

Reframing the 'Everything is everywhere'debate: evidence for high ...
Reframing the 'Everything is everywhere'debate: evidence for high ...
Reframing the 'Everything is everywhere'debate: evidence for high ...

Create successful ePaper yourself
Turn your PDF publications into a flip-book with our unique Google optimized e-Paper software.
60<br />
Aquat Microb Ecol 41: 55–65, 2005<br />
Table 3. Percent divergence within and between clades of Halteria grandinella<br />
and its relatives, calculated using uncorrected d<strong>is</strong>tances<br />
Clade a 1 2 3 4 5 6 7<br />
1 0.13<br />
2 2.93 0.25<br />
3 1.95 3.71 0.25<br />
4 6.69 7.31 6.49 0.39<br />
5 6.05 7.64 4.88 8.22 0.20<br />
6 5.07 5.88 4.10 7.22 5.23 0.41<br />
7 5.61 7.41 5.20 7.80 6.15 4.55 0.20<br />
a Descriptions of populations/clades refer to Table 1 and Fig. 1<br />
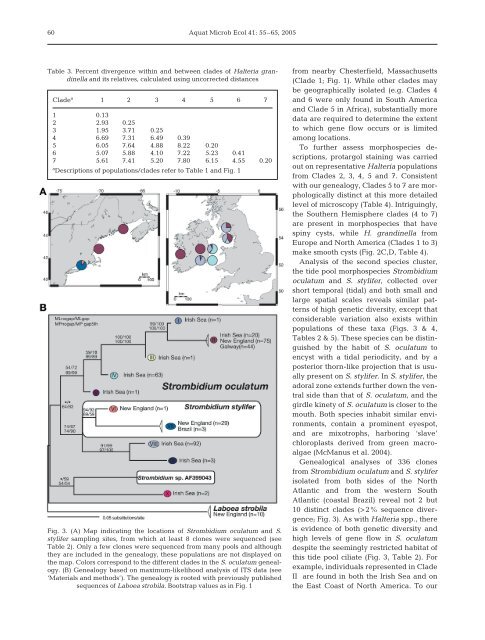
Fig. 3. (A) Map indicating <strong>the</strong> locations of Strombidium oculatum and S.<br />
stylifer sampling sites, from which at least 8 clones were sequenced (see<br />
Table 2). Only a few clones were sequenced from many pools and although<br />
<strong>the</strong>y are included in <strong>the</strong> genealogy, <strong>the</strong>se populations are not d<strong>is</strong>played on<br />
<strong>the</strong> map. Colors correspond to <strong>the</strong> different clades in <strong>the</strong> S. oculatum genealogy.<br />
(B) Genealogy based on maximum-likelihood analys<strong>is</strong> of ITS data (see<br />
‘Materials and methods’). The genealogy <strong>is</strong> rooted with previously publ<strong>is</strong>hed<br />
sequences of Laboea strobila. Bootstrap values as in Fig. 1<br />
from nearby Chesterfield, Massachusetts<br />
(Clade 1; Fig. 1). While o<strong>the</strong>r clades may<br />
be geographically <strong>is</strong>olated (e.g. Clades 4<br />
and 6 were only found in South America<br />
and Clade 5 in Africa), substantially more<br />
data are required to determine <strong>the</strong> extent<br />
to which gene flow occurs or <strong>is</strong> limited<br />
among locations.<br />
To fur<strong>the</strong>r assess morphospecies descriptions,<br />
protargol staining was carried<br />
out on representative Halteria populations<br />
from Clades 2, 3, 4, 5 and 7. Cons<strong>is</strong>tent<br />
with our genealogy, Clades 5 to 7 are morphologically<br />
d<strong>is</strong>tinct at th<strong>is</strong> more detailed<br />
level of microscopy (Table 4). Intriguingly,<br />
<strong>the</strong> Sou<strong>the</strong>rn Hem<strong>is</strong>phere clades (4 to 7)<br />
are present in morphospecies that have<br />
spiny cysts, while H. grandinella from<br />
Europe and North America (Clades 1 to 3)<br />
make smooth cysts (Fig. 2C,D, Table 4).<br />
Analys<strong>is</strong> of <strong>the</strong> second species cluster,<br />
<strong>the</strong> tide pool morphospecies Strombidium<br />
oculatum and S. stylifer, collected over<br />
short temporal (tidal) and both small and<br />
large spatial scales reveals similar patterns<br />
of <strong>high</strong> genetic diversity, except that<br />
considerable variation also ex<strong>is</strong>ts within<br />
populations of <strong>the</strong>se taxa (Figs. 3 & 4,<br />
Tables 2 & 5). These species can be d<strong>is</strong>tingu<strong>is</strong>hed<br />
by <strong>the</strong> habit of S. oculatum to<br />
encyst with a tidal periodicity, and by a<br />
posterior thorn-like projection that <strong>is</strong> usually<br />
present on S. stylifer. In S. stylifer, <strong>the</strong><br />
adoral zone extends fur<strong>the</strong>r down <strong>the</strong> ventral<br />
side than that of S. oculatum, and <strong>the</strong><br />
girdle kinety of S. oculatum <strong>is</strong> closer to <strong>the</strong><br />
mouth. Both species inhabit similar environments,<br />
contain a prominent eyespot,<br />
and are mixotrophs, harboring ‘slave’<br />
chloroplasts derived from green macroalgae<br />
(McManus et al. 2004).<br />
Genealogical analyses of 336 clones<br />
from Strombidium oculatum and S. stylifer<br />
<strong>is</strong>olated from both sides of <strong>the</strong> North<br />
Atlantic and from <strong>the</strong> western South<br />
Atlantic (coastal Brazil) reveal not 2 but<br />
10 d<strong>is</strong>tinct clades (>2% sequence divergence;<br />
Fig. 3). As with Halteria spp., <strong>the</strong>re<br />
<strong>is</strong> <strong>evidence</strong> of both genetic diversity and<br />
<strong>high</strong> levels of gene flow in S. oculatum<br />
despite <strong>the</strong> seemingly restricted habitat of<br />
th<strong>is</strong> tide pool ciliate (Fig. 3, Table 2). For<br />
example, individuals represented in Clade<br />
II are found in both <strong>the</strong> Ir<strong>is</strong>h Sea and on<br />
<strong>the</strong> East Coast of North America. To our













