The Role of Protein and Amino Acid Nutrition in ... - Adisseo.biz
The Role of Protein and Amino Acid Nutrition in ... - Adisseo.biz
The Role of Protein and Amino Acid Nutrition in ... - Adisseo.biz

You also want an ePaper? Increase the reach of your titles
YUMPU automatically turns print PDFs into web optimized ePapers that Google loves.
THE ROLE OF PROTEIN AND AMINO<br />
ACID NUTRITION ON REPRODUCTION<br />
OF DAIRY COWS<br />
José Eduardo P. Santos <strong>and</strong> Charles Staples<br />
Department <strong>of</strong> Animal Sciences<br />
University <strong>of</strong> Florida
<strong>Prote<strong>in</strong></strong> <strong>and</strong> Reproduction<br />
• Negative association between urea N<br />
concentration <strong>and</strong> pregnancy <strong>in</strong> dairy cows when<br />
urea N is excessive<br />
• Studies evaluat<strong>in</strong>g the effects <strong>of</strong> prote<strong>in</strong> on<br />
embryo quality <strong>and</strong> pregnancy usually fed cows<br />
diets with excessive concentrations <strong>of</strong> true<br />
prote<strong>in</strong> or urea
Influence <strong>of</strong> Dietary Ratio <strong>of</strong> DIP to UIP on MUN<br />
MUN, mg/100 ml<br />
20<br />
15<br />
10<br />
5<br />
5.6<br />
a<br />
11.6<br />
b<br />
13.4<br />
c<br />
14.4<br />
c<br />
17.8<br />
d<br />
JDS 76:525<br />
0<br />
80:80 100:100 120:80 100:120 120:120<br />
Intake <strong>of</strong> DIP & UIP as a % <strong>of</strong> NRC
Relationship <strong>of</strong> Bulk Tank MUN <strong>and</strong> Days to First AI<br />
Days to first AI<br />
140<br />
120<br />
100<br />
80<br />
60<br />
40<br />
20<br />
0<br />
Live. Prod. Sci. 37:91<br />
85<br />
78 78<br />
86<br />
103<br />
128<br />
10 12 14 16 18 20<br />
Milk Urea N, mg/100 ml
Overfeed<strong>in</strong>g <strong>Prote<strong>in</strong></strong> delays 1st Ovulation<br />
First ovulation, days<br />
60<br />
50<br />
40<br />
30<br />
20<br />
10<br />
0<br />
**<br />
25<br />
39<br />
** 50 * P = 0.10<br />
34<br />
22 26 17<br />
22<br />
Florida Utah Ma<strong>in</strong>e Ma<strong>in</strong>e<br />
*<br />
Control<br />
Excess DIP/CP<br />
** P = 0.05<br />
*
Overfeed<strong>in</strong>g <strong>Prote<strong>in</strong></strong> Delays Conception<br />
Days open<br />
120<br />
100<br />
80<br />
60<br />
40<br />
20<br />
0<br />
**<br />
96<br />
106<br />
* P = 0.10<br />
** P < 0.06<br />
** *<br />
72<br />
82<br />
Control<br />
Excess CP<br />
71 80<br />
Oregon '79 Ma<strong>in</strong>e '88 Ma<strong>in</strong>e '96
Effect <strong>of</strong> Dietary CP% on Fertility<br />
Concep/Preg Rate, %<br />
100<br />
90<br />
80<br />
70<br />
60<br />
50<br />
40<br />
30<br />
20<br />
10<br />
0<br />
Oregon<br />
*<br />
Israel<br />
Israel<br />
*<br />
OK<br />
Ma<strong>in</strong>e<br />
Israel<br />
*<br />
13-17% CP diets<br />
19-21% CP diets<br />
NY<br />
*<br />
NY<br />
* *<br />
Ma<strong>in</strong>e<br />
LA
Dietary CP% on Blood Urea N<br />
Reference Animal No. 13-17% CP 19-21% CP<br />
- - - - BUN, mg% - - - -<br />
Oregon, 1979<br />
Israel, 1981<br />
Israel, 1983<br />
OK, 1987<br />
Ma<strong>in</strong>e, 1988<br />
Israel, 1989<br />
NY, 1990<br />
NY, 1993<br />
Ma<strong>in</strong>e, 1996<br />
LA, 1999<br />
30<br />
39<br />
250<br />
109<br />
57<br />
139<br />
65<br />
80<br />
