Molecular phylogeny of Psilocybe cyanescens complex in Europe ...
Molecular phylogeny of Psilocybe cyanescens complex in Europe ...
Molecular phylogeny of Psilocybe cyanescens complex in Europe ...

You also want an ePaper? Increase the reach of your titles
YUMPU automatically turns print PDFs into web optimized ePapers that Google loves.
B. LSU D1 doma<strong>in</strong><br />
Maximum likelihood<br />
GTR model<br />
bootstraps<br />
ML/MP<br />
<strong>Psilocybe</strong> moravica (P06)<br />
100/- <strong>Psilocybe</strong> arcana (P05*)<br />
96/- P. moravica var. sternberkiana (P07)<br />
99/78<br />
<strong>Psilocybe</strong> bohemica (P14)<br />
<strong>Psilocybe</strong> serbica (P15)<br />
75<br />
<strong>Psilocybe</strong> <strong>cyanescens</strong> (P21*)<br />
Weraroa novae-zelandiae<br />
<strong>Psilocybe</strong> azurescens (P01*)<br />
<strong>Psilocybe</strong> <strong>cyanescens</strong><br />
80<br />
<strong>Psilocybe</strong> azurescens (P01*)<br />
<strong>Psilocybe</strong> subaerug<strong>in</strong>osa<br />
Weraroa novae-zelandiae<br />
<strong>Psilocybe</strong> <strong>cyanescens</strong> (P21*) C. LSU D1 doma<strong>in</strong><br />
<strong>Psilocybe</strong> australiana Maximum parsimony<br />
<strong>Psilocybe</strong> <strong>cyanescens</strong><br />
<strong>Psilocybe</strong> serbica (P15)<br />
<strong>Psilocybe</strong> arcana (P05*)<br />
78<br />
<strong>Psilocybe</strong> moravica (P06)<br />
<strong>Psilocybe</strong> moravica var. sternberkiana (P07)<br />
85/dt<br />
<strong>Psilocybe</strong> australiana<br />
<strong>Psilocybe</strong> quebecensis<br />
<strong>Psilocybe</strong> subaerug<strong>in</strong>osa<br />
<strong>Psilocybe</strong> bohemica (P14)<br />
<strong>Psilocybe</strong> quebecensis<br />
65/51 98/- <strong>Psilocybe</strong> cubensis<br />
62/- <strong>Psilocybe</strong> subcubensis<br />
A. morphology tree<br />
97/97 <strong>Psilocybe</strong> cubensis<br />
<strong>Psilocybe</strong> cubensis<br />
bootstraps m<strong>in</strong>imum evolution/<br />
66/69 64/62 <strong>Psilocybe</strong> cubensis<br />
maximum parsimony<br />
<strong>Psilocybe</strong> subaerug<strong>in</strong>osa<br />
100/97 P. moravica<br />
<strong>Psilocybe</strong> semilanceata<br />
100/99 P. m. var. stenberkiana<br />
<strong>Psilocybe</strong> fimetaria<br />
73/dt P. bohemica<br />
<strong>Psilocybe</strong> l<strong>in</strong>iformans<br />
88/91 <strong>Psilocybe</strong> tampanensis<br />
P. serbica<br />
<strong>Psilocybe</strong> argentipes<br />
65/dt P. arcana<br />
99/98<br />
<strong>Psilocybe</strong> fasciata<br />
P. <strong>cyanescens</strong><br />
Panaleolus ulig<strong>in</strong>osus<br />
P. azurescens<br />
<strong>Psilocybe</strong> semilanceata<br />
P. cubensis<br />
0.1<br />
Stropharia semiglobata<br />
Hypholoma udum<br />
<strong>Psilocybe</strong> pseudobullacea<br />
P. bohemica<br />
Flammula alnicola<br />
Flammula alnicola<br />
72/81<br />
Hebeloma aff<strong>in</strong>e<br />
E. Elongation factor EF<br />
Maximum likelihood (nt)<br />
P.<br />
P.<br />
bohemica<br />
arcana<br />
0.01<br />
GTR model<br />
bootstraps<br />
ML/MP<br />
100/100<br />
D. ITS<br />
Maximum parsimony (nt)<br />
bootstraps<br />
MP/ML (GTR model)<br />
10<br />
75/83<br />
100/100<br />
100/100<br />
100/99<br />
89/93<br />
100/81<br />
91/100<br />
Galer<strong>in</strong>a badipes<br />
Galer<strong>in</strong>a atk<strong>in</strong>sonia<br />
<strong>Psilocybe</strong> moravica var. sternberkiana (P07)<br />
<strong>Psilocybe</strong> serbica (P15)<br />
<strong>Psilocybe</strong> arcana (P05*)<br />
<strong>Psilocybe</strong> arcana (P05)<br />
<strong>Psilocybe</strong> moravica (P06)<br />
<strong>Psilocybe</strong> bohemica (P14)<br />
<strong>Psilocybe</strong> <strong>cyanescens</strong> (P21)<br />
<strong>Psilocybe</strong> <strong>cyanescens</strong> (P02)<br />
<strong>Psilocybe</strong> <strong>cyanescens</strong><br />
