8. predavanje
8. predavanje
8. predavanje

Create successful ePaper yourself
Turn your PDF publications into a flip-book with our unique Google optimized e-Paper software.
EFEKTORNE FUNKCIJE<br />
HUMORALNE IMUNOSTI<br />
ZAPOČINJANJE ZAPO INJANJE HUMORALNOG IMUNOG<br />
ODGOVORA<br />
Antigeni mikroorganizama koji prodiru u tkiva ili se nađu u krvi<br />
transportuju se i sakupljaju u perifernim limfoidnim organima.<br />
Humoralni imunski odgovor započinje kada antigen-specifični B- limfociti<br />
u folikulima slezine, limfnih čvorova i mukoznog limfoidnog tkiva<br />
prepoznaju antigene.<br />
U folikulima B limfociti vezuju antigene svojim receptorima,<br />
membranskim Ig (IgM, IgD).<br />
Antigeni se prepoznaju u svojoj nativnoj konformaciji i za to nije<br />
potrebna njihova prerada.<br />
Prepoznavanjem antigena dolazi do generisanja signalnih puteva koji<br />
dovode do aktivacije B limfocita.<br />
TIPOVI HUMORALNOG IMUNOG<br />
ODGOVORA U ZAVISNOSTI OD VRSTE<br />
ANTIGENA<br />
T-zavisni imuni odgovor: odgovor na proteinske<br />
antigene.<br />
Zavisan od pomoći CD4 + T limfocita.<br />
T-nezavisni imuni odgovor: odgovor na<br />
neproteinske antigene.<br />
Antigenom indukovan prenos signala<br />
u B-limfocitima<br />
BB-limfocitima<br />
limfocitima<br />
• da bi proces aktivacije započeo, moraju se približiti dva ili više<br />
receptorskih molekula i unakrsno povezati što se postiže<br />
agregacijom dva ili više antigenskih molekula.<br />
• signali sa receptora se prenose preko proteina koji su udruženi sa<br />
receptorima.<br />
• ovi proteini su nekovalntno vezani sa receptornim molekulima<br />
(obično IgM ili IgD) i nazivaju se Igα i Igβ formirajući B receptorski<br />
kompleks (BCR)<br />
• intracitoplazmatski region Igα i Igβ je znatno duži nego<br />
intracitoplazmatski region Ig receptora i sadrži tirozinske aktivacione<br />
motive (ITAM)<br />
• Pored Ig receptornih molekula Igα i Igβ proteina u okviru B ćelijskog<br />
receptorskog kompleksa nalaze se i enzimi kinaze koje vrše<br />
fosforilaciju<br />
Antigenom indukovan prenos signala<br />
u B-limfocitima<br />
BB-limfocitima<br />
limfocitima<br />
• nakon grupisanja dva ili više antigenskih receptora vezivanjem za<br />
odgovarajuće antigene dolazi do aktivacije kinaza, koje se nalaze u<br />
B-ćelijskom receptoru, koje vrše fosforilaciju tirozina u ITAM motivu<br />
čime se stvaraju fosfotirozini. (prvi prvi signal) signal<br />
• za fosfotirozine se zatim vezuju tzv. adapterski proteini koji se i sami<br />
fosforilišu i angažuju brojene signalne molekule.<br />
• signalni molekuli aktiviraju transkripcione faktore koji opet aktiviraju<br />
gene koji su zaduženi za procese proliferacije i diferencijacije Blmfocita<br />
Uloga komplementa<br />
Pored imunoglobulinski receptora, B-limfociti na svojoj površini<br />
poseduju i tzv. CD2 receptore za koje se vezuje C3d komponenta<br />
komplementa<br />
Vezivanje C3d komponente za CD2 rceptor pojačava poja ava aktivaciju B-<br />
limfocita – ovo je tzv drugi signal koji zajedno sa prvima pokreće pokre e<br />
proliferaciju i diferencijaciju B-limfocita<br />