64<br />
119<br />
--<br />
9<br />
9<br />
15<br />
10<br />
25<br />
12<br />
14<br />
9<br />
20<br />
18<br />
15<br />
17<br />
25<br />
24<br />
32<br />
19<br />
24<br />
21<br />
25<br />
Average 14 22
MUN or PUN <strong>and</strong> Pregnancy per AI <strong>in</strong> Dairy Cows<br />
Adapted from Butler et al. J. Anim. Sci. (1996) <strong>and</strong> Ferguson et al. J. Dairy Sci. (1993)<br />
Pregnancy/AI, %<br />
55<br />
50<br />
45<br />
40<br />
35<br />
49.2<br />
320/650<br />
41.6<br />
119/286<br />
< 19 mg/dL<br />
> 19 mg/dL<br />
30
MUN <strong>and</strong> Predicted Pregnancy at 1 st AI <strong>in</strong><br />
Lactat<strong>in</strong>g Dairy Cows<br />
Negative association only <strong>in</strong> the first<br />
postpartum AI<br />
No effect on subsequent AI<br />
Guo et al. J. Dairy Sci. 87:1878–1885 (2004)
Potential Mechanisms for Reduced<br />
Fertility <strong>in</strong> Lactat<strong>in</strong>g Dairy Cows
Stages <strong>of</strong> the Reproductive Process Important for<br />
Establishment <strong>of</strong> Pregnancy <strong>in</strong> Cattle<br />
42 d 42 d<br />
Follicular development <strong>and</strong> oocyte growth<br />
Oocyte maturation<br />
Fertilization<br />
Preimplantation development<br />
IFN-τ<br />
x<br />
Prevention <strong>of</strong> Luteolysis<br />
PGF-2α
Effect <strong>of</strong> Dietary <strong>Prote<strong>in</strong></strong> on PUN <strong>and</strong> Uter<strong>in</strong>e<br />
pH on d 7 <strong>of</strong> the Estrous Cycle <strong>of</strong> Heifers<br />
P < 0.05 P < 0.05<br />
25<br />
7.2<br />
20<br />
7.1<br />
mg/dl<br />
15<br />
10<br />
pH<br />
7.0<br />
6.9<br />
6.8<br />
5<br />
6.7<br />
0<br />
PUN, mg/dl<br />
6.6<br />
Uter<strong>in</strong>e pH<br />
Balanced High RUP High RDP<br />
Balanced High RUP High RDP<br />
Elrod et al. (1993)
Effect <strong>of</strong> Dietary <strong>Prote<strong>in</strong></strong> on PUN <strong>and</strong><br />
Conception Rate (CR) <strong>in</strong> Heifers<br />
mg/dl<br />
25<br />
20<br />
15<br />
10<br />
5<br />
P < 0.05 P < 0.05<br />
%<br />
90<br />
80<br />
70<br />
60<br />
50<br />
40<br />
0<br />
Peak PUN<br />
30<br />
CR<br />
Balanced<br />
High RDP<br />
Balanced<br />
High RDP<br />
Elrod e Butler (1993)
Effect <strong>of</strong> Urea/Sal<strong>in</strong>e Infusion on Uter<strong>in</strong>e<br />
pH on d 7-8 <strong>of</strong> the Estrous Cycle<br />
7.4<br />
Uter<strong>in</strong>e pH<br />
PUN<br />
30<br />
Uter<strong>in</strong>e pH<br />
7.3<br />
7.2<br />
7.1<br />
7.0<br />
6.9<br />
6.8<br />
6.7<br />
25<br />
20<br />
15<br />
10<br />
PUN, mg/dL<br />
6.6<br />
6.5<br />
5<br />
6.4<br />
0 2 4 6 8 10 12 14 16 18 20 22 24 26 28 30 32 34 36 38 40 42 44 46 48<br />
Sal<strong>in</strong>e<br />
Urea<br />
Rhoads et al. J. Dairy Sci. (2004)
Effect <strong>of</strong> Urea Concentration on <strong>in</strong> Vitro Embryo Production<br />
Added to the oocyte maturation medium<br />
Added to the embryo culture medium<br />
Ocon <strong>and</strong> Hansen J. Dairy Sci. 86:1194-1200 (2003)
Effect <strong>of</strong> Dimethadione on pH <strong>and</strong> <strong>in</strong> Vitro Embryo Production<br />
Ocon <strong>and</strong> Hansen J. Dairy Sci. 86:1194-1200 (2003)
Pyruvate Metabolism <strong>of</strong> Ov<strong>in</strong>e Blastocyst Produced by<br />
