre perforato.<br />
Circondante il mamellone è la piattaforma che può essere crenulata da<br />
scrobicoli o liscia. Questa a sua volta è composta di solida calcite vitrea.<br />
Mamellone e piattaforma sono posti sulla cima di un rilievo chiamato<br />
bottone. È qui che s’inserisce l’apparato di ritenzione del radiolo.<br />
Questo apparato è composto di uno speciale tipo di tessuto connettivo<br />
che può fissare saldamente il radiolo al tubercolo. Per ultima c’é una<br />
piattaforma di sottile stereoma che circonda il bottone che viene<br />
chiamata areola. L’anello esterno dei muscoli del radiolo si ancora a<br />
quest’ultima. In molti casi l’area d’attacco dei muscoli è più ampia e<br />
meglio sviluppata da un lato del tubercolo; in questo caso è in questa<br />
direzione che è prevalentemente mosso il radiolo. I tubercoli più piccoli<br />
possiedono diversi un solo mamellone e bottone.<br />
Echi. 17
Echinidi a cuore con tubercoli eterogenei, come illustrato, sono caratteristicamente<br />
ristretti a specie viventi sia come epifauna sia come infauna<br />
di substrati piuttosto grossolani. Specie infossate in sabbia fine e<br />
fango possiedono sempre un rivestimento denso ed uniforme di tubercoli<br />
e spine aborali.<br />
Tubercoli portanti radioli che compiono diverse funzioni, di solito sono<br />
morfologicamente distinti. È peraltro possibile servirsi <strong>della</strong> loro struttura<br />
e distribuzione per comprendere meglio come gli echinidi a cuore<br />
utilizzino i loro radioli. Struttura e funzione dei radioli degli echinidi sono<br />
completamente documentate da A. B. Smith 1980a. The structure<br />
and arrangement of echinoid tubercles. Philosophical Transactions of<br />
the Royal Society, London, Series B, 289, 1-54.<br />
Da: http://www.nhm.ac.uk/research-curation/projects/echinoid-directory/morphology/hearts/tubercles.html
Fasciole<br />
Echi. 18<br />
Nella teca degli echinidi irregolari, le fasciole (una è indicata dalla freccia)<br />
sono aree con morfologia caratteristica densamente ricoperte da piccoli<br />
radioli specializzati. Questi radioli sono molto piccoli, fittamente impacchettati<br />
e posti nella parte terminale delle ghiandole del muco. Questi<br />
radioli sono anche fittamente cigliati lungo le loro aste. Nel vivente (sopra)<br />
queste aree risaltano per le polpose estremità dei radioli, ma la posizione<br />
delle fasciole è pure facilmente distinguibile nella parte scheletrica<br />
come continue zone di piccoli granuli strettamente addensati.<br />
Le fasciole si sono sviluppate principalmente negli echinidi fossatori<br />
perché utili per assolvere due importanti funzioni. Sono utili per la produzione<br />
del muco di rivestimento negli echinidi a cuore infaunali in quanto<br />
è richiesto specialmente nello scavo di sedimenti fini. Questo muco ricoprente<br />
viene emesso dalle fasciole e sparso sopra la superficie adorale<br />
dei radioli. Questo agisce da barriera impedendo alle particelle fini di penetrare<br />
fra i radioli, ostruendo così lo scavo. I piccoli radioli delle fasciole<br />
sono anche responsabili del movimento dell’acqua all’interno dello scavo<br />
ai fini respiratori. Le correnti generate dai radioli delle fasciole, densamente<br />
cigliati, spingono l’acqua dalla superficie all’interno dello scavo e<br />
nello stesso tempo la dirigono posteriormente all’animale all’interno del<br />
sedimento. La presenza delle fasciole negli esemplari fossili costituisce<br />
una chiara indicazione che l’animale è appartenuto all’infauna.<br />
La disposizione delle fasciole costituisce un importante carattere tassonomico.<br />
Sono mostrati di seguito i più importanti tipi di fasciole.<br />
Da: http://www.nhm.ac.uk/research-curation/projects/echinoid-directory/morphology/hearts/fasciole1.html
Fasciole peripetaloidi<br />
Sul lato aborale, attorno alla<br />
parte terminale dei petali<br />
Fasciole subanali<br />
Dietro il piastrone<br />
immediatamente sotto il<br />
periprocto. Sia allungate sia<br />
bilobate<br />
Tipi di fasciole<br />
Fasciole latero-anali<br />
Decorrenti dalla parte posteriore<br />
del petalo anteriore. Passano<br />
lateralmente dietro il periprocto.<br />
Spesso associate ad un elemento<br />
peripetaloide secondario<br />
Fasciole laterali<br />
Passanti attorno l’ambito e sotto<br />
l’ano ad una certa distanza dalle<br />
punte dei petali<br />
Fasciole interne<br />
Avvolgenti l’ambulacro anteriore<br />
ma non i petali pari<br />
Fasciole pseudolaterali<br />
In posizione ambitale ma<br />
passanti sopra il periprocto;<br />
quindi simili alle fasciole<br />
peripetaloidi<br />
Da: http://www.nhm.ac.uk/research-curation/projects/echinoid-directory/morphology/hearts/fasciole2.html<br />
Echi. 19
Radioli degli irregolari<br />
Echi. 20<br />
Gli echinidi a cuore sono fittamente coperti da un denso rivestimento<br />
di radioli filiformi dalla caratteristica punta “a spatola”. Questi radioli<br />
assolvono un numero di funzioni diverse. Sulla faccia aborale essi<br />
aiutano a compattare le pareti dello scavo, mantenendo uno spazio<br />
libero dal sedimento attorno all’animale. I radioli <strong>della</strong> superficie inferiore<br />
sono utilizzati per scavare ed avanzare. I radioli attorno al<br />
margine anteriore sono utilizzati per rimuovere i sedimenti come l’animale<br />
si sposta in avanti. I petali sono sovrastati da un arco di radioli<br />
che crea uno spazio d’acqua per la respirazione. In molti taxa,<br />
piccoli radioli specializzati fittamente impacchettati, secernenti muco,<br />
formano distinte aree chiamate fasciole. Sulla teca spoglia, queste<br />
appaiono come linee fittamente tappezzate di granuli.<br />
Da: http://www.nhm.ac.uk/research-curation/projects/echinoid-directory/morphology/hearts/spines.html
Chiave ai principali cladi di Irregolari<br />
1a. Peristoma largo, centrale e circolare<br />
con tacche boccali. Ambulacri con paia di<br />
pori semplici, indifferenziati<br />
Holectypoida<br />
2a. Placchette di tutte le cinque aree<br />
interambulacrali <strong>della</strong> faccia orale simili.<br />
Neognathostomata – Vai a 3<br />
3a. Paia di pori singoli su ciascuna area<br />
ambulacrale sotto i petali. Le aree di pori<br />
s’espandono di solito vicino al peristoma.<br />
Cassiduloida e gruppo stelo<br />
Neognathostomata<br />
1b. Peristoma di solito piccolo sempre<br />
privo di tacche boccali. Ambulacri con<br />
pedicelli e paia di pori differenziati<br />
(formanti fillodi e/o petali)<br />
Vai a 2<br />
Echi. 22<br />
2b. Area interambulacrale posteriore sulla<br />
faccia orale differenziata in un piastrone.<br />
Atelostomata<br />
3a. Molte paia di pori su ciascuna placca<br />
ambulacrale sotto i petali. Le aree di pori<br />
non s’espandono mai vicino al peristoma.<br />
Clypeasteroida<br />
Da: http://www.nhm.ac.uk/research-curation/projects/echinoid-directory/taxa/pageExtension.jsp?id=584
Chiave per i principali gruppi di Irregolari<br />
1a. Peristoma relativamente ampio, centrale<br />
e circolare con tubercoli boccali. Ambulacri<br />
con semplici paia di pori passanti.<br />
Holectypoida<br />
2a. Placche uguali su tutte le cinque aree<br />
interambulacrali sulla faccia orale.<br />
Neognathostomata – Vai alla 3<br />
1b. Peristoma di solito piccolo e sempre<br />
mancante di tubercoli boccali. Ambulacri con<br />
pedicelli ambulacrali e paia di pori differenziati<br />
(formanti filladi e petali).<br />
Vai alla 2<br />
2b. Area ambulacrale posteriore sulla<br />
superficie orale differenziata in piastrone.<br />
Atelostomata<br />
3a. Singoli paia di pori su ciascuna area<br />
3a. Molte paia di pori in ciascuna placca<br />
ambulacrale al di sotto dei petali. Aree di pori<br />
ambulacrale al di sotto dei petali. Aree a pori<br />