B limfocita<br />
1
Antigenom indukovan prenos signala<br />
u B-limfocitima<br />
BB-limfocitima<br />
limfocitima<br />
Antigenom indukovan prenos signala<br />
u B-limfocitima<br />
BB-limfocitima<br />
limfocitima<br />
PLC – fosfolipaza C<br />
NFAT – nukleusni faktor<br />
aktivisanih T-limfocita<br />
NF- κB – nukleusni<br />
faktor kapa B<br />
AP-1 – aktivacioni<br />
protein 1 koji de sastoji od<br />
fosforilisanog c-Jun i c-<br />
Fos<br />
ERK – ekstracelularnim<br />
signalom aktivisana<br />
kinaza<br />
JNK – c-jun<br />
aminoterminalna kinaza<br />
Pored diferencijacije i proliferacije koja nastaje antigenskom<br />
stimulacijom B- BB-<br />
limfocita, dolazi do promena na njihovoj površini povr površini ini koje<br />
ih pripremaju za dalju saradnju sa T- TT-<br />
limfocitima:<br />
• ekspresija B7 kostimulatora koji obezbeđuje<br />
signal za aktivaciju T-limfocita<br />
• ekspresija receptora za citokine koji su<br />
sekretovani od strane pomoćničkih T limfocita<br />
• smanjenje ekspresije receptora za hemokine<br />
čija je funkcija zadržavanje B limfocita u<br />
folikulima<br />
Antigenom indukovan prenos signala<br />
u B-limfocitima<br />
BB-limfocitima<br />
limfocitima<br />
Posledice aktivacije B-limfocita<br />
antigenom i komponentama<br />
komplementa su:<br />
• započinjanje proliferacije B-limfocita i njihove<br />
diferencijacije u efektorske ćelije<br />
• priprema B-limfocita za interakciju sa Tlimfocitima<br />
pomoćnicima.<br />
Aktivisani naivni B-limfociti ulaze u<br />
ćelijski ciklus (proliferacija i formiranje<br />
klonova limfocita).<br />
Takođe, neki B-limfociti se diferntuju u<br />
plazmocite koji sekretuju IgM antitela.<br />
Ovaj proces je izraženiji kada su u pitanju<br />
multivalentni neproteinski antigeni, koji su<br />
takođe potentni aktivatori komplementa.<br />
Pošto su proteinski antigeni slabi<br />
aktivatori B-limfocita imunski odgovor na<br />
njih je slab. Međutim, oni indukuju<br />
promene u B-limfocitima koje im<br />
omogućuju interakciju sa T-limfocitima.<br />
Te promene su indukcija ekspresije B7<br />
molekula i indukcija receptora za citokine,<br />
čime se stiče sposobnost odgovora na<br />
citokine produkovane od strane T-limfocita.<br />
S druge strane, aktivisani B-limfociti<br />
smanjuju ekspresiju receptora za hemokine<br />
što im onemogućava dalje zadržavanje u<br />
folikulima i obezbeđuje migraciju izvan<br />
folikula<br />
HUMORALNI IMUNI ODGOVOR NA PROTEINSKE ANTIGENE<br />
Humoralni imuni odgovor na proteinske antigene je zavisan od Tlimfocita<br />
pomoćnika (CD4 + T-limfocita).<br />
U ovim procesima neophodan je kontakt između B limfocita i CD4 + Tlimfocita.<br />
Na taj način se obezbeđuju preduslovi za T-B kooperaciju.<br />
2
HUMORALNI IMUNSKI ODGOVOR NA PROTEINSKE<br />
ANTIGENE<br />
Ovaj tip imunskog odgovora se odvija u više faza:<br />
a) aktivacija T-limfocita proteinskim antigenom i diferencijacija T-limfocita<br />
u efektorske ćelije<br />
b) aktivacija B-limfocita antigenom i prezentacija antignskih peptida<br />
efektorskim T-lifmocitima<br />
