

Tactile sensory coding in the glabrous skin of the human hand
Tactile sensory coding in the glabrous skin of the human hand
Tactile sensory coding in the glabrous skin of the human hand

Create successful ePaper yourself
Turn your PDF publications into a flip-book with our unique Google optimized e-Paper software.
TINS -January 1983<br />
<strong>the</strong>m exhibit a static discharge related to<br />
jo<strong>in</strong>t position. Movements also excite all<br />
<strong>the</strong> FA II units, and even a majority <strong>of</strong> <strong>the</strong><br />
FA I and SA I units (57% and 66%, respec-<br />
tively) give some response. It seems<br />
reasonable to conclude that <strong>the</strong> cutaneous<br />
mechanoreceptors, and particularly <strong>the</strong> SA<br />
11 and FA 11 units, may provide not only<br />
exteroceptive but also proprioceptive<br />
<strong>in</strong>formation. The fact that units with func-<br />
tional and structural properties similar to<br />
<strong>the</strong> FA I1 and SA II units <strong>of</strong> <strong>the</strong> sk<strong>in</strong> are<br />
widely distributed <strong>in</strong> deeper fibrous tissues,<br />
e.g. jo<strong>in</strong>t capsulae, <strong>in</strong>terosseous mem-<br />
branes and tendon sheets, fits with this<br />
view.<br />
The functional role <strong>of</strong> cutaneous<br />
mechanoreceptors <strong>in</strong> motor control, k<strong>in</strong>es-<br />
<strong>the</strong>sia and position sense is not at all clear.<br />
There is evidence that <strong>the</strong>y may contribute<br />
to position sense and k<strong>in</strong>aes<strong>the</strong>sia <strong>in</strong> <strong>the</strong><br />
absence <strong>of</strong> afferent <strong>in</strong>put from jo<strong>in</strong>t and<br />
muscle tendon receptors ~9. Moreover,<br />
<strong>in</strong>formation <strong>in</strong> afferents from <strong>the</strong> f<strong>in</strong>gers<br />
accounts for a general facilitatory effect on<br />
motor commands to <strong>the</strong> <strong>hand</strong> and f<strong>in</strong>ger<br />
muscles,S.,9. However, recent f<strong>in</strong>d<strong>in</strong>gs<br />
<strong>in</strong>dicate a more specific role <strong>of</strong> <strong>the</strong>se units<br />
<strong>in</strong> <strong>the</strong> motor control. When <strong>hand</strong>l<strong>in</strong>g<br />
objects <strong>in</strong> <strong>the</strong> precision grip between <strong>the</strong> tips<br />
<strong>of</strong> <strong>the</strong> f<strong>in</strong>gers and <strong>the</strong> thumb, <strong>the</strong> <strong>human</strong><br />
subject automatically balances <strong>the</strong> grip<br />
forces to prevent <strong>the</strong> object from slipp<strong>in</strong>g or<br />
crack<strong>in</strong>g. This very accurate force control<br />
requires <strong>in</strong>formation from tactile units<br />
about <strong>the</strong> frictional conditions between <strong>the</strong><br />
sk<strong>in</strong> and <strong>the</strong> object (G. Westl<strong>in</strong>g and R. S.<br />
Johansson, unpublished observations). Of<br />
<strong>in</strong>terest <strong>in</strong> this context is that <strong>the</strong> primate<br />
motor cortex, which is known to play a key<br />
role <strong>in</strong> <strong>the</strong> control <strong>of</strong> f<strong>in</strong>e f<strong>in</strong>ger move-<br />
ments, receives detailed <strong>in</strong>formation over<br />
very rapid pathways from <strong>the</strong><br />
mechanoreceptive units <strong>of</strong> <strong>the</strong> <strong>glabrous</strong><br />
sk<strong>in</strong> ~7"<br />
Correlation between neuronal and<br />
psychophysical events<br />
The ultimate aim <strong>of</strong> analyses <strong>of</strong> correla-<br />
tions between activity <strong>in</strong> afferent units, on<br />
<strong>the</strong> one <strong>hand</strong>, and psychophysical<br />
phenomena, on <strong>the</strong> o<strong>the</strong>r, is to bridge <strong>the</strong><br />