solitamente espanse vicino al peristoma.<br />
mai espanse vicino al peristoma.<br />
Cassiduloida e stem group<br />
Clypeasteroida<br />
Neognathostomata<br />
Da: http://www.nhm.ac.uk/research-curation/projects/echinoid-directory/taxa/pageExtension.jsp?id=584 e siti correlati<br />
(trad. M. Gnoli)<br />
Echi. 22
Chiave ai principali gruppi dei Clypeasteroidi<br />
1a. Pori confinati ad una sola fila<br />
adriale. Sphaeridia non inclusi nella<br />
placca.<br />
Togocyamus (tardo «stem-group»<br />
clypeasteroide)<br />
2a. Strutture di attacco dei muscoli<br />
<strong>della</strong> lanterna all’interno <strong>della</strong> teca<br />
adiacenti al peristoma formati da paia<br />
d’auricole ambulacrali (vedi frecce).<br />
Sphaeridia appaiati in ciascun<br />
ambulacro. Clypeasteroida<br />
3a. Aree interambulacrali che terminano<br />
adapicalmente in una singola<br />
placca o in serie di placche.<br />
Laganiformi<br />
1a. Pori disposti come bande attraverso<br />
le placche così come lungo le<br />
suture. Sphaeridia incluse nelle<br />
placche.<br />
Vai alla 2<br />
2b. Struttura di attacco dei muscoli<br />
<strong>della</strong> lanterna singola (fusa), “a<br />
cucchiaio” posto nell’in<strong>terra</strong>dio.<br />
Sphaeridium singolo per ambulacro.<br />
Scutellina (vai a 3)<br />
3b. Le aree interambulacrali restano<br />
biseriali fino all’apice.<br />
Scutelliformi<br />
Da: http://www.nhm.ac.uk/research-curation/projects/echinoid-directory/taxa/taxon.jsp?id=732 e siti correlati (trad. M.<br />
Gnoli)<br />
Echi. 23
Holectypoida: biologia e storia<br />
geologica<br />
Gli holectypoidi costituiscono un clade estinto che mostra una mescolanza<br />
di caratteri primitivi e derivati. Essi possiedono una grande e<br />
potente lanterna con denti “a cuneo”, come gli echinidi regolari, ma il<br />
loro periprocto è posto all’esterno del disco apicale come negli echinidi<br />
irregolari. La disposizione dei radioli orali suggerisce che gli holectypoidi<br />
scavavano il sedimento radialmente spostandolo dalla loro parte<br />
inferiore e vivendo probabilmente infossati nel sedimento. La mancanza<br />
di pedicelli respiratorî aborali e l’intreccio piuttosto rado e grossolano<br />
dei radioli indicano, in ogni modo, che devono essere vissuti limitatamente<br />
all’interno di substrati grossolani. Sulla base di queste osservazioni<br />
sembra probabile che i taxa del Giurassico come Holectypus o<br />
Pygaster (figurato sopra) debbano esser vissuti infossati solo per periodi<br />
di tempo limitati, emergendo probabilmente durante la notte, per<br />
alimentarsi allo stesso modo degli echinidi regolari. Un certo numero di<br />
generi più derivati vivevano probabilmente sempre sulla superficie del<br />
sedimento. È possibile che abbiano ricoperto la loro teca di frammenti<br />
clastici prelevati dal fondo come fanno oggi gli echinidi regolari.<br />
Il gruppo comparve nel primo Giurassico e divenne comune ed importante<br />
come membro delle faune d’acqua bassa a partire dal Giurassico<br />
medio. Sviluppò molte linee filetiche specializzate per vivere nei mari<br />
del “chalk” nel tardo Cretaceo, con i rari rappresentanti dei bassi fondali<br />
del Cenomaniano. Si estinsero alla fine del Cretaceo.<br />
Da: http://www.nhm.ac.uk/research-curation/projects/echinoid-directory/taxa/extraInfo.jsp?id=35<br />
Echi. 23a
Morfologia scheletrica del «dollaro<br />
delle sabbie» e affini<br />
I «dollari delle sabbie» ed i loro parenti (clypeasteroidi, cassiduloidi)<br />
possiedono una teca a simmetria bilaterale composta di 10 doppie file<br />
di placche, cinque interambulacrali (numerate 1-5) e cinque<br />
ambulacrali (numerate I-V). Le placche ambulacrali sono perforate<br />
da un singolo o doppio poro per la fuoriuscita dei pedicelli ambulacrali.<br />
Tutti i cinque ambulacri sono di solito identici e sulla superficie<br />
aborale le paia di pori formano un distinto disegno a petali. La teca<br />
(corona) porta due aperture principali, il peristoma e il periprocto. Il<br />
peristoma ospita la bocca ed è posto nella superficie inferiore, nel<br />
centro o verso il bordo anteriore. Questo è identificabile come il punto<br />
verso il quale convergono tutti i cinque ambulacri; è solitamente piuttosto<br />
piccolo a contorno circolare o pentagonale. In vita una “griglia”<br />
di radioli copre il peristoma. Il periprocto ospita l’apertura anale in<br />
posizione variabile. In alcune specie il periprocto è posto sopra l’ambito<br />
e può essere alloggiato in un distinto canale, il solco anale. In<br />
altre si può aprire su un lato tronco verticale, mentre in molti clypeasteroidi<br />
si pone sulla superficie orale. È sempre circondato da plac-<br />
Echi. 24<br />
1
chette dell’interambulacro 5. All’apice <strong>della</strong> teca, al punto d’origine<br />
delle aree ambulacrali, giace il disco apicale. Questo è composto di un<br />
piccolo numero di placchette, che sono le prime a formarsi nella<br />
metamorfosi. È in quel punto che si aprono le aperture delle gonadi.<br />
La disposizione delle placche del disco apicale forma una parte importante<br />
di caratteri tassonomici. La parte apicale degli ambulacri si sviluppa<br />
variamente in petali, zone specializzate di paia di pori ingranditi<br />
dove fuoriescono i pedicelli respiratorî. La forma ed il relativo sviluppo<br />
dei petali è molto uniforme nel gruppo, sebbene lo schema dettagliato<br />
possa essere importante per una definizione dei vari generi. In<br />
molti gruppi gli ambulacri sono composti di placche semplici. In certi<br />
clypeasteroidi, comunque, i petali possono contenere alternanze di<br />
placche grandi e piccole. È anche importante se i pori ambulacrali dei<br />
petali sono singoli o doppi.<br />
La disposizione dei pori è molto variabile in questo gruppo. Nei cassiduloidi<br />
ci sono bande di pori allargati vicino alla bocca che formano i<br />
fillodi. I fillodi ospitano i pedicelli alimentari, e la loro dettagliata disposizione<br />
costituisce un importante carattere tassonomico. I clypeasteroidi<br />
differiscono dai cassidulidi nell’avere un gran numero di pedicelli<br />
perforanti ciascun’area ambulacrale. I pori associati a questi<br />
pedicelli possono essere regolarmente disposti o irregolarmente<br />
sparsi. La forma delle prime placche interambulacrali che circondano<br />
il peristoma è significativa. In alcuni cassidulidi queste si sviluppano<br />
come rilievi che possono innalzarsi notevolmente come strutture “a<br />
piolo”.<br />
I radioli sono sempre corti, tozzi e di struttura uniforme. Sono sempre<br />
cavi e privi di cortex. I tubercoli sono conseguentemente piccoli,<br />
addensati e spesso leggermente infossati. Esiste una differenza molto<br />
piccola nei radioli e nei tubercoli in confronto agli spatangidi. I<br />
tubercoli sono perforati e crenulati.<br />
Nella maggior parte dei «dollari delle sabbie» ci sono cinque lunghe<br />
depressioni che si irradiano dal peristoma, e questi possono dividersi<br />
molte volte a formare un intricato modello di canali alimentari. Questi<br />
sono tappezzati di pedicelli ambulacrali che sono utilizzati per trasportare<br />
il particellato alimentare alla bocca centrale. Un altra caratteristica<br />
saliente di alcuni “dollari...” è la presenza di perforazioni<br />
passanti allungate definite come lunule.<br />
I «dollari delle sabbie» possiedono una lanterna ed una placca peristomale.<br />
La lanterna è insolitamente molto appiattita e manca dei<br />
compassi. La placca peristomale si compone di un singolo processo<br />
interambulacrale, o un paio di processi ambulacrali. Cassiduloidi non<br />
possiedono da adulti ne lanterna ne placca peristomale, sebbene sia<br />
presente una lanterna nella prima ontogenesi che viene in seguito<br />