c) aktivacija B-limfocita od strane efektorskih CD4 + T-limfocita<br />
d) diferencijacija aktivisanih B-limfocita u efektorske ćelije<br />
f) indukcija procesa izotipskog prekopčavanja<br />
g) indukcija procesa afinitetnog sazrevanja<br />
B-limfociti prepoznaju nativni antigen.<br />
Nakon vezivanja antigena dolazi do<br />
receptorom posredovane internalizacije<br />
antigena u citoplazmu B-limfocita, a zatim do<br />
njegove proteolize po tipu prerade egzogenih<br />
antigena i ispoljavanje peptidnih fragmenata<br />
u sklopu MHC molekula II klase.<br />
Takođe, aktivacija antigenom dovodi do<br />
ekspresije B7 molekula na membrani Blimfocita.<br />
CD4 + T-limfociti prepoznaju antigenske<br />
peptide u sklopu MHC molekula II klase na Blimfocitima.<br />
Angažovanjem kostimulatora (B7-CD28)<br />
dolazi do aktivacije CD4 + T-limfocita.<br />
Ovim procesima omogućava se interakcija<br />
limfocita koji su iste specifičnosti za antigen.<br />
B-limfocit prepoznaje nativne<br />
(konformacione) epitope na protienskom<br />
antigenu, dok CD4 + T-limfocit prepoznaje<br />
peptidne fragmente tog istog proteina.<br />
CD4 + T-limfociti prepoznaju antigenske peptide prezentovane od strane B limfocita.<br />
Obezbeđivanjem drugog signala aktivacije preko kostimulatora (B7-CD28) dolazi do<br />
aktivacije T-limfocita koji eksprimira CD40 ligand (CD40L) i sekretuje citokine.<br />
Vezivanjem CD40L od strane CD40 molekula na B-limfocitu kao i vezivanjem citokina<br />
preko receptora za citokine na B-limfocitu omogućena je proliferacija (klonska ekspanzija)<br />
B-limfocita i njihova diferencijacija u efektorske ćelije koje sekretuju antitela.<br />
CD4 + T-limfociti prepoznaju u regionu parakorteksa limfnog čvora prerađene antigenske<br />
peptide ispoljene na profesionalnim APC, nakon čega nastaje proliferacija i diferencijacija<br />
CD4 + T limfocita u efektorske CD4 + T-limfocite.<br />
Ovi efektorski T-limfociti migriraju iz parakorteksa ka limfnom folikulu.<br />
Naivni B-limfociti, koji su u folikulu aktivisani antigenom migriraju iz folikula.<br />
Na periferiji folikula ćelije dolaze u međusobni kontakt, nakog čega započinje T-B<br />
interakcija (kooperacija).<br />
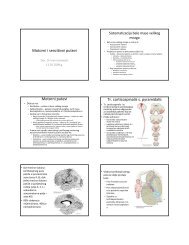
ANTITELA<br />
Mesto za vezivanje antigena -PARATOP<br />
IMUNOLOŠKA IMUNOLO KA SINAPSA<br />
3
IZOTIPSKO PREKOPČAVANJE (“SWITCH”)<br />
Produkcija antitela različitih klasa (izotipova)<br />
U toku procesa<br />
prekopčavanja (na nivou<br />
primarnog transkripta) Sµ se<br />
povezuje sa odgovarajućim S<br />
sekvencama drugih C gena.<br />
Svi ostali nukleotidi između S<br />
sekvenci se uklanjaju procesom<br />
delecije<br />
Prikazan je primer izotipskog<br />
prekopčavanja za sintezu<br />
teškog lanca IgG i teškog lanca<br />
IgE<br />
Stimulacija produkcije IgG (podklase IgG 1 i IgG3) je pod uticajem IFN-gama.<br />
Ove klase antitela učestvuju u procesima opsonizacije i fagocitoze (posredovani Fc<br />
receptorom), u aktivaciji komplementa i neonatalnom imunitetu.<br />
IL-4 utiče na produkciju IgE koji je bitan za imunitet prema helmintima i reakciji ranog tipa<br />
preosetljivosti.<br />