gap between biophysical events <strong>in</strong> <strong>the</strong><br />
nervous system and mental events <strong>in</strong> <strong>the</strong><br />
m<strong>in</strong>d. Thus, we want to ga<strong>in</strong> <strong>in</strong>sight <strong>in</strong> <strong>the</strong><br />
basic rules that <strong>the</strong> bra<strong>in</strong> is bound to follow<br />
when its purpose is to produce a sensation<br />
from <strong>the</strong> afferent message <strong>in</strong> <strong>the</strong> peripheral<br />
nerves.<br />
Detection<br />
The concept <strong>of</strong> a <strong>sensory</strong> threshold has<br />
been much discussed and it has been'<br />
debated whe<strong>the</strong>r central or peripheral<br />
Psychophysical thresholds<br />
E 100 100 -<br />
-d<br />
-5<br />
t-<br />
O<br />
50 50-<br />
o o<br />
Neuronal thresholds<br />
• FAI+FAII [] SAI+SAII<br />
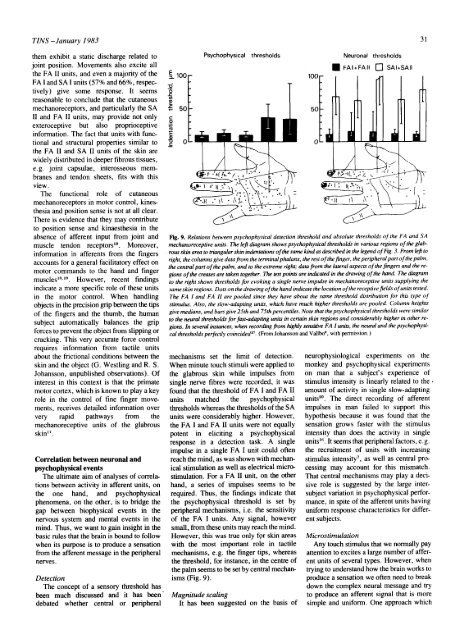
Fig. 9. Relations between psychophysical detection threshold and absolute thresholds <strong>of</strong> <strong>the</strong> FA and SA<br />
meehanoreceptive units. The left diagram shows psychophysical thresholds <strong>in</strong> various regions <strong>of</strong> <strong>the</strong> glab-<br />
rous sk<strong>in</strong> area to triangular sk<strong>in</strong> <strong>in</strong>dentations <strong>of</strong> <strong>the</strong> same k<strong>in</strong>d as described <strong>in</strong> <strong>the</strong> legend <strong>of</strong> Fig. 3. From left to<br />
right, <strong>the</strong> columns give data from <strong>the</strong> term<strong>in</strong>al phalanx, <strong>the</strong> rest <strong>of</strong> <strong>the</strong> f<strong>in</strong>ger, <strong>the</strong> peripheral part <strong>of</strong> <strong>the</strong> palm,<br />
<strong>the</strong> central part <strong>of</strong> <strong>the</strong> palm, and to <strong>the</strong> extreme right; data from <strong>the</strong> lateral aspects <strong>of</strong> <strong>the</strong> f<strong>in</strong>gers and <strong>the</strong> re-<br />
gions <strong>of</strong> <strong>the</strong> creases are taken toge<strong>the</strong>r. The test po<strong>in</strong>ts are <strong>in</strong>dicated <strong>in</strong> <strong>the</strong> draw<strong>in</strong>g <strong>of</strong> <strong>the</strong> <strong>hand</strong>. The diagram<br />
to <strong>the</strong> right shows thresholds for evok<strong>in</strong>g a s<strong>in</strong>gle nerve impulse <strong>in</strong> mechanoreceptive units supply<strong>in</strong>g <strong>the</strong><br />
same sk<strong>in</strong> regions. Dots on <strong>the</strong> draw<strong>in</strong>g <strong>of</strong> <strong>the</strong> <strong>hand</strong> <strong>in</strong>dicate <strong>the</strong> location <strong>of</strong> <strong>the</strong> receptive fields <strong>of</strong> units tested.<br />
The FA I and FA H are pooled s<strong>in</strong>ce <strong>the</strong>y have about <strong>the</strong> same threshold distribution for this type <strong>of</strong><br />
stimulus. Also, <strong>the</strong> slow-adapt<strong>in</strong>g units, which have much higher thresholds are pooled. Column heights<br />
give medians, and bars give 25th and 75th percentiles. Note that <strong>the</strong> psychophysical thresholds were similar<br />
to <strong>the</strong> neural thresholds for fast-adapt<strong>in</strong>g units <strong>in</strong> certa<strong>in</strong> sk<strong>in</strong> regions and considerably higher <strong>in</strong> o<strong>the</strong>r re-<br />
gions. In several <strong>in</strong>stances, when record<strong>in</strong>g from highly sensitive FA I units, <strong>the</strong> neural and <strong>the</strong> psychophysi-<br />
cal thresholds perfectly co<strong>in</strong>cided 27. (From Johansson and Vallbo 9, with permission. )<br />
mechanisms set <strong>the</strong> limit <strong>of</strong> detection.<br />
When m<strong>in</strong>ute touch stimuli were applied to<br />