IVF after Maturation with Granulosa Cells Previously<br />
Exposed to NH 4 Cl<br />
30<br />
Blastocyst<br />
P < 0.05 P = 0.06<br />
Cell<br />
0.6<br />
pMol/3h/blastocyst<br />
25<br />
20<br />
15<br />
0.5<br />
0.4<br />
0.3<br />
0.2<br />
pMol/3h/cell<br />
10<br />
0mM 5mM 10mM 0 Mm 5 mM 10 mM<br />
0.1<br />
NH4Cl (granulosa cells)<br />
Rooke et al. Anim. Reprod. Sci. (2000)
Effect <strong>of</strong> CP <strong>and</strong> RDP on Embryo Quality from<br />
Superovulated Cows<br />
CP (RDP)<br />
Cows<br />
Transferable Embryos<br />
Yes<br />
No<br />
Oocytes<br />
Transferable<br />
%<br />
Viable, vital<br />
sta<strong>in</strong><br />
%<br />
Reference<br />
12.3<br />
(59.7)<br />
27.4<br />
(70.7)<br />
22 4.0 1.6 1.8 49.7 53.1 b Garcia-Bojalil et al.<br />
(1994)<br />
22 4.9 2.0 1.8 54.0 66.7 a<br />
15.7<br />
(ND)<br />
21.9<br />
(ND)<br />
12 3.4 0.3 0.5 82.0 ND<br />
11 5.0 0.6 0.5 83.0 ND<br />
Rhoads et al. (2006)<br />
16.0<br />
(73.0)<br />
16.1<br />
(64.0)<br />
19 4.5 4.0 3.1 44.2 b ND<br />
Blanchard et al.<br />
(1990)<br />
19 5.5 3.3 2.3 66.9 a ND<br />
a, b P < 0.10
Effect <strong>of</strong> level <strong>of</strong> prote<strong>in</strong> feed<strong>in</strong>g on embryonic survival<br />
Rhoads et al. Anim. Reprod. Sci. 91: 1 (2006)<br />
15.5 mg/dL vs 24.4 mg/dL 7.7 mg/dL vs 25.2 mg/dL
<strong>Prote<strong>in</strong></strong> Can Also Influence<br />
Energy Metabolism
Effect <strong>of</strong> DIP & Fat on Reproduction<br />
<strong>of</strong> Dairy Cows (Garcia-Bojalil et al., 1998)<br />
• 45 multiparous Holste<strong>in</strong> cows<br />
• Dietary treatments were<br />
– 1) 11.1% DIP, 0% CaLCFA<br />
– 2) 11.1% DIP, 2.2% CaLCFA<br />
– 3) 15.7% DIP, 0% CaLCFA<br />
– 4) 15.7% DIP, 2.2% CaLCFA<br />
• Diets fed for the first 17 weeks PP
Effect <strong>of</strong> DIP & Fat on Reproduction<br />
<strong>of</strong> Dairy Cows (Garcia-Bojalil et al., 1998)<br />
Dietary <strong>Prote<strong>in</strong></strong> Sources<br />
• 11.1% DIP diets<br />
– Corn gluten meal<br />
– Fish meal<br />
– Blood meal<br />
– Meat <strong>and</strong> bone meal<br />
• 15.7% DIP diets<br />
– Soybean meal <strong>and</strong> urea
Effect <strong>of</strong> DIP <strong>and</strong> Fat on Body<br />
Weight Change <strong>of</strong> Lactat<strong>in</strong>g Cows<br />
Body weight, kg<br />
620<br />
600<br />
580<br />
560<br />
540<br />
520<br />
DIP; P = 0.01<br />
11.1% DIP<br />
" + fat<br />
15.7% DIP<br />
" + fat<br />
0 2 4 6 8 10 12 14<br />
Week <strong>of</strong> lactation
Effect <strong>of</strong> DIP <strong>and</strong> Fat on<br />
Accumulated Plasma Progesterone<br />
Progesterone, ng/ml<br />
70<br />
60<br />
50<br />
40<br />
30<br />
20<br />
10<br />
0<br />
DIP by FAT <strong>in</strong>terxn; P = 0.001<br />
11.1% DIP<br />
" + fat<br />
15.7% DIP<br />
" + fat<br />
1 8 15 22 29 36 43 50<br />
Day <strong>of</strong> lactation
Some <strong>Prote<strong>in</strong></strong> Sources Have<br />
Been L<strong>in</strong>ked with Improved<br />
Fertility
Pregnancy/Conception<br />
Rates Due to Feed<strong>in</strong>g Fish Meal<br />
% conception/pregnancy<br />
90<br />
80<br />
70<br />
60<br />
50<br />
40<br />
30<br />
20<br />
10<br />
0<br />
*<br />
**<br />
**P< 0.05; *P
What are Reasonable<br />
MUN or BUN Values?