riassorbita.<br />
Nei clypeasteroidi le superfici inferiore e superiore del guscio sono<br />
spesso connesse da un’estesa serie di rinforzi e pilastri. Questi sono<br />
solamente elementi strutturali che rinforzano la teca.<br />
Da: http://www.nhm.ac.uk/research-curation/projects/echinoid-directory/morphology/dollars/intro.html<br />
2
Rinforzi interni<br />
Lo scheletro dei «dollari delle sabbie» è rinforzato da una serie<br />
di sostegni, pilastri o coste che corrono fra la superficie superiore<br />
e quella inferiore <strong>della</strong> teca. Il sistema più semplice comprende<br />
10 partizioni parietali interne orientate radialmente che si sviluppano<br />
nella parte periferica del guscio. Questa è la situazione che<br />
si vede nella maggior parte dei fibulariidi. Clypeaster (all’interno<br />
<strong>della</strong> superficie aborale illustrata sotto) possiede una più complessa<br />
rete di barre concentriche che si sviluppano intorno ai sottili<br />
pilastri centrali. È comunque nei piatti «dollari delle sabbie»<br />
(scutellidi) che sviluppano meglio questi tipi di rinforzi. Negli<br />
scutellidi la maggior parte del loro interno è riempita da sostegni<br />
e pilastri e solo la traccia spirale del sistema digerente ne rimane<br />
libera. Il sistema di rinforzo agisce irrobustendo il guscio e permette<br />
ai piatti e sottili «dollari delle sabbie» di approvvigionarsi<br />
di cibo in zone dove le onde potrebbero provocare seri danni alla<br />
teca.<br />
Sopra. Sezione trasversale del dollaro delle sabbie Mellita.<br />
Sotto. Superficie aborale interna di Clypeaster, da Mortensen<br />
(1948) A monograph of the Echinoidea. IV(2). Clypeasteroida. C.<br />
A. Reitzel, Copenhagen.<br />
Da: http://www.nhm.ac.uk/research-curation/projects/echinoid-directory/morphology/dollars/buttressing.html<br />
Echi. 25
Lanterna e placca peristomale<br />
Echi. 26<br />
1
Le più specializzate e modificate lanterne di tutti gli echinidi, si<br />
trovano nei «dollari delle sabbie». Sono composte di quasi tutti<br />
gli elementi degli echinidi regolari (mancano solo i compassi) ma<br />
in confronto a questi sono molto basse e larghe. Le emipiramidi<br />
(H) sono molto schiacciate, deltoidi nella forma e non hanno<br />
foramen magnum. I denti (T) in sezione trasversale sono fatti a<br />
losanga e fortemente inclinati. Le epifisi e le rotule ridotte a<br />
piccoli elementi discoidali, sono molto nascoste alla vista fra le<br />
emipiramidi (nella posizione indicata (R/E).<br />
La placca peristomale segna il punto d’ancoraggio dei muscoli<br />
<strong>della</strong> lanterna. Nei clypeasteroidi consiste in un anello di tozze<br />
proiezioni. Queste, nelle Scutellina (figura sopra - G), sorgono<br />
dalle singole placche interambulacrali, ma nelle Clypeasterina<br />
sono delle strutture ambulacrali appaiate. I Cassiduloidi mancano<br />
di lanterna, ed in questo modo non hanno placca peristomale<br />
da adulti.<br />
La lanterna non può fuoriuscire dalla bocca ed è solo impiegata<br />
per afferrare e sminuzzare il materiale preso attraverso il peristoma.<br />
Poiché non è mai espulsa l’animale non necessita di sacche<br />
faringee o tacche boccali.<br />
La lanterna illustrata è quella di Laganum ed è figurata dall’alto<br />
(vista interna) e di lato.<br />
Da: http://www.nhm.ac.uk/research-curation/projects/echinoid-directory/morphology/dollars/disc.html<br />
2
Lunule<br />
Echi. 27<br />
Una delle caratteristiche più salienti di molti «dollari delle sabbie» è la<br />
presenza di fessure allungate che attraversano completamente la teca,<br />
chiamate lunule. Queste possono svilupparsi in una o due posizioni, lungo<br />
la linea mediana degli ambulacri, o lungo la parte mediana dell’interambulacro<br />
posteriore tra peristoma e periprocto (lunula anale). Le lunule<br />
ambulacrali si possono formare sia per riassorbimento dello scheletro nei<br />
primi stadi di crescita, sia come intaccature marginali che rimangono incluse<br />
nel processo di crescita. La lunula anale si forma sempre per riassorbimento.<br />
La funzione delle lunule è doppia. Principalmente agiscono come<br />
canali di drenaggio <strong>della</strong> pressione, impedendo che l’animale fuoriesca dal<br />
sedimento per l’azione delle onde. Test idrodinamici hanno mostrato che<br />
“dollari” con lunule possono vivere in siti con un alto tasso d’idrodinamismo.<br />
Secondariamente agiscono come un setaccio grossolano del sedimento<br />
sovrastante per il suo particellato organico. I pedicelli ambulacrali<br />
(pa) si estendono all’interno <strong>della</strong> lunula e catturano il particellato alimentare<br />
più grosso che passa attraverso l’apertura, mentre le correnti<br />
generate dalle ciglia convogliano le particelle più piccole nei solchi alimentari<br />
(sa) che bordano l’apertura. Le lunule quindi sono anche impiegate<br />
per una più efficiente raccolta del particellato alimentare.<br />
Da: http://www.nhm.ac.uk/research-curation/projects/echinoid-directory/morphology/dollars/lunules.html
Canali alimentari Echi. 28<br />
Molti «dollari delle sabbie» possiedono solchi appiattiti che s’irradiano<br />
dalla bocca. Questi prendono il nome di canali alimentari (fg) e,<br />
come dice il nome, la loro funzione principale è il trasporto del particellato<br />
alimentare alla bocca. Di solito sono più larghi verso la periferia<br />
e diventano gradualmente più sottili verso la parte centrale. In<br />
questo modo i canali alimentari formano un esteso intreccio sulla<br />
superficie orale <strong>della</strong> teca. Nei «dollari delle sabbie» i canali alimentari<br />
passano sempre in prossimità delle aperture delle lunule. Lo<br />
schema dettagliato delle ramificazioni dei canali alimentari<br />
costituisce un importante carattere sistematico.<br />
I canali alimentari sono tappezzati di mozzi pedicelli e trasportano le<br />
particelle di cibo in due modi. Primo le particelle alimentari sono<br />
spinte verso la bocca lungo le correnti ciliari. In alternativa, le particelle<br />
fini possono essere catturate dai tozzi pedicelli e convogliate<br />
verso la bocca inglobate nel muco pedicellare che scorre lungo il pavimento.<br />
In questo modo i canali alimentari agiscono come nastri<br />
trasportatori del fine particellato organico che cade sulle parti periferiche<br />
<strong>della</strong> teca o catturato dai pedicelli.<br />
Dendraster, l’unico «dollaro delle sabbie» conosciuto come sospensivoro,<br />
elevandosi dal sedimento, è insolito nell’avere i canali alimentari<br />
che si estendono nella superficie aborale, e può così catturare le<br />
particelle di cibo da entrambe le superfici <strong>della</strong> teca.<br />
Da: http://www.nhm.ac.uk/research-curation/projects/echinoid-directory/morphology/dollars/grooves.html
Pedicelli - «dollaro»<br />
I «dollari delle sabbie» possiedono un’unica disposizione dei<br />
pedicelli e dei pori ambulacrali. Mentre gli altri echinidi possiedono<br />
un solo pedicello che sorge da ciascuna placca ambulacrale,<br />
nei «dollari…» ci sono decine o centinaia di pedicelli per<br />
placca. Questi sono molto piccoli e terminano in un disco succhiante<br />
appiccicaticcio. Non possiedono internamente delle ampolle<br />
individuali ma emergono direttamente dalle ramificazioni<br />
laterali del sistema acquifero radiale. La loro funzione principale<br />
è quella di raccogliere il cibo e la loro minuta dimensione<br />
permette una specializzazione nella cattura delle piccole particelle<br />
alimentari che aderiscono ai granuli di sabbia. Ci sono<br />
anche dei pedicelli a forma di foglia sulla superficie aborale<br />
specializzati per gli scambi gassosi, come negli altri echinidi irregolari.<br />
Questi emergono da delle paia di pori “a fessura” che<br />
