

Mode of phytochrome B action in the photoregulation of seed ...
Mode of phytochrome B action in the photoregulation of seed ...
Mode of phytochrome B action in the photoregulation of seed ...

You also want an ePaper? Increase the reach of your titles
YUMPU automatically turns print PDFs into web optimized ePapers that Google loves.
586 Tomoko Sh<strong>in</strong>omura et al.<br />
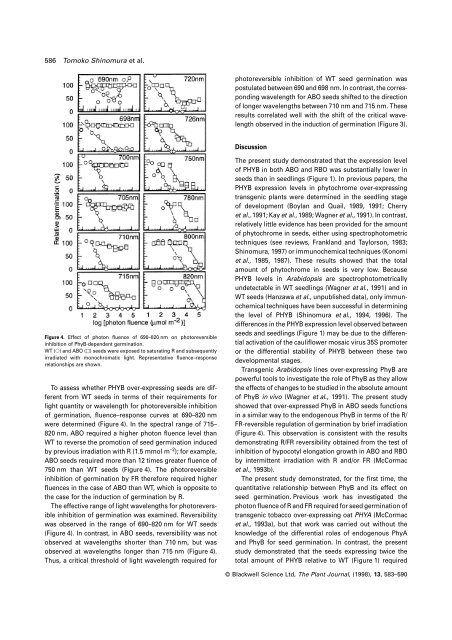
Figure 4. Effect <strong>of</strong> photon fluence <strong>of</strong> 690–820 nm on photoreversible<br />
<strong>in</strong>hibition <strong>of</strong> PhyB-dependent germ<strong>in</strong>ation.<br />
WT (s) and ABO (u) <strong>seed</strong>s were exposed to saturat<strong>in</strong>g R and subsequently<br />
irradiated with monochromatic light. Representative fluence–response<br />
relationships are shown.<br />
To assess whe<strong>the</strong>r PHYB over-express<strong>in</strong>g <strong>seed</strong>s are different<br />
from WT <strong>seed</strong>s <strong>in</strong> terms <strong>of</strong> <strong>the</strong>ir requirements for<br />
light quantity or wavelength for photoreversible <strong>in</strong>hibition<br />
<strong>of</strong> germ<strong>in</strong>ation, fluence–response curves at 690–820 nm<br />
were determ<strong>in</strong>ed (Figure 4). In <strong>the</strong> spectral range <strong>of</strong> 715–<br />
820 nm, ABO required a higher photon fluence level than<br />
WT to reverse <strong>the</strong> promotion <strong>of</strong> <strong>seed</strong> germ<strong>in</strong>ation <strong>in</strong>duced<br />
by previous irradiation with R (1.5 mmol m –2 ); for example,<br />
ABO <strong>seed</strong>s required more than 12 times greater fluence <strong>of</strong><br />
750 nm than WT <strong>seed</strong>s (Figure 4). The photoreversible<br />
<strong>in</strong>hibition <strong>of</strong> germ<strong>in</strong>ation by FR <strong>the</strong>refore required higher<br />
fluences <strong>in</strong> <strong>the</strong> case <strong>of</strong> ABO than WT, which is opposite to<br />
<strong>the</strong> case for <strong>the</strong> <strong>in</strong>duction <strong>of</strong> germ<strong>in</strong>ation by R.<br />
The effective range <strong>of</strong> light wavelengths for photoreversible<br />
<strong>in</strong>hibition <strong>of</strong> germ<strong>in</strong>ation was exam<strong>in</strong>ed. Reversibility<br />
was observed <strong>in</strong> <strong>the</strong> range <strong>of</strong> 690–820 nm for WT <strong>seed</strong>s<br />
(Figure 4). In contrast, <strong>in</strong> ABO <strong>seed</strong>s, reversibility was not<br />
observed at wavelengths shorter than 710 nm, but was<br />
observed at wavelengths longer than 715 nm (Figure 4).<br />
Thus, a critical threshold <strong>of</strong> light wavelength required for<br />
photoreversible <strong>in</strong>hibition <strong>of</strong> WT <strong>seed</strong> germ<strong>in</strong>ation was<br />
postulated between 690 and 698 nm. In contrast, <strong>the</strong> correspond<strong>in</strong>g<br />
wavelength for ABO <strong>seed</strong>s shifted to <strong>the</strong> direction<br />
<strong>of</strong> longer wavelengths between 710 nm and 715 nm. These<br />
results correlated well with <strong>the</strong> shift <strong>of</strong> <strong>the</strong> critical wavelength<br />
observed <strong>in</strong> <strong>the</strong> <strong>in</strong>duction <strong>of</strong> germ<strong>in</strong>ation (Figure 3).<br />
Discussion<br />
The present study demonstrated that <strong>the</strong> expression level<br />
<strong>of</strong> PHYB <strong>in</strong> both ABO and RBO was substantially lower <strong>in</strong><br />
<strong>seed</strong>s than <strong>in</strong> <strong>seed</strong>l<strong>in</strong>gs (Figure 1). In previous papers, <strong>the</strong><br />
PHYB expression levels <strong>in</strong> <strong>phytochrome</strong> over-express<strong>in</strong>g<br />
transgenic plants were determ<strong>in</strong>ed <strong>in</strong> <strong>the</strong> <strong>seed</strong>l<strong>in</strong>g stage<br />