Typical MUN Values <strong>of</strong> Cows Fed<br />
Diets Balanced for N <strong>and</strong> Energy<br />
Reference<br />
MUN, mg%<br />
Oltner <strong>and</strong> Wiktorsson, 1983 13.8<br />
Roseler et al., 1993 11.6<br />
Jonker et al., 1998 13.5
Urea<br />
Detoxification by liver<br />
Ammonia<br />
Deam<strong>in</strong>ation <strong>of</strong> <strong>Am<strong>in</strong>o</strong> <strong>Acid</strong>s<br />
Rumen<br />
Small<br />
Intest<strong>in</strong>e<br />
Muscle
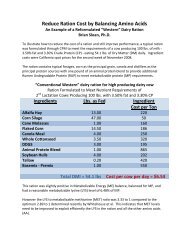
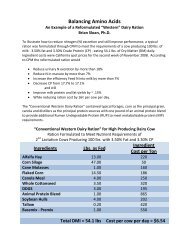
Diet Conta<strong>in</strong><strong>in</strong>g 17% CP <strong>and</strong> Balanced for Met<br />
(2.17% MP) & Lys (6.8% MP) Requirements<br />
• Lactat<strong>in</strong>g cow produc<strong>in</strong>g 45 kg <strong>of</strong> milk with 3.15%<br />
true prote<strong>in</strong> <strong>and</strong> consum<strong>in</strong>g 25 kg <strong>of</strong> DM<br />
– Metabolizable prote<strong>in</strong> required = 3,050 g/d<br />
– Metabolizable prote<strong>in</strong> <strong>in</strong>take = 3,150 g/d<br />
– Predicted PUN = 21 mg/dL<br />
3% surplus <strong>of</strong> MP<br />
• Lactat<strong>in</strong>g cow produc<strong>in</strong>g 20 kg <strong>of</strong> milk with 3.30%<br />
true prote<strong>in</strong> <strong>and</strong> consum<strong>in</strong>g 16 kg <strong>of</strong> DM<br />
– Metabolizable prote<strong>in</strong> required = 1,600 g/d<br />
– Metabolizable prote<strong>in</strong> <strong>in</strong>take = 1,935 g/d<br />
– Predicted PUN = 14 mg/dL<br />
20% surplus <strong>of</strong> MP
N Metabolism by Rumen Bacteria<br />
Peptides<br />
Peptides <strong>and</strong> AA<br />
Mono & Disaccharides<br />
<strong>Prote<strong>in</strong></strong>s<br />
deam<strong>in</strong>ase<br />
Peptides & AA<br />
Microbial cells<br />
& VFAs<br />
NH 3 + Keto-acid<br />
ATP<br />
ADP<br />
Portal<br />
System<br />
NH 3 + H NH 4<br />
Deam<strong>in</strong>ation <strong>of</strong> AA <strong>and</strong> NH 3<br />
production results from lack <strong>of</strong> ATP<br />
available for microbial growth
Effect <strong>of</strong> Dietary <strong>Prote<strong>in</strong></strong> Concentration <strong>and</strong><br />
Quality on Performance <strong>of</strong> Dairy Cows<br />
50<br />
45<br />
40<br />
LoCP = 17.0%<br />
HiCP = 18.3%<br />
40.8<br />
46.2<br />
b<br />
42.9<br />
46.6<br />
b<br />
kg/d<br />
35<br />
30<br />
a<br />
a<br />
25<br />
20<br />
15<br />
10<br />
a<br />
b b b<br />
HiCP-LoDRUP HiCP-HiDRUP LoCP-HiDRUP LoCP-<br />
HiDRUP+Met<br />
DM <strong>in</strong>take, kg/d Milk, kg/d N<strong>of</strong>tsger <strong>and</strong> St. Pierre J. Dairy Sci. (2003)
Excess <strong>of</strong><br />
dietary<br />
prote<strong>in</strong><br />