formano i petali aborali. La fotografia sopra riportata mostra in<br />
dettaglio le spine (sp) e pedicelli (tf) del «dollaro delle sabbie»<br />
Encope. Tutti i pedicelli visibili appartengono ad una sola placca<br />
ambulacrale.<br />
Da: http://www.nhm.ac.uk/research-curation/projects/echinoid-directory/morphology/dollars/tubefeet.html<br />
Echi. 29
Disco apicale e Petali<br />
Il disco apicale si trova alla sommità del guscio nel punto dove si originano<br />
i petali. In quasi tutti i “dollari” viventi ed I loro parenti, il disco si<br />
compone di una singola e larga placca centrale che rappresenta la placca<br />
genitale 2, conosciuta anche come placca madreporica (madreporite).<br />
Questa è densamente perforata da minuti pori, che rappresentano le aperture<br />
del sistema vascolare acquifero, ed è anche perforata da cinque gonopori,<br />
posti in<strong>terra</strong>dialmente alla sommità delle aree ambulacrali. I gonopori<br />
costituiscono il passaggio dei prodotti genitali (uova e sperma)<br />
rilasciati nell’acqua. Una piccola placca oculare è posta alla sommità di<br />
ciascun ambulacro adiacente alla placca genitale 2, inoltre possiede una<br />
perforazione più piccola. La parte superiore del sistema acquifero radiale<br />
esce qui a formare il pedicello ambulacrale terminale. Il disco così fatto,<br />
con una sola placca genitale, è detto monobasale.<br />
In pochi cassidulidi ed in molti taxa fossili, il disco apicale è composto di<br />
quattro placche genitali, non una. In questi casi c’é un singolo gonoporo<br />
per ciascuna placca genitale. In ogni caso, la placca genitale 2 rimane la<br />
più larga. Questo disco prende il nome di tetrabasale.<br />
I petali sono delimitati da file allungate di paia di pori che in vita cono<br />
associati a peduncoli a forma di foglia specializzati per gli scambi gassosi.<br />
Nel “dollaro delle sabbie”, possono essere diritti o piegati, come sopra,<br />
e possono terminare gradualmente, con le paia di pori che diminuiscono<br />
di dimensione e crescono in spaziatura verso l’ambito, o improvvisamente<br />
(come sopra).<br />
Da: http://www.nhm.ac.uk/research-curation/projects/echinoid-directory/morphology/dollars/disc.html<br />
Echi. 30
Classificazione delle scutellomorfe<br />
Echi. 31<br />
Nella sua monografia, Mortensen (1948) incluse semplicemente tutte le scutelliformi<br />
in una sola famiglia, le Scutellidae. Durham (1955) incominciò il<br />
processo di classificazione di questo grande clade e stabilì numerose famiglie.<br />
Egli, comunque, non assegnò un ordine gerarchico per le dieci famiglie<br />
che propose, conseguentemente non fu proposta una classificazione formale.<br />
La più completa trattazione sistematica di questo gruppo rimase non pubblicata<br />
(Mooi, tesi di dottorato non pubblicata).<br />
Considerando le forme viventi, queste sembrano appartenere a due gruppi<br />
principali, le Echinarachnidae, caratterizzate dall’avere i canali alimentari che<br />
formano un tronco principale sulle prime placche ambulacrali dove si trova il<br />
sistema vascolare acquifero, e le forme restanti, che hanno i canali alimentari<br />
che si biforcano al termine del circolo basicoronale con i pedicelli ambulacrali<br />
ed i canali alimentari che si espandono attraversando le aree ambulacrali.<br />
Questi clade sorelle sono qui considerate a livello di superfamiglie: Echinarachnidea<br />
e Scutellidea.<br />
Scutellidea può essere ulteriormente divisa in un certo numero di famiglie<br />
delle quali Mellitidae e Astriclypeidae sono taxa sorelle, unite dall’avere un<br />
sistema interno di microcanali, radioli e tubercoli sulla superficie orale, chiaramente<br />
diversi in aree di approvvigionamento alimentare e locomotoria;<br />
inoltre mostrano l’apertura del periprocto nella prima placca interambulacrale<br />
basicoronale. Le Mellitidae costituiscono il gruppo più derivato possedendo<br />
una lunula anale. Sebbene entrambe i clade possiedano tacche ambulacrali o<br />
lunule, queste sono veramente fatte in modo diverso, come notato da Seilacher<br />
(1979) e sono presumibilmente corrispondenti per evoluzione convergente<br />
o parallela.<br />
Il gruppo sorella vivente delle Mellitidae più Astriclypeidae e costituito dalle<br />
Dendrasteridae, un gruppo di «dollari delle sabbie» mancante di lunule e con<br />
un sistema meno sviluppato di rinforzi interni con canali in<strong>terra</strong>diali a forma<br />
di “V” e un più aperto intreccio radiale nelle aree ambulacrali. Le Dendrasteridae<br />
hanno il periprocto molto marginale che si apre delle placche posteriori<br />
alla prima placca post-basicoronale. Principali ramificazioni dei canali alimentari<br />
che si estendono nel centro delle file interambulacrali verso la periferia.<br />
La posizione dei taxa fossili rimane problematica, ma le Protoscutellidae, col<br />
loro semplice canale alimentare che si biforca solo dopo il secondo o terzo<br />
paio di placche ambulacrali, si pone tra le Echinarachnidea e Scutellidea. Essi<br />
non possono, in ogni modo, essere i diretti progenitori delle Scutellidea, possedendo<br />
cinque gonopori, una condizione derivata. Iheringiella è a volte insolita<br />
per avere i canali alimentari che si biforcano alla fine del secondo paio di<br />
ambulacri. Le diramazioni del tronco principale formano dei tratti rilevati sulla<br />
teca piuttosto che dei solchi, come nelle Echinarachniidae.<br />
Durham (1955) riconobbe che le Scutellidae contengono una mescolanza di<br />
1
forme primitive estinte, ma Scutella e Parascutella mostrano dei rinforzi interni<br />
ed una disposizione dei canali alimentari delle Dendrasteridae, ma conservano<br />
una condizione primitiva possedendo aree ambulacrali continue.<br />
Abertella ed il suo strettissimo parente Parmulechinus, formano un gruppo<br />
sorella del clade (Astriclypeidae+Mellitidae), possedendo il denso reticolato<br />
interno di quel clade, ma l’apertura del periprocto in una placca interambulacrale<br />
più arretrata ed una forte differenzia nelle tubercolazioni orali.<br />
Le monophorasteride rappresentano un altro clade completamente estinto.<br />
Queste possiedono una lunula anale e di solito sono considerate come un<br />
«gruppo sorella» delle mellitide. Inoltre, similmente a quest’ultime, il loro<br />
periprocto giace associato al primo paio di placche basicoronali interambulacrali.<br />
Contro questo, in ogni modo, esse possiedono aree intrerambulacrali<br />
molti sottili e né pori accessori né canali alimentari che si estendono attraverso<br />
le aree ambulacrali.<br />
Scutaster possiede lunule “a festoni”, come le Mellitidae, ma manca d’una<br />
lunula anale ed ha le aree interambulacrali molto sottili. Fino a quando non<br />
sarà ridescritto su materiale migliore, è lasciato come incertae sedis nelle<br />
Scutellidea.<br />
Le scutellimorfe neoteniche costituiscono un grosso problema, essendo molto<br />
difficile distinguerle dalle piccole laganimorfe. Mooi (1990) and Mooi &<br />
Chen (1996) hanno fornito un gran contributo per chiarire la posizione sistematica<br />
di alcune di queste piccole forme. In particolare, la presenza di una<br />
pesante fascia diverticolare fornirebbe un eccellente carattere per separare le<br />
piccole scutellimorfe neoteniche dalle laganimorfe.<br />
La classificazione qui adottata è la seguente (*= estinte):<br />
Infraordine Scutelliformi Haekel, 1896<br />
Famiglia Echinarachniidae Lambert, 1914<br />
*Famiglia Protoscutellidae Durham, 1955<br />
*Genere Iheringiella Berg, 1898<br />
Famiglia Scutellidae Gray, 1825<br />
Famiglia Astriclypeidae Stefanin, 1911<br />
* Famiglia Monophorasteridae Lahille, 1896<br />
Famiglia Mellitidae Stefanini, 1911<br />
La lista e la chiave dettagliata dei generi di Scutelliformi si può trovare in<br />
