Instituto de Neurobiología - Universidad Nacional Autónoma de ...
Instituto de Neurobiología - Universidad Nacional Autónoma de ...
Instituto de Neurobiología - Universidad Nacional Autónoma de ...

You also want an ePaper? Increase the reach of your titles
YUMPU automatically turns print PDFs into web optimized ePapers that Google loves.
<strong>Instituto</strong> <strong>de</strong> <strong>Neurobiología</strong>, UNAM Campus Juriquilla<br />
Jornadas 2009<br />
agitando las anémonas en agua <strong>de</strong>sionizada para <strong>de</strong>scargar los nematocistos; la mezcla<br />
fue centrifugada y liofilizado el sobrenadante (extracto crudo, EC). Se trabajó con un lote<br />
<strong>de</strong> 14 mg <strong>de</strong> proteínas totales <strong>de</strong>l EC y se <strong>de</strong>terminó que 2 μg <strong>de</strong> proteína producen 96%<br />
<strong>de</strong> hemólisis. La electroforesis <strong>de</strong>snaturalizante en gel <strong>de</strong> poliacrilamida (SDS-PAGE) <strong>de</strong>l<br />
extracto crudo mostró cinco bandas mayoritarias, cuyos pesos moleculares relativos fueron<br />
9, 12, 20, 36 y 125 kDa. El EC se fraccionó mediante una columna <strong>de</strong> exclusión molecular<br />
(Sepha<strong>de</strong>x G-50). Se obtuvieron 3 fracciones (F1, F2 y F3); la fracción F2 provocó 100%<br />
<strong>de</strong> hemólisis con 2 μg <strong>de</strong> proteína y presenta seis bandas mayoritarias <strong>de</strong> 9, 10, 12, 17,<br />
25 y 33 kDa. F2 se eluyó en una columna <strong>de</strong> Sepha<strong>de</strong>x G-20, se obtuvieron 3 fracciones<br />
(FI, FII y FIII), la fracción FII mostró 100% <strong>de</strong> actividad hemolítica con 2 μg <strong>de</strong> proteína y<br />
pesos moleculares relativos <strong>de</strong> 9, 12, 13, 15, 17 y 20 kDa. FII se eluyó en una columna <strong>de</strong><br />
fase inversa (C4), se obtuvieron seis fracciones. Al <strong>de</strong>terminar la actividad hemolítica en<br />
estas fracciones, ninguna provocó hemólisis; se concluyó que las toxinas pudieron haber<br />
sido inactivadas por los disolventes orgánicos utilizados. En posteriores experimentos la<br />
purificación se realizará mediante columnas <strong>de</strong> intercambio iónico.<br />
Estudiar el mecanismo <strong>de</strong> acción <strong>de</strong> estas toxinas podría proporcionar información sobre el<br />
efecto y función que tienen sobre el resto <strong>de</strong> los organismos marinos con los que interacciona<br />
esta anémona marina.<br />
Categoría T<br />
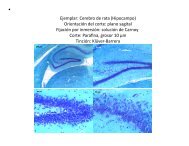
30. IDENTIFICACIÓN DE MARCADORES DE SUBPOBLACIONES DE CÉLULAS CAJAL-<br />
RETZIUS DURANTE EL DESARROLLO CORTICAL<br />
Fra<strong>de</strong> Pérez M. D., Miquelajáuregui Graf A. y Varela Echavarría A., Departamento <strong>de</strong><br />
<strong>Neurobiología</strong> <strong>de</strong>l Desarrollo y Neurofisiología. <strong>Instituto</strong> <strong>de</strong> <strong>Neurobiología</strong>, UNAM Campus<br />
Juriquilla<br />
Las células Cajal-Retzius (C-R) se localizan en la zona marginal <strong>de</strong> la corteza cerebral <strong>de</strong><br />
mamíferos. Estas células secretan Reelina, proteína requerida para la migración radial <strong>de</strong><br />
neuronas corticales en ruta a los diferentes estratos característicos <strong>de</strong> la corteza. La pérdida<br />
<strong>de</strong> la actividad <strong>de</strong> reelina ocasiona la inversión <strong>de</strong> los estratos corticales y en humanos se<br />
ha asociado a una forma congénita <strong>de</strong> lisencefalia.<br />
Los sitios <strong>de</strong> origen y rutas <strong>de</strong> migración <strong>de</strong> las células C-R no han sido dilucidados<br />
completamente. Existe evi<strong>de</strong>ncia que revela al “hem” cortical como la fuente principal <strong>de</strong><br />
células C-R pero otros sitios <strong>de</strong> origen han sido propuestos en los bor<strong>de</strong>s <strong>de</strong> la corteza en<br />
<strong>de</strong>sarrollo <strong>de</strong> don<strong>de</strong> se postula migran hasta ocupar la superficie cortical. Recientemente<br />
las rutas <strong>de</strong> migración <strong>de</strong> subpoblaciones <strong>de</strong> células C-R han sido estudiadas utilizando<br />
marcadores genéticos. Algunos estudios han relacionado la expresión génica con los sitios<br />
<strong>de</strong> generación llevando a la hipótesis <strong>de</strong> que las células C-R provenientes <strong>de</strong> diferentes<br />
centros <strong>de</strong> origen expresan perfiles genéticos y siguen rutas migratorias particulares.<br />
Con la finalidad <strong>de</strong> i<strong>de</strong>ntificar marcadores <strong>de</strong> las distintas subpoblaciones <strong>de</strong> células CR, en<br />
este trabajo se analizó mediante hibridación in situ en el telencéfalo <strong>de</strong> embriones <strong>de</strong> ratón<br />
un grupo <strong>de</strong> 10 genes que se ha propuesto se expresan en células C-R durante el <strong>de</strong>sarrollo<br />
embrionario (Rspo2, C6330 Rik, Sulf2, Acaa1b, Mab21l1, Scn3b, C6300 Rik, Hdac11, Tcf4 y<br />
Stmn4). Dos criterios importantes para la selección <strong>de</strong> marcadores moleculares <strong>de</strong> diferentes<br />
subpoblaciones <strong>de</strong> células C-R son expresión en los sitios propuestos <strong>de</strong> generación y<br />
también en la neocorteza, indicando posible migración celular.<br />
Los resultados obtenidos a la fecha indican que Rspo2 y Mab21l1 no parecen ser candidatos<br />
a<strong>de</strong>cuados, ya que no se expresan en la neocorteza, mientras que Scn3b cumple con<br />
ambos criterios.<br />
Septiembre 22