NH 3<br />
urea<br />
prote<strong>in</strong><br />
AA<br />
?<br />
NH 3 /NH 4<br />
+<br />
Urea + NH 3<br />
?<br />
Follicle<br />
development<br />
Toxic effects on<br />
oocytes <strong>and</strong><br />
granulosa cells<br />
Enhances<br />
blastocyst<br />
development<br />
Delays embryo<br />
transport<br />
Impairs early<br />
embryonic development
Summary - <strong>Prote<strong>in</strong></strong><br />
‣ Only excessive amounts <strong>of</strong> prote<strong>in</strong>/N might <strong>in</strong>fluence embryo<br />
quality <strong>and</strong> pregnancy<br />
alter the uter<strong>in</strong>e environment (pH)<br />
‣ Associated with <strong>in</strong>creased urea-N <strong>in</strong> reproductive tissues<br />
Lack <strong>of</strong> ATP for adequate microbial growth relative to N availability <strong>in</strong> the rumen<br />
‣ Recommendations<br />
Formulate diets based on metabolizable prote<strong>in</strong> needs <strong>of</strong> the<br />
animal<br />
Lactat<strong>in</strong>g cows: reduce the CP content to ~ 16 to 17% when<br />
high quality prote<strong>in</strong> sources are utilized <strong>and</strong> balance for<br />
am<strong>in</strong>o acids
<strong>Am<strong>in</strong>o</strong> <strong>Acid</strong>s <strong>and</strong> Reproduction<br />
• Bov<strong>in</strong>e conceptus requires am<strong>in</strong>o acids for development<br />
– Embryo growth<br />
– Placental development<br />
• Some am<strong>in</strong>o acids have physiological functions beyond<br />
build<strong>in</strong>g blocks for tissue deposition<br />
– Signal<strong>in</strong>g molecules (Arg<strong>in</strong><strong>in</strong>e -> NO, polyam<strong>in</strong>es)<br />
– Energy metabolism (Glyc<strong>in</strong>e)<br />
– Neurotransmiter (GABA)<br />
– DNA methylation (Methion<strong>in</strong>e)
Bov<strong>in</strong>e Peri-Implantation Events<br />
Conceptus<br />
position<br />
Conceptus<br />
Development<br />
Estradiol<br />
(pg/ml)<br />
10<br />
5<br />
Fimbria<br />
Oviduct<br />
Utero-tubal<br />
junction<br />
Shedd<strong>in</strong>g <strong>of</strong><br />
zona pellucida<br />
425µ<br />
150µ 170µ 205µ 340µ<br />
IFN τ production<br />
by trophoblast<br />
3<br />
mm<br />
Implantation<br />
250 mm<br />
Progesterone<br />
Estradiol<br />
6<br />
3<br />
Progesterone<br />
(ng/ml)<br />
0<br />
4<br />
8<br />
Days after mat<strong>in</strong>g<br />
12 16<br />
20
Some <strong>Am<strong>in</strong>o</strong> <strong>Acid</strong>s are Neurotransmiters<br />
‣ Aspartate, glyc<strong>in</strong>e, glutamate, <strong>and</strong> gamma am<strong>in</strong>obutyric acid<br />
(GABA)<br />
‣ B<strong>in</strong>d<strong>in</strong>g <strong>of</strong> N-Methyl-D-Aspartate to receptors results <strong>in</strong><br />