Mooi (1989).<br />
_______________________________________________________________________________________<br />
Mortensen, T. 1948. A Monograph of the Echinoidea IV.2 Clypeasteroida. C. A. Reitzel,<br />
Copenhagen.<br />
J. W. Durham 1955. Classification of clypeasteroid echinoids. University of California<br />
Publications in Geological Sciences 31(4), 73-198.<br />
2
Mooi, R. 1989. Living and fossil genera of the Clypeasteroida (Echinoidea:<br />
Echinodermata): an illustrated key and annotated checklist. Smithsonian Contributions<br />
to Zoology 488, 1-51.<br />
Mooi, R. 1990. Progenetic minaturization in the sand dollar Sinaechinocyamus:<br />
implications for clypeasteroid phylogeny. Pp. 137-143 in C. de Ridder et al. (eds)<br />
Echinoderm Research: Proceedings of the Second European Conference on<br />
Echinoderms Rotterdam: A. A. Balkema.<br />
Mooi, R. & Chen, C.-P. 1996. Weight belts, diverticula, and the phylogeny of the sand<br />
dollars. Bulletins of Marine Science 58, 186-195.<br />
Seilacher, A. 1979. Constructional morphology of sand dollars. Paleobiology 5, 191-<br />
221.<br />
Da: http://www.nhm.ac.uk/research-curation/projects/echinoid-directory/taxa/extraInfo.jsp?id=28<br />
3
Principali caratteri scheletrici che supportano il cladogramma [taxa attuali in grassetto]:<br />
1. Cresta periradiale sviluppata sulle placche basicoronali; partizioni interne di barre non radiali ed a stella.<br />
2. Tronco dei canali alimentari primari lungo che si triforca o irregolarmente strombato distalmente.<br />
3. Canali alimentari biforcati.<br />
4. Canali alimentari curvi verso l’esterno, con biforcazioni secondarie che sorgono nel lato interno.<br />
5. Canali alimentari che si biforcano due volte dando origine ad un canale sopra il centro di ogni fila di<br />
placche (sistema vascolare acquifero si espande sulle aree interambulacrali). Tronco dei canali alimentari<br />
primari formato da una vera doccia piuttosto che una porzione sollevata di pori densi.<br />
6. Cinque gonopori.<br />
7. Canali alimentari che si biforcano alla fine distale del cerchietto basicoronale.<br />
8. Periprocto a metà strada tra margine e peristoma, che si apre nel primo paio delle placche postbasicoronali<br />
interambulacrali.<br />
9. Interambulacri disgiunti sulla superficie orale, ma prime placche ambulacrali post-basicoronali non<br />
significativamente ingrandite.<br />
10. Canali alimentari che si estendono sopra la superficie aborale attorno la parte posteriore del guscio.<br />
11. Guscio a margini sottili; ampia fessura posteriore e fessure ambulacrali posteriori; rinforzi interni<br />
molto fitti formanti una struttura a nido d’ape.<br />
12. Interambulacri disgiunti sulla superficie orale, con seconde aree ambulacrali ingrandite.<br />
13. Tubercoli sulla faccia orale diversificati in aree locomotorie ed aree per la selezione trofica (corrispondenti<br />
ad una diversificazione funzionale dei radioli); pori accessori assenti (o estremamente radi) sulla<br />
superficie aborale; periprocto che si apre nelle prime placche post-basicoronali interambulacrali.<br />
14. Lunule ambulacrali o fessure di tipo trasversale; canali alimentari primari privi di grossi pori.<br />
15. Periprocto posto in una profonda depressione anale o lunula.<br />
16. Lunula anale completamente formata.<br />
17. Fessure ambulacrali o lunule “a festone”; aree ambulacrali di dimensioni simili a quelle interambulacrali<br />
all’ambito.<br />
4
1a. No lunule a qualsiasi dimensione.<br />
Chiave ai principali gruppi di Scutellidea<br />
Vai alla 5<br />
2a. Lunula anale sempre presente<br />
nell’interambulacro posteriore.<br />
Vai alla 3<br />
3a. Canali acquiferi radiali e pedicelli associati che si<br />
estendono lontani sulle aree ambulacrali; aree<br />
interambulacrali agli ambiti di rado meno del 50%<br />
dell’ampiezza delle aree ambulacrali.<br />
Mellitidae<br />
4a. Lunule o fessure “a festone”; assenti dagli interambulacri<br />
posteriori. Aree interambulacrali all’ambito<br />
solo di ampiezza di circa il 20% di quelle ambulacrali.<br />
Scutasteridae<br />
1b. Aperture o lunule presenti da dimensioni piccole.<br />
Vai alla 2<br />
2b. Lunula anale assente, solo lunule ambulacrali o<br />
aperture.<br />
Vai alla 4<br />
3b. Canali alimentari e pori per I pedicelli confinati alle<br />
aree ambulacrali; Aree interambulacrali che si assottigliano<br />
verso l’ambito, dove sono meno del 20%<br />
dell’ampiezza delle aree interambulacrali.<br />
Monophorasteridae<br />
4b. Lunule o fessure di tipo trasversale; sempre<br />
presenti negli interambulacri posteriori. Aree interambulacrali<br />
all’ambito grandi come quelle ambulacrali.<br />
Astriclypeidae<br />
Da: http://www.nhm.ac.uk/research-curation/projects/echinoid-directory/taxa/key.jsp?id=266<br />
Echi. 31a
Infraordine Scutelliformi Haekel,<br />
1896<br />
Diagnosi<br />
Clypeasteroidi con:<br />
• Struttura d’attacco <strong>della</strong> lanterna composta di<br />
muscoli congiunti a cinque “pioli” in<strong>terra</strong>diali<br />
fusi<br />
• Disposizione delle placche interambulacrali<br />
all’apice<br />
Distribuzione primo Eocene all’Attuale; cosmopolita<br />
Da: http://www.nhm.ac.uk/research-curation/projects/echinoid-directory/taxa/taxon.jsp?id=798<br />
Echi. 32
Classe CRINOIDEA<br />
In figura alcune forme di crinoidi attuali (A-D), tutti Articulata.<br />
Come si è visto nel lucido introduttivo al Phylum Echinodermata, i crinoidi<br />
costituiscono una Classe di Pelmatozoi, Subphylum Crinozoa i cui primi rappresentanti<br />
risalgono sicuramente all’Ordoviciano inf. e secondo Simms<br />
(1993) al Cambriano medio (Echmatocrinus del Burgess Shale); e sono ancor<br />
oggi presenti con un numero ridotto di forme, la cui più nota è forse la Comatula,<br />
D (= Antedon). Questa vive nell’ambiente marino libera, senza peduncolo<br />
nelle prime fasi di vita, per passare da adulto ad un modo di vita epipelagico<br />
ancorata a tronchi o rami galleggianti. Altre forme attuali, illustrate sopra, sono:<br />
A - Teliocrinus; B - Isocrinus e C - Ptilocrinus.<br />
Come si vedrà nel capitolo riguardante la tassonomia di questi organismi, i<br />
rappresentanti attuali costituiscono una minima parte delle numerose forme<br />
fossili che sono state descritte (c.a. 6000).<br />
1
Fig. 1 e 2<br />
La morfologia generale di un crinoide provvisto di peduncolo per il suo fissaggio,<br />
come parzialmente illustrato in figura, è formata da un sistema d’ancoraggio,<br />
un peduncolo, un calice o teca formato da un insieme di placche<br />
coronali, disposte secondo due principali modelli, dalle<br />
quali si innalzano le braccia che servono prevalentemente<br />
per la cattura del particellato alimentare per il sostentamento<br />
dell’animale. Si preferisce, in ogni modo, descrivere le<br />
varie parti qui di seguito, con maggior dettaglio.<br />
SISTEMA D’ANCORAGGIO -<br />
Fig. 3<br />
In funzione del modo di vita adottato dall’organismo, il fissaggio dell’animale<br />
può essere al fondo marino, adottando accorgimenti anatomici come quello<br />
mostrato, oppure a particolari “elementi galleggianti”, come nel caso del gen.<br />
pelagico del primo Devoniano Scyphocrinites (Camerata) che pur possedendo<br />
un lungo peduncolo era fissato ad un’unità pneumatica vuota assemblata con<br />
placchette di calcite, conosciuta come “lobolite”. Altre forme sempre pelagiche,<br />
durante il Giurassico, si ancoravano a tronchi galleggianti come mostrano<br />
degli esemplari, di circa 4 m di lunghezza, rinvenuti nel calcare litografico di<br />
Solnhofen in Baviera.<br />
2
PEDUNCOLO -<br />
Questo elemento, come tutte le altre parti di un crinoide, è composto di dischetti<br />
di calcite, circolari o subpoligonali interconnessi fra loro in modo<br />