Citokini (kao što je TGF beta) utiču na sekreciju IgA u mukoznim tkivima koji su važni za<br />
lokalni imunitet sluznica.<br />
U B-limfocitu koji sekretuje<br />
antitela IgM klase došlo je do<br />
preuređenja genskih<br />
segmenata (VDJ povezan sa<br />
najbližim<br />
segmentom<br />
Cµ genskim<br />
Preuređnje VDJ se dešava na<br />
nivou DNK a povezivanje VDJ<br />
sa Cµ na nivou primarnog<br />
transkripta<br />
Signali od strane T-limfocita<br />
(ligacija CD40, i citokini)<br />
indukuju rekombinaciju<br />
prekopčavajućih<br />
regiona<br />
(“switch”)<br />
Switch (S) regioni(Sµ, Sγ, Sε,<br />
Sα) su konzervisane sekvence<br />
nukleotida smeštene ispred<br />
odgovarajućih C gena. Jedino<br />
Cδ nema S sekvence<br />
Pomoćnički T-limfociti stimulišu B-limfocite da produkuju antitela<br />
različitih klasa (izotipova).<br />
Za sekreciju IgM, čija je osnovna efektorska funkcija aktivacija<br />
komplementa, nije neophodna pomoć od strane T-limfocita.<br />
Pod uticajem CD40L ispoljenog na pomoćničkim T-limfocitima i<br />
citokina koje ove ćelije produkuju dolazi do procesa izotipskog<br />
prekopčavanja u B-limfocitima i sekrecije drugih klasa antitela.<br />
EFEKTORNE FUNKCIJE HUMORALNE IMUNOSTI<br />
Imunost posredovana antitelima se odlikuje brojnim efektornim<br />
mehanizmima koje organizam koristi pre svega u cilju zaštite od<br />
ekstracelularnih mikroorganizama i njihovih toksina ali je njegova<br />
uloga takođe jako bitna i u odbrani od onih mikroorganizama kao što<br />
su virusi koji žive i razmnožavaju se u citoplazmi ćelija domaćina.<br />
Antitela sprečavaju nastanak infekcije<br />
na jedan od sledećih načina<br />
• blokiraju sposobnost mikroorganizma da se vežu za ćelije<br />
domaćina<br />
• vezuju se za toksine mikroorganizama<br />
• olakšavaju proces fagocitoze procesom opsonizacije<br />
• zajedno sa T limfocitima učestvuju u uništavanju<br />
mikroorganizama<br />
• aktiviraju sistem komplementa<br />
4
Neutralizacija – Blokiranje biološke<br />
aktivnost ciljnih molekula (toksin se<br />
veže za svoj receptor)<br />
Opsonizacija – Interakcija sa specifičnim<br />
receptorimanarazličitim ćelijama,<br />
uključujući makrofage, neutrofile,<br />
bazofile i mast ćelije omogućavajući im da<br />
“prepoznaju" i odgovore na antigen<br />
Aktivacija komplementa – Dovodi do direktne<br />
lize i poboljšava fagocitozu<br />
KAKVA JE FUNKCIJA POJEDINIH<br />
DELOVA MOLEKULA ANTITELA?<br />
Preko hipervarijabilnog regiona Fab fragmenta svako antitelo<br />
se vezuje za odgovarajući odgovaraju i antigen koji je smešten sme ten na površini povr ini<br />
nekog mikroorganizma ili na molekulu nekog toksina. Na taj<br />
način na in se blokiraju štetni tetni efekti bakterije ili bakterijskog tkosina.<br />
Fc fragment antitela se koristi za konačnu elminaciju<br />
mikroorganizma ili toksina odnosno za aktivaciju tzv.<br />
efektorskih mehanizama. Fc regionom različitih<br />
imunoglobulinskih izotipova, antitela se vezuju ili za tzv, Fc<br />
receptore na fagocitnim ćelijama ili se komponente<br />
komplementa vezuju za Fc fragment čime počinje aktivacija<br />