activation <strong>of</strong> ion channels <strong>in</strong> neurons <strong>and</strong> allows for entry <strong>of</strong> Na +<br />
<strong>and</strong> Ca ++ <strong>and</strong> exit <strong>of</strong> K +<br />
Stimulates the release <strong>of</strong> GnRH
Hormonal relationship between the hypothalamus,<br />
anterior pituitary <strong>and</strong> ovary<br />
Senger, 2003
DNA Methylation<br />
• Embryos up to the 8 cell stage:<br />
– DNA is de-methylated<br />
• 8 cell to Morula:<br />
– DNA is methylated<br />
– Modifies <strong>and</strong> add epigenetic <strong>in</strong>formation to the<br />
genome <strong>of</strong> the embryo. This process “epigenetic<br />
reprogram<strong>in</strong>g” is complete <strong>in</strong> the early embryonic<br />
stages
Methylation Pathways<br />
(DNA, prote<strong>in</strong>s, lipids)<br />
Phosphatidyl<br />
Chol<strong>in</strong>e<br />
Phosphatidil-<br />
Etanolam<strong>in</strong>e<br />
CH 3<br />
S-adenosyl-Met<br />
S-adenosylhomocyste<strong>in</strong>e<br />
CDP-Chol<strong>in</strong>e<br />
Dimethylglyc<strong>in</strong>e<br />
Met<br />
Tetrahydr<strong>of</strong>olate<br />
B 12<br />
Chol<strong>in</strong>e<br />
Beta<strong>in</strong>e<br />
Homocysthe<strong>in</strong>e<br />
CH 3 -Tetrahydr<strong>of</strong>olate<br />
Acetylchol<strong>in</strong>e<br />
Cyste<strong>in</strong>e<br />
Glutathione
<strong>Am<strong>in</strong>o</strong> <strong>Acid</strong> Concentrations <strong>in</strong> the Oviduct<br />
<strong>and</strong> Uterus <strong>of</strong> Dairy Heifers<br />
<strong>Am<strong>in</strong>o</strong> acid concentration, µM<br />
2,200<br />
2,000<br />
1,800<br />
1,600<br />
1,400<br />
1,200<br />
1,000<br />
800<br />
600<br />
400<br />
200<br />
0<br />
Oviduct<br />
Uterus<br />
Asp Glu Asn Ser His Gln Gly Thr Arg Tau Ala Tyr Met Trp Val Phe Ile Leu Lys<br />
Hugentobler et al. Mol. Reprod. Dev. 94: 445-454 (2007
Methion<strong>in</strong>e Concentrations <strong>in</strong> Different<br />
Body Compartments <strong>of</strong> Dairy Cows<br />
• Plasma:<br />
– 16 to 35 µMol/L<br />
• Uter<strong>in</strong>e fluid:<br />
– 31 to 46 µMol/L<br />
• Oviduct:<br />
– 31 to 49 µMol/L<br />
Hugentobler et al. Mol. Reprod. Dev. 94: 445-454 (2007
2 Experiments<br />
• Bov<strong>in</strong>e embryo development cultured <strong>in</strong> presence<br />
<strong>of</strong> different concentrations <strong>of</strong> methion<strong>in</strong>e<br />
– What is the adequate concentration <strong>in</strong> vitro for early<br />
embryo development?<br />
– Can embryo development be improve if<br />
concentrations are greater than normally found <strong>in</strong><br />
reproductive tissues?