<strong>Psilocybe</strong> azurescens (P01)<br />
<strong>Psilocybe</strong> semilanceata<br />
<strong>Psilocybe</strong> semilanceata<br />
<strong>Psilocybe</strong> semilanceata<br />
<strong>Psilocybe</strong> fasciata<br />
<strong>Psilocybe</strong> samuiensis<br />
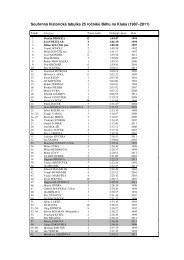
Fig. 1 Phylogenetic trees as <strong>in</strong>ferred from various molecular and<br />
morphological data. a M<strong>in</strong>imum evolution tree based on morphological,<br />
phenological and ecological data (bootstraps m<strong>in</strong>imum evolution/<br />
maximum parsimony; both 1,000 replicates). b Maximum likelihood<br />
tree (loglk=−2707.37145; gamma shape 0.015; proportion <strong>of</strong> <strong>in</strong>variant<br />
sites 0.000) based on partial LSU rDNA sequences. c Topology <strong>of</strong><br />
species <strong>of</strong> <strong>in</strong>terest as <strong>in</strong>ferred by maximum parsimony method. d<br />
azurescens, and group 2 encompass<strong>in</strong>g P. serbica, P.<br />
bohemica, P. arcana and P. moravica (<strong>in</strong>clud<strong>in</strong>g its var.<br />
sternbekiana). The latter group is very well supported by<br />
bootstrap values. Furthermore, the taxa <strong>of</strong> the second group<br />
do not differ among each other <strong>in</strong> the sequence <strong>of</strong> all three<br />
DNA markers. We have also analyzed concatenated <strong>in</strong>trons<br />
from EF1α and have obta<strong>in</strong>ed the same topology as from<br />
the cod<strong>in</strong>g sequence (not shown). The absence <strong>of</strong> DNA<br />
polymorphism among the taxa <strong>of</strong> the P. serbica <strong>complex</strong> <strong>in</strong><br />
0.1<br />
57/61<br />
70/63<br />
-/51<br />
67/100<br />
<strong>Psilocybe</strong> moravica var. sternberkiana (P07)<br />
<strong>Psilocybe</strong> bohemica (P14)<br />
<strong>Psilocybe</strong> moravica (P06)<br />
<strong>Psilocybe</strong> serbica (P15)<br />
<strong>Psilocybe</strong> arcana (P05*)<br />
<strong>Psilocybe</strong> <strong>cyanescens</strong> (P21)<br />
<strong>Psilocybe</strong> <strong>cyanescens</strong> (P02)<br />
100/100 <strong>Psilocybe</strong> azurescens (P01*)<br />
Asterophora parasitica<br />
Tephrocybe gibberosa<br />
Clitocybe nebularis<br />
Collybia tuberosa<br />
Anthracophyllum archeri<br />
Marasmius alliaceus<br />
Hygrophoropsis aurantiaca<br />
Hebeloma cyl<strong>in</strong>drosporum<br />
Phaeomarasmius proximans<br />
Mythicomyces corneipes<br />
Lyophyllum caerulescens<br />
Exidia glandulosa<br />
Amanita brunnescens<br />
Mycol Progress<br />
Maximum likelihood tree (loglk = −1938.37061; gamma shape 0.496;<br />
proportion <strong>of</strong> <strong>in</strong>variant sites 0.386) as <strong>in</strong>ferred from ITS1+5.8 S+ITS2<br />
sequences. e Maximum likelihood tree (loglk=−1693.32043; gamma<br />
shape 1.026; proportion <strong>of</strong> <strong>in</strong>variant sites 0.509) as <strong>in</strong>ferred from<br />
nucleotide sequences cod<strong>in</strong>g for elongation factor 1−α (EF1α).<br />
Numbers above branches <strong>in</strong>dicate ML/MP bootstraps (number <strong>of</strong><br />
replicates 300/1,000)<br />
studied markers <strong>in</strong>dicates that these taxa represent, <strong>in</strong> fact, a<br />
s<strong>in</strong>gle species despite their phenotypic variability.<br />
In general, molecular markers can successfully uncover<br />
the existence <strong>of</strong> cryptic species. Such species do not display<br />
morphological variation but can be well dist<strong>in</strong>guished<br />
thanks to the polymorphism <strong>of</strong> DNA markers (Geiser et<br />
al. 1998; Roy et al. 1998; Cruse et al. 2002). In our case,<br />
however, the opposite situation seems to be at hand: a<br />
group <strong>of</strong> morphologically dist<strong>in</strong>guishable fungal taxa that