čitavog sistema komplementa. Funkcija Fc fragmenta je<br />
moguća jedino pod uslovom da je antitelo vezano za antigen<br />
preko Fab fragamenta<br />
Fc RECEPTORI<br />
Svaki Fc receptor ima svoj:<br />
• ekstracelularni deo, (kojim se vezuje za Fc fragment<br />
imunoglobulina)<br />
• Intramembranski<br />
• intracitoplazmatski<br />
Intracitoplazmatski deo Fc receptora je predstavljem tzv.<br />
tirozinkinaznim imunoreceptorskim motivom koji može biti<br />
aktivacioni (ITAM) i inhibitorni (ITIM).<br />
Fc RECEPTORI<br />
Antitela često esto ostvaruju svoj zaštitni za titni efekat vezujući vezuju i se Fab fragmentom<br />
za antigenske determinante mikroorganizama ili toksina čime ime sprečavaju spre avaju<br />
vezivanje mikroorganizama za ćelije elije domaćina. doma ina.<br />
Mnogo češće e biološka biolo ka i protektivna funkcija antitela se ostvaruje<br />
vezivanjem antitela preko Fc fragmenta za receptor na efektornim<br />
ćelijama elijama ili komplement, pa se takvi receptori nazivaju Fc receptorima.<br />
receptorima<br />
Fc<br />
c receptori se mogu naći na naći i na različitim razli različitim itim ćelijama elijama kao kao što to su: su su:<br />
• Neutrofili<br />
• Makrofagi<br />
• B limfociti<br />
• Mastociti<br />
• Bazofilni leukociti<br />
• Trombociti i dr.<br />
Fc RECEPTORI<br />
Fosforilacija ITAM motiva uslovljava:<br />
• Pojačanu fagocitozu<br />
• Antitelno zavisnu ćelijsku<br />
citotoksičnost<br />
• Oslobađanje medijatora<br />
• Pojačanu antigensku prezentaciju<br />
Fosforilacija ITIM motiva uslovljava:<br />
• Smanjenje svih gore pomenutih<br />
ćelijskih aktivnosti<br />
5
Fc RECEPTORI Fc RECEPTORI<br />
Fc RECEPTORI<br />
NEUTRALIZACIJA<br />
Neutralizacija je proces u kome se antitela vezuju za<br />
antigenske determinante mikroorganizama ili njihovih<br />
toksina čime ime neutrališu neutrali u njihovu infektivnost odnosno<br />
toksične toksi ne efekte.<br />
Većina Ve ina mikroorganizama se za ćelije elije domaćina doma ina vezuju<br />
preko molekula koji su sastavni deo njihovog omotača. omota a.<br />
Antitela imaju sposobnost da se vežu ve u za ove molekule i<br />
na taj način na in spreče spre e njihovo vezivanje za ćelije elije domaćina doma ina<br />
Većina Ve ina egzotoksina bakterija takođe imaju sposobnost da<br />
se vezuje za ćelije elije domaćina doma ina i izazovu štetne tne efekte. U<br />
procesu neutralizacije antitela se takođe vezuju za<br />
molekule toksina onemugućavaju<br />
onemugu avajući i im vezivanje za ćelije. elije.<br />
Promena klase teških lanaca dovodi do produkcije antitela koja su u<br />
zavisnosti od prirode mikroorganizma optimalna za eliminaciju tog<br />
mikroorganizma,<br />
Procersom sazrevanja afiniteta koji se postiže posti e prolongiranom i ponavljanom<br />
stimulacijom stvara se sve veći ve i afinitet vezivanja antitela za odgovarajući<br />
odgovaraju i<br />
antigen odnosno povećava pove ava sposobnost vezivanja antitela.<br />
6
OPSONIZACIJA<br />
Predstavlja proces u kome antitela oblažu obla oblažu u površinu povr površinu inu<br />
mikroorganizama vezujući vezuju vezujući i se svojim Fab fragmentima za antigenske<br />