Effect <strong>of</strong> Methion<strong>in</strong>e Concentration on <strong>in</strong> Vitro<br />
Development to Blastocyst <strong>in</strong> Cattle<br />
40<br />
35<br />
Day 7<br />
a,b<br />
P < 0.05<br />
40<br />
35<br />
b<br />
Day 8<br />
b<br />
b<br />
a,b<br />
P < 0.01<br />
b<br />
b<br />
%<br />
30<br />
25<br />
20<br />
15<br />
10<br />
a<br />
b<br />
b<br />
b<br />
b<br />
b<br />
%<br />
30<br />
25<br />
20<br />
15<br />
10<br />
a<br />
5<br />
5<br />
0<br />
0 35 50 100 200 400<br />
µM<br />
0<br />
0 35 50 100 200 400<br />
µM<br />
Bonilla et al. J. Dairy Sci. vol. 92, E-Suppl. 1 (Abstr.) pp. 69 (2009)
Effect <strong>of</strong> Methion<strong>in</strong>e Concentration on<br />
Blastocyst Cell Number<br />
250<br />
Blastocyst<br />
250<br />
Exp<strong>and</strong>ed Blastocyst<br />
200<br />
200<br />
Cell number<br />
150<br />
100<br />
Cell number<br />
150<br />
100<br />
50<br />
50<br />
0<br />
0 35 50 100 200 400<br />
µM<br />
0<br />
0 35 50 100 200 400<br />
µM<br />
Bonilla et al. J. Dairy Sci. vol. 92, E-Suppl. 1 (Abstr.) pp. 69 (2009)
Effect <strong>of</strong> Methion<strong>in</strong>e Concentration on <strong>in</strong> Vitro<br />
Development to Blastocyst <strong>in</strong> Cattle<br />
Blastocyst/oocyte, %<br />
50<br />
40<br />
30<br />
20<br />
10<br />
a<br />
b<br />
Day 7<br />
c<br />
c<br />
a,b, c<br />
P < 0.05<br />
bc<br />
bc<br />
Blastocyst/oocyte, %<br />
50<br />
40<br />
30<br />
20<br />
10<br />
a<br />
b<br />
Day 8<br />
b<br />
b<br />
a,b<br />
P = 0.01<br />
b<br />
b<br />
0<br />
0 7 14 21 28 35<br />
µM<br />
0<br />
0 7 14 21 28 35<br />
µM<br />
Bonilla et al. J. Dairy Sci. vol. 92, E-Suppl. 1 (Abstr.) pp. 69 (2009)
Effect <strong>of</strong> Methion<strong>in</strong>e Concentration on Glutathione<br />
Concentration <strong>and</strong> % <strong>of</strong> Advanced Blastocyst<br />
Glutathione, pMol on d 7<br />
0.8<br />
0.7<br />
0.6<br />
0.5<br />
0.4<br />
0.3<br />
0.2<br />
0.1<br />
0<br />
Glutathione on D 7<br />
a<br />
a,b<br />
P < 0.005<br />
b<br />
b<br />
b<br />
b<br />
b<br />
0 7 14 21 28 35<br />
µM<br />
Advanced Blastocyst, %<br />
90<br />
80<br />
70<br />
60<br />
50<br />
40<br />
30<br />
20<br />
10<br />
0<br />
Day 7<br />
a,b, c<br />
P < 0.001<br />
c<br />
bc bc<br />
bc<br />
b<br />
a<br />
0 7 14 21 28 35<br />
µM<br />
Bonilla et al. J. Dairy Sci. vol. 92, E-Suppl. 1 (Abstr.) pp. 69 (2009)
Metabolic fates <strong>of</strong> arg<strong>in</strong><strong>in</strong>e <strong>in</strong> mammalian cells<br />
Wu <strong>and</strong> Morris Jr. Biochem. J. (1998) 336, 1-17
NO <strong>and</strong> Polyam<strong>in</strong>es Enhance Angiogenesis <strong>and</strong><br />
are Potent Vasodilator <strong>in</strong> the Lungs <strong>and</strong> Placenta<br />
• Increase blood flow<br />
<strong>and</strong> nutrient supply<br />
to the fetus (Sladek<br />
et al., 1997)<br />
• Reverse some <strong>of</strong> the<br />
effects <strong>of</strong> <strong>in</strong>trauter<strong>in</strong>e<br />
growth<br />
retardation <strong>in</strong><br />
humans <strong>and</strong> sheep<br />
(de Boo et al. 2005;<br />
Thureen et al., 2002)
Approximate Embryonic <strong>and</strong> Fetal Losses<br />
When Calv<strong>in</strong>g Rate is 30%<br />
100<br />
80<br />
10-15%<br />
fertilization<br />
failure<br />
25% <strong>of</strong> the pregnancies<br />
are lost after placentation<br />
Two-cell Bov<strong>in</strong>e Embryo<br />
Embryo<br />
Cow<br />
Number<br />
60<br />
40<br />
20<br />
0<br />
SURVIVAL DEPENDS ON<br />
--Genetic <strong>in</strong>heritance<br />
