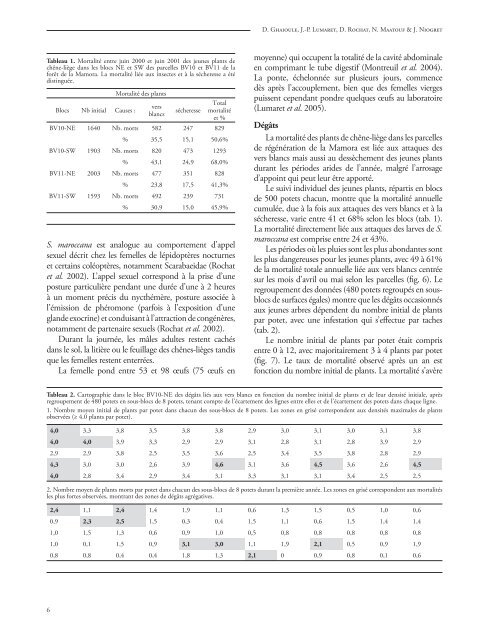
D. Ghaioule, J.-P. Lumaret, D. Rochat, N. Maatouf & J. NiogretTableau 1. Mortalité entre juin 2000 et juin 2001 <strong>des</strong> jeunes plants dechêne-liège dans <strong>les</strong> blocs NE et SW <strong>des</strong> <strong>par</strong>cel<strong>les</strong> BV10 et BV11 de laforêt de la Mamora. La mortalité liée aux insectes et à la sécheresse a étédistinguée.Blocs Nb initial Causes :Mortalité <strong>des</strong> plants<strong>vers</strong><strong>blancs</strong>sécheresseTotalmortalitéet %BV10-NE 1640 Nb. morts 582 247 829% 35,5 15,1 50,6%BV10-SW 1903 Nb. morts 820 473 1293% 43,1 24,9 68,0%BV11-NE 2003 Nb. morts 477 351 828% 23,8 17,5 41,3%BV11-SW 1593 Nb. morts 492 239 731% 30,9 15,0 45,9%S. maroccana est analogue au comportement d’appelsexuel décrit chez <strong>les</strong> femel<strong>les</strong> de lépidoptères nocturneset certains coléoptères, notamment Scarabaeidae (Rochatet al. 2002). L’appel sexuel correspond à la prise d’uneposture <strong>par</strong>ticulière pendant une durée d’une à 2 heuresà un moment précis du nycthémère, posture associée àl’émission de phéromone (<strong>par</strong>fois à l’exposition d’uneglande exocrine) et conduisant à l’attraction de congénères,notamment de <strong>par</strong>tenaire sexuels (Rochat et al. 2002).Durant la journée, <strong>les</strong> mâ<strong>les</strong> adultes restent cachésdans le sol, la litière ou le feuillage <strong>des</strong> chênes-lièges tandisque <strong>les</strong> femel<strong>les</strong> restent enterrées.La femelle pond entre 53 et 98 œufs (75 œufs enmoyenne) qui occupent la totalité de la cavité abdominaleen comprimant le tube digestif (Montreuil et al. 2004).La ponte, échelonnée sur plusieurs jours, commencedès après l’accouplement, bien que <strong>des</strong> femel<strong>les</strong> viergespuissent cependant pondre quelques œufs au laboratoire(Lumaret et al. 2005).DégâtsLa mortalité <strong>des</strong> plants de chêne-liège dans <strong>les</strong> <strong>par</strong>cel<strong>les</strong>de régénération de la Mamora est liée aux attaques <strong>des</strong><strong>vers</strong> <strong>blancs</strong> mais aussi au <strong>des</strong>sèchement <strong>des</strong> jeunes plantsdurant <strong>les</strong> pério<strong>des</strong> ari<strong>des</strong> de l’année, malgré l’arrosaged’appoint qui peut leur être apporté.Le suivi individuel <strong>des</strong> jeunes plants, ré<strong>par</strong>tis en blocsde 500 potets chacun, montre que la mortalité annuellecumulée, due à la fois aux attaques <strong>des</strong> <strong>vers</strong> <strong>blancs</strong> et à lasécheresse, varie entre 41 et 68% selon <strong>les</strong> blocs (tab. 1).La mortalité directement liée aux attaques <strong>des</strong> larves de S.maroccana est comprise entre 24 et 43%.Les pério<strong>des</strong> où <strong>les</strong> pluies sont <strong>les</strong> plus abondantes sont<strong>les</strong> plus dangereuses pour <strong>les</strong> jeunes plants, avec 49 à 61%de la mortalité totale annuelle liée aux <strong>vers</strong> <strong>blancs</strong> centréesur <strong>les</strong> mois d’avril ou mai selon <strong>les</strong> <strong>par</strong>cel<strong>les</strong> (fig. 6). Leregroupement <strong>des</strong> données (480 potets regroupés en sousblocsde surfaces éga<strong>les</strong>) montre que <strong>les</strong> <strong>dégâts</strong> occasionnésaux jeunes arbres dépendent du nombre initial de plants<strong>par</strong> potet, avec une infestation qui s’effectue <strong>par</strong> taches(tab. 2).Le nombre initial de plants <strong>par</strong> potet était comprisentre 0 à 12, avec majoritairement 3 à 4 plants <strong>par</strong> potet(fig. 7). Le taux de mortalité observé après un an estfonction du nombre initial de plants. La mortalité s’avèreTableau 2. Cartographie dans le bloc BV10-NE <strong>des</strong> <strong>dégâts</strong> liés aux <strong>vers</strong> <strong>blancs</strong> en fonction du nombre initial