determinante na površini povr površini ini bakterija. bakterija. Takva Takva antitela antitela se nazivaju<br />
OPSONINI.<br />
Kada se nekoliko molekula antitela, opsonina, veže svojim Fab<br />
fragmentima za površinu bakterije, njihovi Fc regioni formiraju niz<br />
koji prominira sa površine i ako su to antitela npr. IgG1 ili IgG3 oni<br />
će se vezati za Fc receptore na neutrofilima ili makrofagima (FcγRI).<br />
Vezivanje opsonizovanih bakterija za Fc receptore na fagocitnim<br />
ćelijama olakšava proces ingestije ali isto tako izaziva i povećanu<br />
sekreciju lizozomalnih enzima, reaktivnih kiseoničkih metabolita,<br />
azot monoksida i proteolitičkih enzima što povećava sposobnost<br />
ubijanja fagocitovanih bakterija i njihovu razgradnju<br />
Opsonizacija ima najveću ulogu u fagocitozi tzv, inkapsuliranih<br />
bakterija kao što je pneumokok.<br />
OPSONIZACIJA<br />
OPSONIZACIJA ĆELIJSKA ELIJSKA CITOTOKSIČNOS<br />
CITOTOKSI<br />
CITOTOKSIČNOST NOST T ZAVISNA<br />
OD ANTITELA - ADCC<br />
ĆELIJSKA ELIJSKA CITOTOKSIČNOS<br />
CITOTOKSI<br />
CITOTOKSIČNOST NOST ZAVISNA<br />
OD ANTITELA - ADCC<br />
Proces ubijanja mikroorganizama koji su obloženi oblo eni antitelima a od<br />
strane NK ćelija elija ili eozinofila.<br />
NK ćelije elije na svojoj površini povr ini eksprimiraju Fc receptore za koje se<br />
vezuju Fc fragmenti opsonizujućih opsonizuju ih antitela. Ovo predstavlja signal za<br />
NK ćeliju eliju da iz svojih granula počne po ne da oslobađa sadržaj sadr aj koji je<br />
citotoksičan citotoksi an za opsonizovane ćelije elije<br />
Na sličan sli an način na in eozinofilni leukociti, koji imaju na svojoj površini povr ini Fc<br />
receptore, se vezuju za opsonizovane helminte. U ovom slučaju slu aju<br />
opsonizujuća opsonizuju a antitela su iz klase IgE. Nakon vezivanja za helminte iz<br />
granula eozinofilnih leukocita se oslobađa sadržaj sadr aj koji može mo e da prodre<br />
kroz debeli omotač omota (tegument) i prodre u unutrašnjost unutra njost parazita i<br />
izazove njegovu smrt.<br />
Sistem komplement<br />
komplementa<br />
a (C)<br />
o Sistem proteina plazme koji mogu biti direktno<br />
aktivirani patogenima ili indirektno patogenima<br />
obloženim oblo enim antitelima.<br />
o Proizvode kaskadu reakcija na površini povr ini patogena.<br />
o Stvara aktiv tivne ne komponente sa različitim razli itim<br />
efektornim funkcijama (inflama inflamacija ja, , pojačava poja ava<br />
fagocitozu, stvara kompleks koji ošte o tećuje uje<br />
membranu).<br />
membranu<br />
o Svežem Sve em serumu daje liti itičku ku aktiv tivnost, nost, koja se<br />
razara grejanjem na 56 oC C tokom 30 min.<br />
min<br />
7
Sistem komplement<br />
komplementa<br />
a (C) Funkcije komplementa<br />
Sistem komplementa je glavni efektorni mehanizam humoralnog imuniteta imuniteta<br />
i<br />
takođe važan va an efektorni mehanizam prirodnog imuniteta.<br />
Sastoji se od serumskih proteina i proteina u ćelijskoj elijskoj membrani koji reaguju<br />
međusobno kao i sa drugim molekulima imunog sistema na način na in koji je visoko<br />
