CICLO REPLICATIVO - Sezione di Microbiologia
CICLO REPLICATIVO - Sezione di Microbiologia
CICLO REPLICATIVO - Sezione di Microbiologia

You also want an ePaper? Increase the reach of your titles
YUMPU automatically turns print PDFs into web optimized ePapers that Google loves.
SCUOLA <strong>di</strong> MEDIDHE e FARMACEUTICHE<br />
2013<br />
REPLICAZIONE dei VIRUS<br />
Prof. Oliviero E. Varnier<br />
WWW.MICROBIOLOGIA.UNIGE.IT <strong>Sezione</strong> <strong>di</strong> <strong>Microbiologia</strong> – Dipartimento <strong>di</strong> Scienze Chirurgiche Diagnostiche Integrate (DISC)<br />
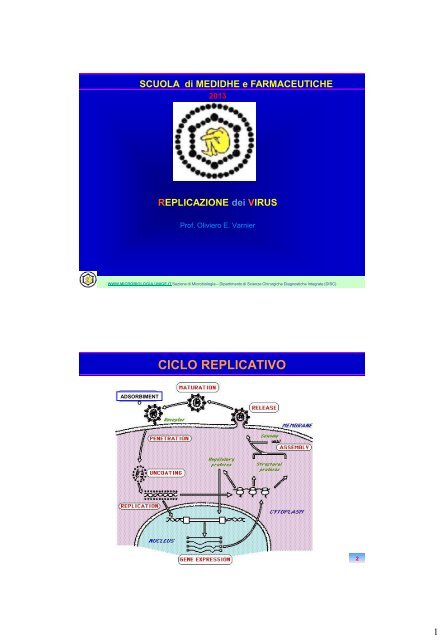
<strong>CICLO</strong> <strong>REPLICATIVO</strong><br />
ADSORBIMENT<br />
O<br />
2<br />
1
ADSORBIMENTO<br />
Alcuni virus richiedono, per il loro adsorbimento sulla<br />
cellula, l’interazione con molecole secondarie definite<br />
corecettori.<br />
HIV utilizza alcune proteine <strong>di</strong> trans-membrana,<br />
associate alle proteine G (proteine che usano il GDP<br />
per controllare il loro ciclo <strong>di</strong> “segnalazione”), che<br />
hanno come ligan<strong>di</strong> naturali le chemochine.<br />
3<br />
ADSORBIMENTO<br />
Gli antirecettori virali sono glicoproteine nei virus<br />
provvisti <strong>di</strong> involucro pericapsi<strong>di</strong>co o proteine del<br />
capside nei virus nu<strong>di</strong>.<br />
In molti virus con pericapside, gli antirecettori si<br />
presentano come proiezioni che sporgono da questa<br />
struttura;<br />
i virus nu<strong>di</strong> possono presentare gli antirecettori ubicati<br />
in rientranze che solcano la superficie del virione.<br />
4<br />
2
Adsorbimento<br />
Le interazioni che gli antirecettori virali instaurano<br />
con i recettori presenti sulla cellula ospite sono <strong>di</strong><br />
tipo sterico.<br />
La complementarietà spaziale permette la formazione<br />
<strong>di</strong> un certo numero <strong>di</strong> legami deboli che, nel loro<br />
insieme, portano all’adsorbimento stabile del virione<br />
alla cellula.<br />
Affinché ciò avvenga, è necessario un ambiente<br />
abbastanza ricco <strong>di</strong> ioni capaci <strong>di</strong> neutralizzare le<br />
cariche <strong>di</strong> superficie delle strutture complementari, in<br />
modo da ridurne la repulsione elettrostatica.<br />
5<br />
ADSORBIMENTO<br />
L’adsorbimento è un processo in<strong>di</strong>pendente dalla<br />
temperatura e non richiede l’intervento <strong>di</strong> reazioni che<br />
producono energia.<br />
Per molti virus nu<strong>di</strong> questa fase rappresenta un processo<br />
irreversibile poiché causa, nella struttura del<br />
virione, delle alterazioni che fanno perdere al virus la<br />
sua capacità infettante: è fondamentale che al processo<br />
<strong>di</strong> adsorbimento segua la fase <strong>di</strong> penetrazione.<br />
6<br />
3
ADSORBIMENTO<br />
I poliovirus, in seguito all’adsorbimento, perdono una<br />
proteina del capside <strong>di</strong>ventando così suscettibili alle<br />
ribonucleasi e proteasi cellulari.<br />
Talvolta, se all’adsorbimento non segue la penetrazione,<br />
i virus possono ristaccarsi integri e infettare altre<br />
cellule.<br />
È questo il caso <strong>di</strong> ortomixovirus e paramixovirus,<br />
sul cui pericapside si trovano, oltre agli antirecettori,<br />
anche varie molecole <strong>di</strong> un’altra glicoproteina dotata <strong>di</strong><br />
attività enzimatica capace <strong>di</strong> scindere il legame tra<br />
virus e cellula.<br />
7<br />
ADSORBIMENTO<br />
Oltre al meccanismo <strong>di</strong> entrata, me<strong>di</strong>ato dalla<br />
interazione recettore-antirecettore, alcuni virus posno<br />
entrare all’interno della cellula ospite dopo essersi<br />
complessati con anticorpi virus-specifici presenti in<br />
quantità “sub-neutralizzanti”.<br />