<strong>the</strong> <strong>glabrous</strong> sk<strong>in</strong> while impulses from<br />
s<strong>in</strong>gle nerve fibres were recorded, it was<br />
found that <strong>the</strong> threshold <strong>of</strong> FA I and FA II<br />
units matched <strong>the</strong> psychophysical<br />
thresholds whereas <strong>the</strong> thresholds <strong>of</strong> <strong>the</strong> SA<br />
units were considerably higher. However,<br />
<strong>the</strong> FA I and FA II units were not equally<br />
potent <strong>in</strong> elicit<strong>in</strong>g a psychophysical<br />
response <strong>in</strong> a detection task. A s<strong>in</strong>gle<br />
impulse <strong>in</strong> a s<strong>in</strong>gle FA I unit could <strong>of</strong>ten<br />
reach <strong>the</strong> m<strong>in</strong>d, as was shown with mechan-<br />
ical stimulation as well as electrical micro-<br />
stimulation. For a FA U unit, on <strong>the</strong> o<strong>the</strong>r<br />
<strong>hand</strong>, a series <strong>of</strong> impulses seems to be<br />
required. Thus, <strong>the</strong> f<strong>in</strong>d<strong>in</strong>gs <strong>in</strong>dicate that<br />
<strong>the</strong> psychophysical threshold is set by<br />
peripheral mechanisms, i.e. <strong>the</strong> sensitivity<br />
<strong>of</strong> <strong>the</strong> FA I units. Any signal, however<br />
small, from <strong>the</strong>se units may reach <strong>the</strong> m<strong>in</strong>d.<br />
However, this was true only f9 r sk<strong>in</strong> areas<br />
with <strong>the</strong> most important role <strong>in</strong> tactile<br />
mechanisms, e.g. <strong>the</strong> f<strong>in</strong>ger tips, whereas<br />
<strong>the</strong> threshold, for <strong>in</strong>stance, <strong>in</strong> <strong>the</strong> centre <strong>of</strong><br />
<strong>the</strong> palm seems to be set by central mechan-<br />
isms (Fig. 9).<br />
Magnitude scal<strong>in</strong>g<br />
It has been suggested on <strong>the</strong> basis <strong>of</strong><br />
neurophysiological experiments on <strong>the</strong><br />
monkey and psychophysical experiments<br />
on man that a subject's experience <strong>of</strong><br />
stimulus <strong>in</strong>tensity is l<strong>in</strong>early related to <strong>the</strong>.<br />
amount <strong>of</strong> activity <strong>in</strong> s<strong>in</strong>gle slow-adapt<strong>in</strong>g<br />
units 2°. The direct record<strong>in</strong>g <strong>of</strong> afferent<br />
impulses <strong>in</strong> man failed to support this<br />
hypo<strong>the</strong>sis because it was found that <strong>the</strong><br />
sensation grows faster with <strong>the</strong> stimulus<br />
<strong>in</strong>tensity than does <strong>the</strong> activity <strong>in</strong> s<strong>in</strong>gle<br />
units xr. It seems that peripheral factors, e.g.<br />
<strong>the</strong> recruitment <strong>of</strong> units with <strong>in</strong>creas<strong>in</strong>g<br />
stimulus <strong>in</strong>tensity 7, as well as central pro-<br />
cess<strong>in</strong>g may account for this mismatch.<br />
That central mechanisms may play a deci-<br />
sive role is suggested by <strong>the</strong> large <strong>in</strong>ter-<br />
subject variation <strong>in</strong> psychophysical perfor-<br />
mance, <strong>in</strong> spite <strong>of</strong> <strong>the</strong> afferent units hav<strong>in</strong>g<br />
uniform response characteristics for differ-<br />
ent subjects.<br />
Microstimulation<br />
Any touch stimulus that we normally pay<br />
attention to excites a large number <strong>of</strong> affer-<br />
ent units <strong>of</strong> several types. However, when<br />
try<strong>in</strong>g to understand how <strong>the</strong> bra<strong>in</strong> works to<br />
produce a sensation we <strong>of</strong>ten need to break<br />
down <strong>the</strong> complex neural message and try<br />
to produce an afferent signal that is more<br />
simple and uniform. One approach which<br />
31