<strong>of</strong> development (Boylan and Quail, 1989, 1991; Cherry<br />
et al., 1991; Kay et al., 1989; Wagner et al., 1991). In contrast,<br />
relatively little evidence has been provided for <strong>the</strong> amount<br />
<strong>of</strong> <strong>phytochrome</strong> <strong>in</strong> <strong>seed</strong>s, ei<strong>the</strong>r us<strong>in</strong>g spectrophotometric<br />
techniques (see reviews, Frankland and Taylorson, 1983;<br />
Sh<strong>in</strong>omura, 1997) or immunochemical techniques (Konomi<br />
et al., 1985, 1987). These results showed that <strong>the</strong> total<br />
amount <strong>of</strong> <strong>phytochrome</strong> <strong>in</strong> <strong>seed</strong>s is very low. Because<br />
PHYB levels <strong>in</strong> Arabidopsis are spectrophotometrically<br />
undetectable <strong>in</strong> WT <strong>seed</strong>l<strong>in</strong>gs (Wagner et al., 1991) and <strong>in</strong><br />
WT <strong>seed</strong>s (Hanzawa et al., unpublished data), only immunochemical<br />
techniques have been successful <strong>in</strong> determ<strong>in</strong><strong>in</strong>g<br />
<strong>the</strong> level <strong>of</strong> PHYB (Sh<strong>in</strong>omura et al., 1994, 1996). The<br />
differences <strong>in</strong> <strong>the</strong> PHYB expression level observed between<br />
<strong>seed</strong>s and <strong>seed</strong>l<strong>in</strong>gs (Figure 1) may be due to <strong>the</strong> differential<br />
activation <strong>of</strong> <strong>the</strong> cauliflower mosaic virus 35S promoter<br />
or <strong>the</strong> differential stability <strong>of</strong> PHYB between <strong>the</strong>se two<br />
developmental stages.<br />
Transgenic Arabidopsis l<strong>in</strong>es over-express<strong>in</strong>g PhyB are<br />
powerful tools to <strong>in</strong>vestigate <strong>the</strong> role <strong>of</strong> PhyB as <strong>the</strong>y allow<br />
<strong>the</strong> effects <strong>of</strong> changes to be studied <strong>in</strong> <strong>the</strong> absolute amount<br />
<strong>of</strong> PhyB <strong>in</strong> vivo (Wagner et al., 1991). The present study<br />
showed that over-expressed PhyB <strong>in</strong> ABO <strong>seed</strong>s functions<br />
<strong>in</strong> a similar way to <strong>the</strong> endogenous PhyB <strong>in</strong> terms <strong>of</strong> <strong>the</strong> R/<br />
FR-reversible regulation <strong>of</strong> germ<strong>in</strong>ation by brief irradiation<br />
(Figure 4). This observation is consistent with <strong>the</strong> results<br />
demonstrat<strong>in</strong>g R/FR reversibility obta<strong>in</strong>ed from <strong>the</strong> test <strong>of</strong><br />
<strong>in</strong>hibition <strong>of</strong> hypocotyl elongation growth <strong>in</strong> ABO and RBO<br />
by <strong>in</strong>termittent irradiation with R and/or FR (McCormac<br />
et al., 1993b).<br />
The present study demonstrated, for <strong>the</strong> first time, <strong>the</strong><br />
quantitative relationship between PhyB and its effect on<br />
<strong>seed</strong> germ<strong>in</strong>ation. Previous work has <strong>in</strong>vestigated <strong>the</strong><br />
photon fluence <strong>of</strong> R and FR required for <strong>seed</strong> germ<strong>in</strong>ation <strong>of</strong><br />
transgenic tobacco over-express<strong>in</strong>g oat PHYA (McCormac<br />
et al., 1993a), but that work was carried out without <strong>the</strong><br />
knowledge <strong>of</strong> <strong>the</strong> differential roles <strong>of</strong> endogenous PhyA<br />
and PhyB for <strong>seed</strong> germ<strong>in</strong>ation. In contrast, <strong>the</strong> present<br />
study demonstrated that <strong>the</strong> <strong>seed</strong>s express<strong>in</strong>g twice <strong>the</strong><br />
total amount <strong>of</strong> PHYB relative to WT (Figure 1) required<br />
© Blackwell Science Ltd, The Plant Journal, (1998), 13, 583–590