--Non-genetic <strong>in</strong>heritance<br />
(from egg & sperm)<br />
--Environment<br />
57%<br />
early<br />
embryo<br />
mortality 15%<br />
late<br />
embryo<br />
mortality<br />
10%<br />
fetal loss<br />
AI Fertilization Day 28 Day 45 Term<br />
US<br />
Palpation
Calcium-Independent iNOS Activity <strong>in</strong> Mur<strong>in</strong>e Placenta<br />
on Different Days <strong>of</strong> Gestation<br />
Baylis et al. Mol. Human Reprod. vol.5 pp. 277–286, 1999
In Situ Hybridization for Inducible Nitric Oxide Synthase (iNOS)<br />
mRNA <strong>in</strong> Human Placenta<br />
A<br />
B<br />
A = sta<strong>in</strong><strong>in</strong>g <strong>in</strong> the syncytioblast <strong>and</strong> cytotrophoblast<br />
B = negative control<br />
C = light photomicroscopy <strong>of</strong> a placental arteriole<br />
D = dark photomicroscopy <strong>of</strong> a placental arteriole<br />
C<br />
D<br />
Baylis et al. Mol. Human Reprod. vol.5 pp. 277–286, 1999
In Situ Hybridization for Cationic AA Transporter <strong>in</strong><br />
Ov<strong>in</strong>e Endometrium
Effect <strong>of</strong> Daily Arg<strong>in</strong><strong>in</strong>e i.v. Infusion on Progesterone <strong>and</strong><br />
Ovarian Artery Resistance Index<br />
Progesterone, ng/mL<br />
9<br />
8<br />
7<br />
6<br />
5<br />
4<br />
3<br />
2<br />
AUC: P < 0.004<br />
Arg<strong>in</strong><strong>in</strong>e<br />
Control<br />
Resistance <strong>in</strong>dex<br />
0.6<br />
0.5<br />
0.4<br />
0.3<br />
0.2<br />
P < 0.05<br />
1<br />
0.1<br />
0<br />
0 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15<br />
Day <strong>of</strong> the cycle<br />
0<br />
Control<br />
Arg<strong>in</strong><strong>in</strong>e<br />
Arg<strong>in</strong><strong>in</strong>e <strong>in</strong>fused at 27 mg/kg <strong>of</strong> BW from estrus to d 15<br />
Luther et al. unpublished
Number <strong>of</strong> CL <strong>and</strong> Fetuses <strong>in</strong> Ewes Treated<br />
or Not with i.v. Arg<strong>in</strong><strong>in</strong>e<br />
Number per ewe<br />
2.0<br />
1.5<br />
1.0<br />
0.5<br />
Control<br />
Arg<strong>in</strong><strong>in</strong>e<br />
P < 0.05 P < 0.03<br />
0.0<br />
CL D 25 D 45<br />
Luther et al. unpublished
Number <strong>of</strong> CL <strong>and</strong> Fetuses <strong>in</strong> Sows Fed a Diet<br />
with 1% Arg<strong>in</strong><strong>in</strong>e<br />
Number or Kg<br />
18.0<br />
15.0<br />
12.0<br />
9.0<br />
6.0<br />
3.0<br />
Control<br />
Arg<strong>in</strong><strong>in</strong>e<br />
P < 0.05<br />
P < 0.05<br />
0.0<br />
Piglets/liter Live/liter Liter birth weight<br />
Mateo et al. J. Nutr. 137: 652-656 (2007)
Summary<br />
• Glyc<strong>in</strong>e is the ma<strong>in</strong> am<strong>in</strong>o acid <strong>in</strong> the reproductive tract <strong>of</strong><br />
bov<strong>in</strong>e<br />
– Energy source for the early embryo; synthesis <strong>of</strong> other AA; tissue deposition<br />
• Arg<strong>in</strong><strong>in</strong>e<br />
– Might mediate improvements <strong>in</strong> CL function <strong>and</strong> placentation through gaseous<br />
signal<strong>in</strong>g, angiogenesis <strong>and</strong> blood flow<br />
• Methion<strong>in</strong>e<br />
– Tissue deposition<br />
– Phospholipid synthesis<br />
– DNA methylation<br />
– Concentrations <strong>in</strong> the oviduct <strong>and</strong> endometrium seem adequate for optimum<br />
early embryo development
THANK YOU<br />
José Eduardo P. Santos<br />
Department <strong>of</strong> Animal Sciences<br />
University <strong>of</strong> Florida<br />
Jepsantos@ufl.edu