de plants et de leur densité initiale, aprèsregroupement de 480 potets en sous-blocs de 8 potets, tenant compte de l’écartement <strong>des</strong> lignes entre el<strong>les</strong> et de l’écartement <strong>des</strong> potets dans chaque ligne.1. Nombre moyen initial de plants <strong>par</strong> potet dans chacun <strong>des</strong> sous-blocs de 8 potets. Les zones en grisé correspondent aux densités maxima<strong>les</strong> de plantsobservées (≥ 4.0 plants <strong>par</strong> potet).4,0 3,3 3,8 3,5 3,8 3,8 2,9 3,0 3,1 3,0 3,1 3,84,0 4,0 3,9 3,3 2,9 2,9 3,1 2,8 3,1 2,8 3,9 2,92,9 2,9 3,8 2,5 3,5 3,6 2,5 3,4 3,5 3,8 2,8 2,94,3 3,0 3,0 2,6 3,9 4,6 3,1 3,6 4,5 3,6 2,6 4,54,0 2,8 3,4 2,9 3,4 3,1 3,3 3,1 3,1 3,4 2,5 2,52. Nombre moyen de plants morts <strong>par</strong> potet dans chacun <strong>des</strong> sous-blocs de 8 potets durant la première année. Les zones en grisé correspondent aux mortalités<strong>les</strong> plus fortes observées, montrant <strong>des</strong> zones de <strong>dégâts</strong> agrégatives.2,4 1,1 2,4 1,4 1,9 1,1 0,6 1,3 1,5 0,5 1,0 0,60,9 2,3 2,5 1,5 0,3 0,4 1,5 1,1 0,6 1,5 1,4 1,41,0 1,5 1,3 0,6 0,9 1,0 0,5 0,8 0,8 0,8 0,8 0,81,0 0,1 1,5 0,9 3,1 3,0 1,1 1,9 2,1 0,5 0,9 1,90,8 0,8 0,4 0,4 1,8 1,3 2,1 0 0,9 0,8 0,1 0,66
Dégâts <strong>par</strong> <strong>les</strong> <strong>vers</strong> <strong>blancs</strong> sur le chêne-liègeplus importante (≥ 50%) lorsque la densité initiale étaitsupérieure à 5 plants <strong>par</strong> potet. Les résultats présentésdans la figure 2 concernent le bloc BV10-SW, mais ceuxobtenus pour <strong>les</strong> 3 autres blocs analysés ne diffèrent passignificativement. Très vraisemblablement, une larvequi commence à commettre <strong>des</strong> <strong>dégâts</strong> dans un potetquelconque exploite la majorité <strong>des</strong> plants présents, lamortalité cumulée étant d’autant plus importante quele nombre initial de plants est élevé. La mortalité peutatteindre 70% <strong>des</strong> individus dans le cas de 10 plants misensemble dans le même potet. Il semble donc que la miseen place multiple de plants ou glands dans un même potet(<strong>des</strong>tinée en principe à pallier <strong>les</strong> risques liés à une mauvaisereprise ou germination) atteint ici une limite économique:plus <strong>les</strong> plants sont nombreux et plantés d’une manièredense (ce qui augmente le coût de la plantation), plus <strong>les</strong><strong>dégâts</strong> observés sont importants.Une étude antérieure réalisée en 1997 sur <strong>des</strong><strong>par</strong>cel<strong>les</strong> de régénération dans <strong>les</strong> cantons A, B et D soumisà un gradient croissant de continentalité avait montré quele taux moyen de mortalité <strong>des</strong> jeunes plants liée aux <strong>vers</strong><strong>blancs</strong> était respectivement de 14,2 et 13,0% (Rachdi &Haddan 1998). Nos observations effectuées en 2000 et2001 dans le canton B sont plus précises car <strong>les</strong> plantsont été suivis individuellement, en faisant la <strong>par</strong>t <strong>des</strong> <strong>vers</strong><strong>blancs</strong> <strong>par</strong> rapport à la mortalité globale. Les <strong>dégâts</strong> liés àceux-ci se situent dans une fourchette comprise entre 24 et43%. Cette différence <strong>par</strong> rapport aux travaux de Rachdi& Haddan (op. cit.) pourrait être liée à la pluviométrieenregistrée lors de l’année d’observation et à la densité<strong>des</strong> larves de grande taille (dernier stade) présentes dans<strong>les</strong> <strong>par</strong>cel<strong>les</strong>. Les attaques larvaires répétées entraînent<strong>des</strong> <strong>dégâts</strong> importants (mortalité) durant au moins <strong>les</strong> 4premières années de la vie <strong>des</strong> jeunes chênes-lièges, jusqu’àce que <strong>les</strong> plants atteignent une taille suffisante permettantleur survie (Rachdi & Haddan 1998; Lumaret et al.2005).Les <strong>dégâts</strong> liés aux <strong>vers</strong> <strong>blancs</strong> (sensu lato) sontcommuns dans de nombreuses régions du monde. EnFrance, <strong>les</strong> pullulations spectaculaires de Melolonthamelolontha L. sont anciennes; à <strong>par</strong>tir de 1935 el<strong>les</strong> avaiententraîné de gros dommages dans <strong>les</strong> prairies herbagères(Hurpin 1957) et <strong>les</strong> cultures comme celle de la pommede terre (Bedin 1982). En milieu forestier, Abgral (1991)rapporte que M. melolontha est capable de provoquer degros <strong>dégâts</strong> dans <strong>les</strong> pépinières et <strong>les</strong> jeunes plantations defeuillus et résineux. Une autre espèce, Melolontha papposaIlliger, cause <strong>des</strong> <strong>dégâts</strong> importants en Espagne dans <strong>les</strong>plantations d’olivier (Duran et al. 1996).Au Maroc, certains ravageurs <strong>des</strong> cultures ont faitl’objet d’étu<strong>des</strong> bioécologiques. Dans son inventaire<strong>des</strong> espèces de Scarabaeoidea, qui couvre une dizaine derégions du Maroc, Abaha (1991) a identifié 53 espècesdont 7 sont de véritab<strong>les</strong> ravageurs <strong>des</strong> cultures (tournesol,tomate, cultures fruitières et forestières). En forêt de laMamora, de nombreux <strong>vers</strong> <strong>blancs</strong> ont été signalés depuislongtemps sous <strong>les</strong> réserves et autour <strong>des</strong> souches <strong>des</strong> vieuxchênes-lièges morts (Marion 1955). Dans ce cas il s’agissaitplutôt de larves de Dynastidae inféodées aux vieux arbreset à la matière humique du sol. Par contre, <strong>les</strong> <strong>vers</strong> <strong>blancs</strong>présents un peu <strong>par</strong>tout dans <strong>les</strong> sols sablonneux <strong>des</strong><strong>par</strong>cel<strong>les</strong> de régénération de la Mamora ap<strong>par</strong>tiennentpour beaucoup à la famille <strong>des</strong> Melolonthidae; ces larvess’alimentent principalement <strong>des</strong> racines vivantes <strong>des</strong>graminées, <strong>des</strong> arbres et <strong>des</strong> arbustes.Isolement de la phéromone de Sphodroxia maroccanaL’identification de la phéromone sexuelle deS. maroccana est à replacer dans la perspective de disposerd’un outil d’avertissement qui permettrait de piéger <strong>les</strong>mâ<strong>les</strong>, de détecter leur présence et surtout d’estimer leurabondance. Il ne s’agit en aucun cas d’un moyen de luttedirecte du ravageur car <strong>les</strong> adultes, qui ne s’alimentent pas,ne représentent pas le stade ravageur; <strong>les</strong> femel<strong>les</strong> pourleur <strong>par</strong>t ne seraient pas piégées. Par contre, lors de l’étéqui précède <strong>les</strong> travaux de plantation dans une <strong>par</strong>celle derégénération de chêne-liège (travaux effectués en automneet en hiver), l’observation d’une très forte densité demâ<strong>les</strong> pourrait permettre de prendre localement certainesmesures préventives de protection <strong>des</strong> plants (<strong>par</strong> exempleutilisation d’un insecticide systémique en granulés aumoment de la plantation), sans que cette pratique onéreuseet polluante soit généralisée à toutes <strong>les</strong> <strong>par</strong>cel<strong>les</strong>.Pour l’instant l’extraction de la phéromone sexuelle deS. maroccana <strong>par</strong> solvants organiques ne s’est pas révéléeconcluante faute d’un matériel biologique suffisant au bonmoment et aucune différence consistante entre <strong>les</strong> extraitsn’a été observée entre mâ<strong>les</strong> et femel<strong>les</strong>, ni <strong>par</strong> rapport auxtémoins solvants. Lors de l’isolement <strong>par</strong> micro-extractionen phase solide, <strong>les</strong> contractions abdomina<strong>les</strong> de lafemelle associées à une large exposition de la membraneintersegmentaire entre ses segments abdominaux VIII etIX ont été observées à plusieurs reprises.Une quinzaine de composés volatils de faible poidsmoléculaire (PM