elastico, in modo da permettere il movimento dell’intera “colonna”. Questi<br />
dischetti detti “entrochi”, sono perforati centralmente creando un “lumen”<br />
centrale lungo tutta la struttura. Questa cavità ospita canali e nervi del celoma<br />
e gli organi assiali, che fra l’altro, sono interessati ai movimenti dell’animale.<br />
In figura i caratteri del peduncolo – A e B, E articoli di forme attuali, i rimanenti mostrano<br />
forme fossili. In D tipi di faccette articolari di varie forme mostranti il foro centrale assiale<br />
(“lumen”); in A e C alcune segmenti “nodali” con incavi articolari per l’inserzione dei cirri.<br />
CALICE o TECA -<br />
Il calice rappresenta la parte principale del crinoide e consiste di un calice<br />
dorsale e tegmen. Il calice o teca è un corpo provvisto di placche a protezione<br />
delle diverse cavità celomatiche e di un sistema vascolare acquifero aperto.<br />
Il calice dorsale è formato di placche poligonali (solitamente pentamere)<br />
lateralmente connesse fra il tegmen ed il peduncolo. Due sono i tipi principali<br />
di calice: il monociclico ed il diciclico. Il calice monociclico si compone di una<br />
sola serie di placche dalle quali si estendono le braccia. Il calice diciclico, come<br />
si può dedurre, si compone di due file di placche sotto quelle che portano<br />
le braccia.<br />
CALICE MONOCICLICO<br />
Questo è il diagramma “esploso” e<br />
svolto di un calice monociclico di<br />
crinoide. Le placche <strong>della</strong> teca<br />
portanti le braccia, possiedono<br />
delle tacche chiamate "faccette<br />
brachiali" dove s’inseriscono le<br />
braccia. Queste placche sono det-<br />
3
te "radiali", la serie di placche più piccole sopra le radiali sono le "orali", e<br />
quelle sotto le "basali". Le placche orali con la serie di placchette triangolari<br />
costituiscono la piramide anale. Le due placche ombreggiate fanno parte di<br />
una serie di placche anali che sostengono la piramide anale, e sono note come<br />
l’«anale X» (ombreggiata chiaro), e la «radiale anale» (ombreggiata scuro).<br />
CALICE DICICLICO<br />
Piramide anale<br />
Questo è il diagramma<br />
“esploso” e svolto<br />
di un calice diciclico.<br />
Le placche che<br />
portano le braccia<br />
presentano delle tacche<br />
chiamate "faccette<br />
brachiali" dalle<br />
quali si estendono le<br />
braccia. Queste placche sono note come "radiali", le placche più piccole sopra<br />
le radiali sono le "orali", e quelle sotto sono le "basali". La fila di placche sul<br />
fondo sono chiamate "infrabasali". Le placche orali con la serie di placchette<br />
triangolari formano la piramide anale. Le tre placche ombreggiate sono parte di<br />
una serie di placche anali sostenenti la piramide anale, e sono note come<br />
l’«anale X» (ombreggiata scuro), e le «radiali anali divise» (le due grigie).<br />
TEGMEN -<br />
Come illustrato nella fig. 2, questo è posto superiormente il calice dorsale e si<br />
compone <strong>della</strong> piramide anale sommitale, circondata da numerose placchette<br />
che proteggono la parte terminale del tubo digerente.<br />
I crinoidi furono e sono animali che si nutrono, come tali necessitano di bocca<br />
ed ano. Come la maggior parte degli echinodermi, i crinoidi possiedono un<br />
tratto digestivo a “senso unico” con una bocca, una sorta di sistema intestinale,<br />
e un ano. Sfortunatamente possiedono bocca ed ano affiancati all’interno<br />
del tegmen; di conseguenza l’apparato digerente ha forma di una “U”.<br />
L’apertura anale è protetta dalla piramide anale, una sporgenza ripiegata che si<br />
protrude dal tegmen. Questo è posto nell’area dorsale <strong>della</strong> teca circondata<br />
dalle braccia per l'alimentazione che si innalzano verso l’alto. La bocca è posta<br />
strategicamente alla base delle braccia che utilizzano i loro solchi ambulacrali<br />
cigliati per trasportare il particellato alimentare raccolto verso il basso all’interno<br />
<strong>della</strong> bocca. Durante questo processo, le placchette di ricoprimento a<br />
protezione del tegmen ed i numerosi pedicelli ambulacrali si estendono verso<br />
l’alto permettendo al cibo d’entrare nella bocca.<br />
4
BRACCIA -<br />
Le braccia dei crinoidi sono delle strutture d’alimentazione specializzate per<br />
catturare il particellato alimentare in sospensione, flottante attorno l’animale.<br />
Esse sono costituite da un insieme di segmentti i brrachi ial li i, posti uno sull’altro a<br />
formante un lungo braccio composto di pinnule, solchi ambulacrali, pedi icel ll li i<br />
ambul lacrral li i e pl lacchetttte di i rri icoprri imentto. (Fig. 4 e 5)<br />
Nell’immagine a sinistra: particolare di un braccio<br />
di crinoide attuale, mostrante il sol s lco al li imenttarre<br />
che si dirama alle pi innul le e dirige il particellato<br />
alimentare, catturato dai pedi icel ll li i, come un<br />
«nastro trasportatore» all’interno <strong>della</strong> bocca.<br />
(Da C. Messing, 1987)<br />
FILOGENESI e TASSONOMIA<br />
I crinoidi la cui teca è costituita da quattro serie di placche (Disparida, Cladida,<br />
e Camerata) compaiono tutti nel primo Ordoviciano; i Flexibilia nell’Ordoviciano<br />
medio, e gli Articulata si<br />
evolsero nel primo Mesozoico.<br />
Camerati, disparidi,<br />
flessibili, e cladidi si estinsero<br />
alla fine dell’era Paleozoica,<br />
circa 225 milioni<br />
d’anni fa. Gli Articulata sono<br />
gli unici crinoidi post-<br />
Paleozoico. La loro evoluzione<br />
è da mettere in relazione<br />
all’estinzione di massa<br />
del Permo-Triassico o<br />
subito dopo.<br />
5
Le principali tappe evolutive nella suddetta filogenesi dei crinoidi riguardano<br />
(vedi l’albero filogenetico di figura): i Crinoidi divergono dagli echinodermi<br />
rombiferidi per (1) la mancanza di rombi porali, gonoporo e brachiole biseriali;<br />
lo sviluppo di vere braccia con estensione degli ambulacri; lo spostamento<br />
dell’ano al tegmen; l’aggiunta di placche anali e una migliore simmetria pentamera.<br />
I crinoidi primitivi che ne risultarono furono forme con un calice a quattro<br />
serie di placche, che costituirono un “gruppo base” a bassa diversità<br />
(Ausich, 1998). Da questa costruzione del calice a quattro serie di placche, si<br />
generarono i Disparida per perdita <strong>della</strong> serie di placche basali (2). La linea<br />
filetica dei Cladida, si generò per la perdita <strong>della</strong> serie di “placche superiori” (3)<br />
e dette luogo all’inizio tre nuove linee filetiche di gran successo: i Camerata (4)<br />
con brachiali e in<strong>terra</strong>diali fisse incluse nel calice, disposizione delle placche<br />
posteriori simmetriche e sutura delle placche rigida; i Flexibilia (5) con la<br />
bocca esposta sul tegmen e sutura delle placche allentata e gli Articulata (6)<br />
per la perdita <strong>della</strong> placca anale ed un sistema endoneurale incluso nelle placche<br />
del calice. Il carattere monofiletico <strong>della</strong> sottoclasse Articulata è stato dedotto<br />
e sostenuto da Simms & Sevastopulo (1993).<br />
SISTEMATICA dei CRINOIDI<br />
I Crinoidi, con più di 6000 specie descritte, hanno costituito il gruppo dominante<br />
degli echinodermi nel Paleozoico. Ora il gruppo è rappresentato da sole<br />
600 specie. Questo esiguo numero di rappresentanti viventi complica il lavoro<br />
di ricostruire la filogenesi dei crinoidi. Il taxon Crinoidea fu stabilito nel 1821 da<br />
J.S. Miller ponendo tutti i crinoidi peduncolati fuori dal gruppo degli Stellaroidea<br />
(stelle di mare). I passi successivi per risolvere la storia dei crinoidi si<br />
basarono tutti sulla disposizione delle placche <strong>della</strong> teca nei confronti <strong>della</strong><br />