kontrolisan.<br />
Aktivacija komplementa obuhvata sekvencijalnu proteolizu proteina proteina<br />
komplementa pri čemu emu se stvaraju proteolitički proteoliti ki enzimi.<br />
Proteini koji čine ine sistem komplementa su proteini plazme koji su normalno<br />
inaktivni; aktiviraju aktiviraju<br />
se u određenim uslovima da bi stvorili produkte koji<br />
ostvaruju brojne efektorne funkcije komplementa.<br />
Produkti aktivacije komplementa se kovalentno vezuju za površinu povr inu<br />
bakterija ili za antitela vezana za bakterije ili druge antigene. antigene<br />
Aktivaciju komplementa inhibiraju regulatorni proteini koji su prisutni prisutni<br />
u<br />
ćelijskoj elijskoj membrani domaćina doma ina a nema ih na membrani bakterija.<br />
Proteini sistema komplementa<br />
C1(qrs), C2, C3, C4, C5, C6, C7, C8, C9<br />
faktor fa tori B, D, H, H I, properdin (P)<br />
mannose binding lectin (MBL),<br />
C1 inhibitor (C1-INH, (C1 INH, serpin), serpin),<br />
C4-vezuju C4 vezujući i protein protein<br />
(C4-BP), (C4 BP), faktor koji ubrzava raspadanje (DAF - decay<br />
accelerating factor ),<br />
C1 receptor (CR1), protein-S protein S (vitrone ( vitronektin) tin)<br />
- Izotipovi antitela (IgM (IgM > IgG3 gG3 > IgG1>> IgG2 IgG2) gG2) ) kada se vežu ve vežu u za antigen<br />
- Kompleks Ag-At Ag Ag-At At<br />
- Virusi<br />
- Subcelularne membrane<br />
- Nekrotične<br />
Nekroti Nekrotične ne ćelije<br />
elije<br />
- Agregirani Ig<br />
- β-amiloid amiloid (Alzheimer)<br />
(Alzheimer)<br />
- CRP<br />
LEKTINSKI PUT - aktivira se, u odsustvu antitela, antitela antitela, , na terminalnim reziduama re reziduama ziduama<br />
manoze proteina ili polisaharida na ćelijskoj elijskoj membrani membrani bakterija<br />
ALTERNATIVNI PUT – aktiviraju skoro sve strane supstance:<br />
- LPS<br />
- Endotoksin iz ćelijskog elijskog zida gram-negativnih gram gram-negativnih negativnih bakterija<br />
bakterija<br />
- Ćelijski elijski zidovi nekih vrsta kvasca<br />
- Proteini kobrinog otrova (Cobra venom faktor)<br />
Povoljni efekti delovanja komplementa:<br />
- Opsonizacija Opsonizacija<br />
i olakšavanje olak avanje fagocitoze<br />
- Hemotaksa i aktivacija fagocita<br />
- Liza bakteri ba terija a i inficiranih ćelija elija<br />
- Regula egulacija cija humoralnog imunog<br />
odgovora<br />
- Uklanjanje imunih kompleksa<br />
- Uklanjanje apoptotičnih<br />
apoptoti nih ćelija elija<br />
Nepovoljni efekti delovanja komplementa:<br />
- Inflamacija<br />
Inflamacija<br />
- Ana nafilaksa filaksa<br />
Putevi utevi aktivacije komplement<br />
komplementa<br />
KLASIČNI PUT LEKTINSKI PUT<br />
Zavisan od antitela<br />
AKTIVACIJA C3 I GENERISANJE<br />
C5 KONVERTAZE<br />
AKTIVACIJA C5<br />
LITIČKI PUT<br />
Nezavisni od antitela<br />
ALTERNATIVNI PUT<br />
Putevi utevi aktivacije komplement<br />
komplementa<br />
C1 protein komplementa postaje aktivan,<br />
vezuje i razlaže druga dva proteina<br />
komplementa: C4 i C2<br />
KLASIČNI KLASI KLASIČNI NI PUT - aktiviraju:<br />
Nastaje kompleks C4bC2b koji se<br />
kovalentno veže za ćelijsku površinu<br />
