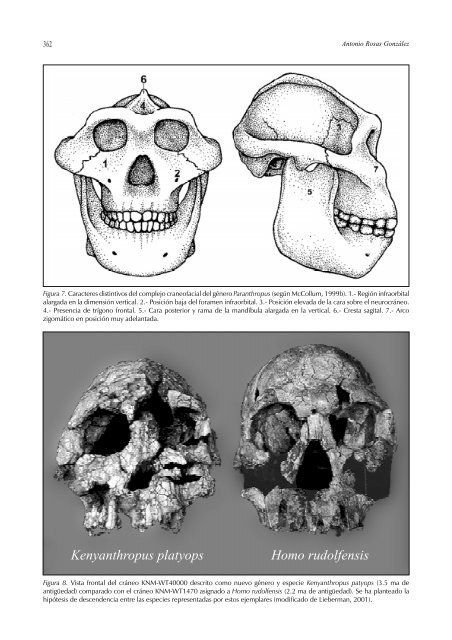
362 Antonio Rosas González Figura 7. Caracteres distintivos <strong>de</strong>l complejo craneofacial <strong>de</strong>l género Paranthropus (según McCollum, 1999b). 1.- Región infraorbital alargada <strong>en</strong> la dim<strong>en</strong>sión vertical. 2.- Posición baja <strong>de</strong>l foram<strong>en</strong> infraorbital. 3.- Posición <strong>el</strong>evada <strong>de</strong> la cara sobre <strong>el</strong> neurocráneo. 4.- Pres<strong>en</strong>cia <strong>de</strong> trígono frontal. 5.- Cara posterior y rama <strong>de</strong> la mandíbula alargada <strong>en</strong> la vertical. 6.- Cresta sagital. 7.- Arco zigomático <strong>en</strong> posición muy a<strong>de</strong>lantada. K<strong>en</strong>yanthropus platyops Homo rudolf<strong>en</strong>sis Figura 8. Vista frontal <strong>de</strong>l cráneo KNM-WT40000 <strong>de</strong>scrito como nuevo género y especie K<strong>en</strong>yanthropus patyops (3.5 ma <strong>de</strong> antigüedad) comparado con <strong>el</strong> cráneo KNM-WT1470 asignado a Homo rudolf<strong>en</strong>sis (2.2 ma <strong>de</strong> antigüedad). Se ha planteado la hipótesis <strong>de</strong> <strong>de</strong>sc<strong>en</strong><strong>de</strong>ncia <strong>en</strong>tre las especies repres<strong>en</strong>tadas por estos ejemplares (modificado <strong>de</strong> Lieberman, 2001).
CAPÍTULO 20: PAUTAS Y PROCESOS DE EVOLUCIÓN EN EL LINAJE HUMANO 363 estas formas <strong>de</strong>ntro <strong>de</strong>l género Paranthropus. El m<strong>en</strong>cionado género se caracteriza por una marcada megadontia post-canina y <strong>el</strong>evado <strong>de</strong>sarrollo <strong>de</strong>l esqu<strong>el</strong>eto facial (Fig. 7). La especie más antigua, P. aethiopicus, conserva aún características primitivas (por ejemplo, prognatismo ac<strong>en</strong>tuado y base <strong>de</strong>l cráneo ext<strong>en</strong>dida), si<strong>en</strong>do sus hipotéticas especies <strong>de</strong>sc<strong>en</strong>di<strong>en</strong>tes P. robustus <strong>en</strong> Sudáfrica, y P. boisei, <strong>en</strong> <strong>el</strong> este <strong>de</strong> Africa (con marcada flexión <strong>de</strong> la base <strong>de</strong>l cráneo). El grupo <strong>de</strong> los parántropos su<strong>el</strong>e consi<strong>de</strong>rarse como un grupo monofilético. Sin embargo, algunos autores han cuestionado esta interpretación alegando que los caracteres <strong>de</strong>rivados <strong>de</strong>l clado, muy <strong>en</strong> particular la megadontia post-canina, pue<strong>de</strong> ser converg<strong>en</strong>te. De acuerdo con esta última posibilidad, P. aethiopicus habría originado a P. boisei <strong>en</strong> <strong>el</strong> este <strong>de</strong> Africa, mi<strong>en</strong>tras que <strong>en</strong> Sudáfrica A. africanus habría dado orig<strong>en</strong> a la especie P. robustus. Obviam<strong>en</strong>te, <strong>en</strong> este caso <strong>el</strong> género Paranthropus no t<strong>en</strong>dría s<strong>en</strong>tido filog<strong>en</strong>ético. En opinión <strong>de</strong> McCollum (1999a) <strong>el</strong> patrón repetitivo <strong>de</strong> magadontia postcanina <strong>en</strong> homínidos ha frustrado los int<strong>en</strong>tos <strong>de</strong> reconstruir su historia filog<strong>en</strong>ética. El estudio <strong>de</strong> las formas robustas pone <strong>de</strong> manifiesto la aparición converg<strong>en</strong>te <strong>de</strong> características anatómicas usualm<strong>en</strong>te asociadas a adaptaciones <strong>de</strong>l aparato masticador a dietas duras. Por ejemplo, <strong>el</strong> cráneo KNM-WT 40000 reci<strong>en</strong>tem<strong>en</strong>te <strong>de</strong>scubierto y clasificado como nuevo género y especie K<strong>en</strong>yanthropus platyops parece atestiguar una cierta diversificación <strong>de</strong> los homínidos durante <strong>el</strong> Plioc<strong>en</strong>o medio. K. platyops pres<strong>en</strong>ta una combinación <strong>de</strong> caracteres primitivos y <strong>de</strong>rivados (Fig. 8). Entre los <strong>de</strong>rivados se manifiesta una cara aplanada con una reducción <strong>de</strong>l prognatismo <strong>de</strong> la región subnasal (orthognatia) pero estas características no se acompañan <strong>de</strong> megadontia post-canina. En <strong>el</strong> extremo contrario, una especie tardía <strong>de</strong>l género, Australopithecus garhi, aparece <strong>en</strong> <strong>el</strong> registro africano hace 2.5 ma, y manifiesta una peculiar combinación <strong>de</strong> caracteres (megadontia post-canina junto con di<strong>en</strong>tes anteriores también gran<strong>de</strong>s y marcado prognatismo pero sin hipertrofia <strong>de</strong> las superestructuras faciales). Se ha interpretado que pudiera tratarse <strong>de</strong>l antepasado directo <strong>de</strong>l género Homo (Asfaw et al. 1999). Un tema can<strong>de</strong>nte <strong>en</strong> <strong>el</strong> actual <strong>de</strong>bate <strong>de</strong> la evolución humana concierne al orig<strong>en</strong> y significado evolutivo <strong>de</strong>l género Homo. El género Homo y los problemas <strong>de</strong> las filog<strong>en</strong>ias morfológicas La evolución <strong>de</strong>l género Homo ha sido <strong>en</strong>t<strong>en</strong>dida hasta muy reci<strong>en</strong>tem<strong>en</strong>te como un proceso lineal (o anagénesis), mediante <strong>el</strong> cual nuestra especie se habría originado a través <strong>de</strong> tres fases sucesivas (Tobias, 1982). Según este mo<strong>de</strong>lo, la condición primitiva <strong>de</strong>l género la <strong>de</strong>fine la especie Homo habilis, <strong>de</strong> pequeño tamaño <strong>en</strong>cefálico y <strong>el</strong>evado <strong>de</strong>sarrollo <strong>de</strong> la cara. En calidad <strong>de</strong> intermedio se sitúa la especie Homo erectus, caracterizada por un increm<strong>en</strong>to <strong>de</strong>l <strong>en</strong>céfalo respecto a la anterior. Finalm<strong>en</strong>te, las difer<strong>en</strong>tes varieda<strong>de</strong>s <strong>de</strong> la especie Homo sapi<strong>en</strong>s, con un gran volum<strong>en</strong> <strong>en</strong>cefálico y una acusada reducción <strong>de</strong>l esplacnocráneo, serían los únicos repres<strong>en</strong>tantes vivos <strong>de</strong>l género Homo. Entre tanto, los conocidos Nean<strong>de</strong>rtales ocupaban una posición filog<strong>en</strong>ética incierta. La intrigante combinación <strong>de</strong> caracteres <strong>de</strong> los Nean<strong>de</strong>rtales, tales como su ac<strong>en</strong>tuado prognatismo mediofacial asociado a un volum<strong>en</strong> <strong>en</strong>cefálico similar al <strong>de</strong> nuestra especie, proporcionaba una <strong>de</strong> las incógnitas más persist<strong>en</strong>tes <strong>de</strong> la paleoantropología. Para algunos, los Nean<strong>de</strong>rtales formaban parte <strong>de</strong> la asc<strong>en</strong><strong>de</strong>ncia directa <strong>de</strong> los <strong>humano</strong>s mo<strong>de</strong>rnos (Coon 1939, Wolpoff 1980). Para otros, los Nean<strong>de</strong>rtales eran “algo” distinto a nuestra especie (Le Gros Clark 1964), lo que <strong>de</strong>jaba su orig<strong>en</strong> <strong>en</strong> un difuso limbo a la vez que hacía difícil explicar los parecidos con nuestra especie (p.ej. gran volum<strong>en</strong> <strong>en</strong>cefálico). D<strong>en</strong>tro <strong>de</strong> este esc<strong>en</strong>ario, <strong>el</strong> problema consistía <strong>en</strong> i<strong>de</strong>ntificar <strong>el</strong> límite inferior <strong>de</strong> nuestro propio género y <strong>el</strong> criterio utilizado se basaba <strong>en</strong> asignar a Homo toda especie asociada a instrum<strong>en</strong>tos líticos. Más reci<strong>en</strong>tem<strong>en</strong>te se ha utilizado también <strong>el</strong> volum<strong>en</strong> <strong>en</strong>cefálico como un criterio <strong>de</strong>terminante. Según estos criterios, los primeros repres<strong>en</strong>tantes <strong>de</strong>l género Homo hac<strong>en</strong> acto <strong>de</strong> pres<strong>en</strong>cia <strong>en</strong> torno a hace 2.5 ma, repres<strong>en</strong>tados por la especie H. rudolf<strong>en</strong>sis, y poco <strong>de</strong>spués se <strong>en</strong>cu<strong>en</strong>tran los restos <strong>de</strong> H. habilis (Fig. 9). En ambas especies se aprecia un significativo increm<strong>en</strong>to <strong>de</strong>l volum<strong>en</strong> <strong>en</strong>cefálico respecto a las especies <strong>de</strong>l género Australopithecus, aunque pres<strong>en</strong>tan <strong>en</strong>tre sí marcadas difer<strong>en</strong>cias <strong>en</strong> <strong>el</strong> tamaño y forma <strong>de</strong> la cara y la <strong>de</strong>ntición. Wood y Collard (1999) han pres<strong>en</strong>tado una propuesta <strong>de</strong> re<strong>de</strong>finición <strong>de</strong> lo que significa ser Homo. Dos principios inspiran esta nueva propuesta. Por un lado, la consi<strong>de</strong>ración <strong>de</strong> que un género <strong>de</strong>be ser un grupo monofilético; es <strong>de</strong>cir, todas sus especies <strong>de</strong>b<strong>en</strong> compartir un único antepasado común. Por otro, que sus especies <strong>de</strong>b<strong>en</strong> compartir un mismo régim<strong>en</strong> adaptativo. En opinión <strong>de</strong> estos autores, la inclusión <strong>de</strong> las especies H. habilis y H. rudolf<strong>en</strong>sis <strong>en</strong> <strong>el</strong> género Homo hace que no se cumplan ninguno <strong>de</strong> los dos requisitos. Según sus resultados H. habilis aparece (Fig. 2A) <strong>en</strong> una politomía no resu<strong>el</strong>ta junto con los parántropos (Fig. 2F). En la actualidad, difer<strong>en</strong>tes autores aceptan esta propuesta y ti<strong>en</strong><strong>de</strong>n a consi<strong>de</strong>rar a H. habilis como Australopithecus habilis (Collard y Ai<strong>el</strong>lo 2000). Lo mismo ocurre con H. rudolf<strong>en</strong>sis, cuya megadontia postcanina <strong>en</strong> ocasiones lo agrega al clado <strong>de</strong> los parántropos y <strong>de</strong>nota, <strong>en</strong> opinión <strong>de</strong> estos autores, un régim<strong>en</strong> adaptativo similar a éste. El reci<strong>en</strong>te <strong>de</strong>scubrimi<strong>en</strong>to <strong>de</strong> K. Platyops, <strong>de</strong> edad más antigua y morfología similar, hace p<strong>en</strong>sar <strong>en</strong> una posible r<strong>el</strong>ación <strong>de</strong> antepasado <strong>de</strong>sc<strong>en</strong>di<strong>en</strong>te <strong>en</strong>tre ambas especies. Así, H. rudolf<strong>en</strong>sis pasaría a ser K<strong>en</strong>yanthropus rudolf<strong>en</strong>sis (Fig. 8). Por tanto, según la re<strong>de</strong>finición propuesta, <strong>el</strong> género Homo queda restringido a las formas que compart<strong>en</strong> un orig<strong>en</strong> africano <strong>de</strong>s<strong>de</strong> hace unos 1.8 ma, con un claro increm<strong>en</strong>to <strong>de</strong>l <strong>en</strong>céfalo por <strong>en</strong>cima <strong>de</strong> los 800 cm 3 y una explotación similar <strong>de</strong> los recursos alim<strong>en</strong>ticios. Homo ergaster es la especie más antigua que cumple estos requisitos y es consi<strong>de</strong>rada, por lo tanto, la especie basal <strong>de</strong>l género. Algunos autores difer<strong>en</strong>ciaron a H. erectus como una especie