posizione delle braccia. Tradizionalmente, le placche <strong>della</strong> teca furono descritte<br />
sia come monocicliche sia come dicicliche, con la prima serie portante le<br />
braccia come “orali”, la serie immediatamente sottostante come “radiali” e sotto<br />
ancora le “basali”. Per la disposizione diciclica, la serie più bassa è costituita<br />
dalle “infrabasali”. La nuova interpretazione di Simms (1993) sull’omologia<br />
delle placche coronali dei crinoidi, che si basa sui loro rapporti rispetto al peduncolo<br />
e non rispetto alle braccia, è sostenuta dall’ontogenesi e dalla posizione<br />
delle altre placche nella teca. Conseguentemente gli alberi filogenetici<br />
proposti sono questi:<br />
6
Articulata (Triassico inf. - Attuale)<br />
Tutti i crinoidi viventi sono degli Articulata, originatisi nel Triassico, costituiscono<br />
i soli crinoidi apparsi dopo il Paleozoico. La maggior parte è monociclica<br />
avendo perso una serie di placche coronali. Tradizionalmente si assume<br />
che quelle perse siano le infrabasali, ma secondo Simms qualsiasi fra le serie<br />
delle basali o infrabasali potrebbe essere stato abbandonato. Basandosi sui<br />
cambiamenti nell’orientazione fra le braccia ed il peduncolo Simms dimostra<br />
che la più probabile fra le serie di placche persa sia quella delle radiali, e non<br />
quella delle infrabasali. Questa interpretazione suggerisce che gli Articulata,<br />
fra i Cladida, Flexibilia e Cyathocrina sono tutti derivati dal gruppo d’Aethocrinus<br />
per la perdita di una delle serie di placche <strong>della</strong> teca.<br />
1 3<br />
2<br />
Immagini di alcuni Articulata: 1) Antedon medi<strong>terra</strong>nea (Attuale); 2) Saccocoma pe-<br />
ctinata (Giurassico, Holtzmaden); 3) Pentacrinites (Giurassico, Europa)<br />
Flexibilia (Ordoviciano medio - Permiano sup.)<br />
Questi crinoidi fanno parte dei cosiddetti "Cladidi", compaiono nell’Ordoviciano<br />
e come la maggior parte degli altri crinoidi non sopravissero all’estinzione<br />
Permo-Trias. Morfologicamente i Flexibilia presentano un calice diciclico conico<br />
con placche articolari allentate; braccia uniseriali senza pinnule; un tegmen<br />
flessibile con molte piccole placche ed un peduncolo circolare privo di cirri.<br />
7
Nell’immagine a destra, il<br />
classico rappresentante dei<br />
Flexibilia: il gen. Taxocrinus<br />
Camerata (Ord. inf. - Permiano sup.)<br />
I Camerati costituiscono un gran gruppo di crinoidi Paleozoici rientrante nel<br />
clade Disparida che comprende circa 210 generi. Si sono originati nel primo<br />
Ordoviciano e si estinsero dopo l’estinzione di massa del Permo-Triassico.<br />
Possiedono un calice sia monociclico sia diciclico. I Camerata sono caratterizzati<br />
dall’avere molte placche formanti il calice, da un tegmen dove le<br />
grosse placche si dispongono rigidamente, brachiali ed interbrachiali fisse;<br />
dalla mancanza d’idroporo (nelle placche orali) e di braccia con pinnule.<br />
Nella fig. a destra, due dei<br />
generi più rappresentativi<br />
<strong>della</strong> sottoclasse:<br />
Paradichtocrinus (Permiano<br />
inf., U.S.A.) e tegmen;<br />
Scyphocrinites (Devoniano<br />
inf., Gondwana) col suo<br />
organo d’ancoraggio e/o<br />
galleggiamento (lobolite).<br />
Aethocrinus<br />
Si è discusso su quello che fosse il crinoide più antico; alcuni sostengono che<br />
Echmatocrinus del Cambiano medio del Burgess Shale rappresenti il crinoide<br />
più vecchio, mentre altri discutono le affinità coi crinoidi d’Echmatocrinus e<br />
considerano come tale Aethocrinus dell’Ordoviciano inferiore. Simms (1993)<br />
considera Aethocrinus come il più antico gruppo di crinoidi e suggerisce che<br />
rappresenti il «sister group» di tutti gli altri. Simms afferma ciò sull’evidenza<br />
suggerita dall’organizzazione delle placche in quattro serie sia primitiva, che<br />
8
Aethocrinus appaia molto prima nella documentazione fossile, e che possieda<br />
altre caratteristiche primitive.<br />
Cyathocrinina<br />
I Cyathocrinina sono caratterizzati dall’avere calice diciclico ed una placca radiale<br />
anale molto simile a quella dei rimanenti Cladidi. La caratteristica più diagnostica<br />
dei primi Cyathocrinina è la presenza di una fenditura endotecale nelle<br />
placche del calice, la fenditura endotecale, in ogni modo, è stata persa nella<br />
maggior parte delle forme più<br />
recenti. In figura: Cyathocrinites<br />
Perittocrinidae<br />
I Perittocrinidi rappresentano un<br />
gruppo a calice monociclico che<br />
prese origine nel primo Ordoviciano. Questi sono gli unici, fra i disparidi, ad<br />
avere una fenditura endotecale nelle placche del calice. L’analisi filogenetica<br />
dei Perittocrinidi di Simms suggerisce che questi rappresentino il «sister<br />
group» primitivo dei taxa Hybocrinida e Disparida.<br />
Hybocrinida<br />
Gli Hybocrinida fanno parte del clade dei Disparida e morfologicamente un<br />
gruppo disparide. Sono stratigraficamente esclusivi dell’Ordoviciano-Siluriano<br />
inferiore. Possiedono calice monociclico, una placca aggiuntiva adiacente alla<br />
placca anale e peduncolo circolare.<br />
Disparida<br />
Nell’analisi filogenetica dei Disparida, Simms impiega il termine “Disparidi” per<br />
riferirsi al clade che include Camerata, Perittocrinidae, Hybocrinida e Disparida.<br />
Una descrizione del taxon Disparida sembra mancare, è più probabile l’uso<br />
del taxon Cladida. [Vedi il primo dei due “alberi” figurati (da Ausich, W. I., 1999)]<br />
In figura: A e B rappresentazione<br />
schematica di calice diciclico A) e<br />
monociclico B) presentati secondo<br />
l’omologia tradizionale delle placche;<br />
in C) secondo l’omologia delle placche<br />
rivista (Simms, 1993), dove la serie di<br />
placche <strong>della</strong> teca perdute sarebbero<br />
le “radiali”. Le figure mostrano il<br />
calice in veduta peduncolare)<br />
9
Si riportano di seguito alcune pagine che illustrano crinoidi pelagici fossili provenienti dal<br />
Marocco e dalla Cina<br />
Si estinguono i crinoidi pelagici<br />
Procedendo nell’illustrazione <strong>della</strong> mostra, all’altezza <strong>della</strong> galleria dei fossili al di<br />
sopra di tutte le innumerevoli forme mostrate si è aggiunta una vetrina di nuovo<br />
allestimento dove si possono vedere i crinoidi pelagici (immagine sotto).<br />
Le due lastre a crinoidi più grandi rappresentano le più belle e più preziose che sono<br />
mostrate nel museo. La grande lastra proveniente dal Marocco è stata sovvenzionata<br />
dalla libera associazione di condiscendenza del Museo, i pezzi rimanenti sono un<br />
dono degli Amici del Museo di Storia Naturale.<br />
I “Gigli di mare” non sono piante, ma una classe d’animali Pelmatozoi di un ceppo al<br />
quale appartengono anche i ricci e le stelle marine, gli <strong>Echinodermi</strong>. Essi catturano le<br />
microscopiche particelle di cibo, flottante nel mare, con le braccia che si estroflettono<br />
dai loro calici. I crinoidi sono noti nella Storia <strong>della</strong> <strong>terra</strong> per 500 milioni d’anni ed esistono<br />
tuttora sotto la forma di “comatule” libere natanti, che sono ben conosciute in<br />
prossimità delle barriere coralline. In passato la maggior parte dei crinoidi era ancorata<br />
al fondo del mare con un peduncolo. Queste forme si trovano oggi solo nei mari più<br />
profondi.<br />
Crinoidi galleggianti dal Marocco:<br />
10
I crinoidi più grossi e più belli non vivevano ancorati ai fondali ma abitavano i mari<br />
aperti. Un gruppo costituito dagli Scyphocrinites fu rigoglioso nel passato geologico,<br />
circa 410 milioni d’anni fa. I ritrovamenti più belli provengono dal Marocco. Infatti, i nativi<br />
costituiscono probabilmente i visitatori più diffusi delle oasi dei fiumi sud-orientali<br />