patogena taj kompleks se naziva:<br />
C3-konvertaza!<br />
C3-konvertaza razlaže C3 komponentu<br />
komplementa na dva fragmenta : C3a i<br />
C3b a nastala C3b komponenta se<br />
takođe veže za površinu patogena<br />
Neki od C3b molekula se veže za<br />
prisutni kompleks C4b2b i formira<br />
kompleks C4b2b3b koji se naziva<br />
C5-konvertaza<br />
8
C1 protein komplementa postaje aktivan,<br />
vezuje i razlaže druga dva proteina<br />
komplementa: C4 i C2<br />
Nastaje kompleks C4bC2b koji se<br />
kovalentno veže za ćelijsku površinu<br />
patogena taj kompleks se naziva:<br />
C3-konvertaza!<br />
C3-konvertaza razlaže C3 komponentu<br />
komplementa na dva fragmenta : C3a i<br />
C3b a nastala C3b komponenta se<br />
takođe veže za površinu patogena<br />
Neki od C3b molekula se veže za<br />
prisutni kompleks C4b2b i formira<br />
kompleks C4b2b3b koji se naziva<br />
C5-konvertaza<br />
KASNA FAZA AKTIVACIJE KOMPLEMENTA<br />
REGULATORNI PROTEINI AKTIVACIJE KOMPLEMENTA<br />
DAF – faktor koji ubrzava raspad: sprečva vezivanje<br />
faktora B za C3b ili vezivanje C4b2b za C3b<br />
Faktor 1 plazme: proteolitičko razlaganje C3b<br />
MCP – membranski kofaktorski protein: kofaktor za<br />
proteolitičko razlaganje C3b u neaktivne fragmente i<br />
deluje zajedno sa Faktorom 1<br />
C1 INH – C1 inhibitor zaustavlja aktivaciju<br />
komplementa u najranijim fazama<br />
C3b<br />
C3c<br />
C4a<br />
C4b<br />
C5a<br />
Bb<br />
C5b-9<br />
Raspadni produkt hidrolize C3<br />
komponente nazvan C3b se taloži talo i<br />
na ćelijskoj elijskoj površini povr ini patogena.<br />
Formiraju se stabilne veze sa<br />
polisaharidima ćelijske elijske membrane.<br />
C3b se zbog regulatornih proteina<br />
ne vezuje za ćelijsku elijsku membranu<br />
normalnih ćelija elija domaćina. doma ina.<br />
Za vezanu komponentu C3b se<br />
dalje vezuje faktor B koga ubrzo<br />
razlaže jedna proteaza plazme na<br />
Bb i Ba komponentu. Nastaje<br />
kompleks: C3bBb koji se naziva<br />
C3 konvertaza alternativnog puta.<br />
Kada se za ovu konvertazu veže<br />
još jedan molekul C3b nastaje<br />
kompleks<br />
naziva:<br />
C3bBbC3b koji se<br />
C5 konvertaza alternativnog puta<br />
Struktura litičkog kompleksa<br />
FIZIOLOŠKE FIZIOLO KE AKTIVNOSTI POJEDINIH AKTIVACIONIH<br />
PRODUKATA SISTEMA KOMPLEMENTA<br />
C3a<br />
Anafilatoksinska aktivnost (degranulacija eozinofila, mastocita i bazofila)<br />
Agregacija trombocita<br />
Opsonizacija<br />
Inhibicija formiranja imunskih kompleksa<br />
Započinjanje procesa čišćenja od imunskih kompleksa<br />
Neutralizacija virusa<br />
Oslobađanje neutrofila iz kostne srži<br />
Neke anafilatoksinske aktivnosti (indukcija oslobađanja histamina iz mastocita i<br />
bazofila što dovodi do vazodilatacije i kontrakcije mišića)<br />
Opsonizacija (poboljšana fagocitoza)<br />
Anafilatoksinska aktivnost<br />
Hemotaksa neutrofila<br />
Imunska adherenca za endotel krvnih sudova preko makrofagnih receptora.<br />
Inhibicija migracija<br />
Indukcija širenja makrofaga i monocita<br />
Oštećenje ćelijskih membrana (liza ciljnih ćelija)<br />
Neutralizacija virusa<br />
9