del Tafilalat ben noto con Erfoud. Le lastre con i crinoidi sono qui raccolte dai nativi<br />
dal duro suolo desertico, nel quale scavano profonde e lunghe gallerie con un laborioso<br />
lavoro manuale per trovarle. Le lastre rotte devono essere unite e preparate per<br />
raggiungere il loro indiscutibile splendore (fotografia sopra). Fra le corone ed i calici ci<br />
sono dei corpi fatti a sfera del diametro di 10 cm o più. Questi “palloncini”, detti<br />
Loboliti, sono mostrati nella vetrina sottostante (immagine sotto a sinistra). La ricostruzione<br />
mostra il lobolite galleggiante riempito di gas, dal quale pende il peduncolo<br />
al quale è connesso il calice (immagine destra in basso). I crinoidi galleggianti del<br />
Marocco appartengono all’evoluzione più strana dei crinoidi in senso assoluto.<br />
Quanto potesse galleggiare un simile ancoraggio, è ancora oggetto di ricerca.<br />
Il secondo gruppo di crinoidi pelagici è costituito da forme che si ancoravano a legni<br />
flottanti. Già da molto tempo sono noti nell’area dei resti lignei vicino a Stoccarda, dove<br />
si trovano su lastre di scisto nero. Si possono vedere nelle nuove lastre <strong>della</strong> vetrina<br />
dei crinoidi (Traumatocrinus caudex) del Mesozoico (circa 220 milioni d’anni fa) <strong>della</strong><br />
Cina meridionale. Una grande lastra ne mostra due completi con le parti superiori<br />
perfettamente conservate (immagine in basso a sinistra). La parte con i peduncoli (mostrata<br />
a fianco), è la prova del modo di vivere perforando il legno. Oggi non ci sono<br />
più crinoidi che adottano questo sistema di vita, probabilmente perché i legno così<br />
perforato, dopo un tempo relativamente corto, affonderebbe sul fondo, dove i grandi<br />
“gigli di mare” non potrebbero vivere.<br />
11
Contattare: Dr. Hans Hess, libero collaboratore del Museo<br />
12
Da: http://www.nmb.bs.ch/NaturmuseumBasel/Frameset1/Frameset.html<br />
E-mail: hans.hess@bluewin.ch<br />
Bibliografia telematica dei vari capitoli<br />
http://www.ucmp.berkeley.edu/echinodermata/feed_arms.html<br />
http://www.ucmp.berkeley.edu/echinodermata/calyx.html<br />
http://www.ucmp.berkeley.edu/echinodermata/stem.html<br />
http://www.ucmp.berkeley.edu/echinodermata/tegmen.html<br />
http://www.ucmp.berkeley.edu/echinodermata/tegmen.html<br />
http://www.ucmp.berkeley.edu/echinodermata/cup.html<br />
http://www.ucmp.berkeley.edu/echinodermata/gut.html<br />
http://www.ucmp.berkeley.edu/echinodermata/stem_lumen.html<br />
http://www.ucmp.berkeley.edu/echinodermata/tegmen.html<br />
http://www.ucmp.berkeley.edu/echinodermata/feed_arms.html<br />
http://www.ucmp.berkeley.edu/echinodermata/circum_ring.html<br />
Tutti sono inclusi nel sito:<br />
http://www.ucmp.berkeley.edu/echinodermata/<br />
13
1) Quando appaiono i primi echinodermi?<br />
(una sola risposta)<br />
ڤ<br />
Permiano inferiore<br />
ڤ<br />
Cambriano inferiore<br />
ڤ<br />
Carbonifero superiore<br />
ڤ<br />
Giurassico<br />
<strong>Echinodermi</strong> Quiz<br />
2) A quale gruppo principale appartengono i crinoidi?<br />
(una sola risposta)<br />
ڤ<br />
<strong>Echinodermi</strong><br />
ڤ<br />
Molluschi<br />
ڤ<br />
Cnidari<br />
ڤ<br />
Artropodi<br />
3) Quale è una caratteristica comune a tutti i gruppi degli echinodermi?<br />
(una sola risposta)<br />
ڤ<br />
Presenza di una teca<br />
ڤ<br />
Il sistema vascolare acquifero<br />
ڤ<br />
La presenza dei radioli<br />
ڤ<br />
Avere le braccia<br />
4) Quale di queste non costituisce una forma d’echinide?<br />
(una sola risposta)<br />
ڤ<br />
Emisferica<br />
ڤ<br />
Triangolare<br />
ڤ<br />
Discoidale<br />
ڤ<br />
Sferoidale<br />
5) Qual è la ragione principale dei pedicelli ambulacrali<br />
(una sola risposta)<br />
ڤ<br />
Per protezione<br />
ڤ<br />
Per dormire, riposare<br />
ڤ<br />
Per il movimento<br />
ڤ<br />
Per mangiare<br />
6) Quale potrebbe essere il modo di vita più confacente ad un “dollaro delle sabbie”?<br />
(una sola risposta)<br />
ڤ<br />
Infaunale superficiale<br />
ڤ<br />
Libero natante<br />
ڤ<br />
Epifaunale<br />
ڤ<br />
Planctonico<br />
7) Quale caratteristica morfologica è più vantaggiosa ad un crinoide?<br />
(una sola risposta)<br />
ڤ<br />
Un corpo corto<br />
ڤ<br />
Una glabella larga<br />
ڤ<br />
Possedere braccia flessibili<br />
ڤ<br />
Lunghe spine
8) Quale può essere il motivo principale di non poter vivere in acqua salmastra?<br />
(una sola risposta)<br />
ڤ<br />
Possedere le parti scheletriche secrete dal mesoderma<br />
ڤ<br />
Il sistema acquifero regolato da pressione osmotica<br />
ڤ<br />
Possedere peristoma e periprocto molto distanti fra loro<br />
ڤ<br />
Presenza <strong>della</strong> lanterna di Aristotele<br />
9) Quando ebbero il loro massimo periodo di sviluppo i crinoidi?<br />
(una sola risposta)<br />
ڤ<br />
tardo Permiano<br />
ڤ<br />
primo Permiano<br />
ڤ<br />
primo Carbonifero<br />
ڤ<br />
tardo Cretacico<br />
10) Quale è stata la durata temporale degli echinidi irregolari in tempo assoluto?<br />
(una sola risposta)<br />
ڤ<br />
500 milioni d’anni<br />
ڤ<br />
350 milioni d’anni<br />
ڤ<br />
100 milioni d’anni<br />
ڤ<br />
200 milioni d’anni<br />
Ogni risposta esatta = 1 punto, max = 10 punti<br />
Pere vedere le risposte corrette clicca su <strong>Echinodermi</strong> Quiz soluzioni i
<strong>Echinodermi</strong> Quiz Soluzioni<br />
1) Quando appaiono i primi echinodermi?<br />
(una sola risposta)<br />
ڤ Permiano inferiore<br />
ڤ Cambriano inferiore……………………………………………………………………◄<br />
ڤ Carbonifero superiore<br />
ڤ Giurassico<br />
2) A quale gruppo principale appartengono i crinoidi?<br />
(una sola risposta)<br />
ڤ <strong>Echinodermi</strong>……………………………………………………………………………◄<br />
ڤ Molluschi<br />
ڤ Cnidari<br />
ڤ Artropodi<br />
3) Quale è una caratteristica comune a tutti i gruppi degli echinodermi?<br />
(una sola risposta)<br />
ڤ Presenza di una teca<br />
ڤ Il sistema vascolare acquifero…………………………………………………………◄<br />
ڤ La presenza dei radioli<br />
ڤ Avere le braccia<br />
4) Quale di queste non costituisce una forma d’echinide?<br />
(una sola risposta)<br />
ڤ Emisferica<br />
ڤ Triangolare……………………………………………………………………………◄<br />
ڤ Discoidale<br />
ڤ Sferoidale<br />
5) Qual è la ragione principale dei pedicelli ambulacrali<br />
(una sola risposta)<br />
ڤ Per protezione<br />
ڤ Per dormire, riposare<br />
ڤ Per il movimento……………………………………………………………………◄<br />
ڤ Discoidale<br />
ڤ Per mangiare<br />
6) Quale potrebbe essere il modo di vita più confacente ad un “dollaro delle sabbie”?<br />
(una sola risposta)<br />
ڤ Infaunale superficiale………………………………………………………………◄<br />
ڤ Libero natante<br />
ڤ Epifaunale<br />
ڤ Planctonico<br />
7) Quale caratteristica morfologica è più vantaggiosa ad un crinoide?<br />
(una sola risposta)<br />
ڤ Un corpo corto<br />
ڤ Una glabella larga<br />
ڤ Possedere braccia flessibili…………………………………………………………◄<br />
ڤ Lunghe spine
8) Quale può essere stato il motivo principale di non poter vivere in acqua salmastra?<br />
(una sola risposta)<br />
ڤ Possedere le parti scheletriche secrete dal mesoderma<br />
ڤ Il sistema acquifero regolato da pressione osmotica………………………………◄<br />
ڤ Possedere peristoma e periprocto molto distanti fra loro<br />
ڤ Presenza <strong>della</strong> lanterna di Aristotele<br />
9) Quando ebbero il loro massimo periodo di sviluppo i crinoidi?<br />
(una sola risposta)<br />
ڤ tardo Permiano<br />
ڤ primo Permiano<br />
ڤ primo Carbonifero…………………………………………………………………◄<br />
ڤ tardo Cretacico<br />
10) Quale è stata la durata temporale degli echinidi irregolari in tempo assoluto?<br />
(una sola risposta)<br />
ڤ 500 milioni d’anni<br />
ڤ 350 milioni d’anni<br />
ڤ 100 milioni d’anni<br />
ڤ 200 milioni d’anni……………………………………………………………………◄<br />
Ogni risposta esatta = 1 punto, max = 10 punti