Il virus così rivestito può essere catturato dai recettori<br />
per le immunoglobuline presenti sulla superficie<br />
della cellula.<br />
Questo tipo <strong>di</strong> infezione, anticorpo-me<strong>di</strong>ata, è stato<br />
osservato per i virus West Nile, Dengue, influenza A<br />
e HIV-1.<br />
8<br />
4
ADSORBIMENTO<br />
L’esistenza <strong>di</strong> un meccanismo alternativo <strong>di</strong><br />
entrata suggerisce che, in assenza <strong>di</strong> una<br />
risposta immunitaria capace <strong>di</strong> produrre elevati<br />
titoli <strong>di</strong> anticorpi neutralizzanti…..<br />
….alcuni virus non richiedono la presenza <strong>di</strong><br />
un recettore cellulare per entrare all’interno<br />
della cellula ospite e sono potenzialmente<br />
capaci <strong>di</strong> allargare il proprio tropismo.<br />
9<br />
PENETRAZIONE<br />
Subito dopo l’adsorbimento, il virus penetra nella<br />
cellula attraverso <strong>di</strong>versi meccanismi che richiedono<br />
l’impiego <strong>di</strong> energia da parte della cellula.<br />
I virus con pericapside hanno un doppio strato<br />
fosfolipi<strong>di</strong>co che circonda il nucleocapside.<br />
L’entrata <strong>di</strong> questi virus all’interno della cellula<br />
prevede la fusione del pericapside virale con la<br />
membrana cellulare attraverso un processo me<strong>di</strong>ato<br />
dalle glicoproteine virali presenti sulla superficie del<br />
virione.<br />
10<br />
5
PENETRAZIONE<br />
Diversi virusi possiedono glicoproteine fusogene<br />
che facilitano questo processo.<br />
Fra queste, la glicoproteina gp41 <strong>di</strong> HIV, la proteina<br />
GP2 del virus Ebola e la proteina HA2 del virus<br />
influenzale, la quale viene attivata dal pH acido<br />
degli endosomi.<br />
Il virus influenzale, a <strong>di</strong>fferenza della maggior parte<br />
dei virus rivestiti, penetra me<strong>di</strong>ante un processo <strong>di</strong><br />
endocitosi, pertanto la fusione avviene con la<br />
membrana dell’endosoma.<br />
L’entrata dei virus nu<strong>di</strong> all’interno della cellula ospite<br />
avviene principalmente me<strong>di</strong>ante endocitosi me<strong>di</strong>ata<br />
da recettore.<br />
11<br />
ADSORBIMENTO<br />
12<br />
6
ADSORBIMENTO<br />
temperatura in<strong>di</strong>pendente<br />
richiede proteine d’adesione a proteine cellulari<br />
recettori cellulari<br />
13<br />
PENETRAZIONE DI VIRUS CON PERICAPSIDE<br />
Herpesvirus, paramyxovirus, HIV<br />
14<br />
7
PENETRAZIONE DI VIRUS CON PERICAPSIDE<br />
15<br />
Fusione<br />
con la<br />
membrana<br />
plasmatica<br />
fusione con le<br />
membrane<br />
endosomiche<br />
aci<strong>di</strong>che<br />
Entrata via endosomi<br />
16<br />
8
PENETRAZIONE DI VIRUS CON PERICAPSIDE<br />
from Schaechter et al, Mechanisms of Microbial Disease, 3rd ed, 1998<br />
17<br />
LA PENETRAZIONE DEL VIRUS VIA ENDOSOMI<br />
SINONIMI:<br />
VIROPESSI / ENDOCITOSI / PINOCITOSI<br />
18<br />
9
LA PENETRAZIONE DEI VIRUS SENZA PARICAPSIDE<br />
entrata <strong>di</strong>rettamente<br />
attraverso la membrana<br />
plasmatica<br />
19<br />
LA PENETRAZIONE DI VIRUS NUDI VIA ENDOSOMI<br />
Nel caso del poliovirus, un virus nudo appartenente<br />
alla famiglia Picornaviridae, è stato proposto un<br />
meccanismo <strong>di</strong> penetrazione alternativo che consiste<br />
nella traslocazione dell’RNA virale attraverso la<br />
membrana citoplasmatica.<br />
A seguito dell’adsorbimento del virus alla cellula<br />
ospite, una proteina virale andrebbe a formare un<br />
poro a livello della membrana cellulare attraverso il<br />
quale l’RNA virale verrebbe introdotto nel citoplasma.<br />
20<br />
10
LA PENETRAZIONE DI VIRUS NUDI VIA ENDOSOMI<br />
21<br />
ADSORBIMENTO, PENETRAZIONE E SCAPSIDAZIONE<br />
a) Adsorbimento b) Endocitosi c) Penetrazione d) scapsidazione<br />
22<br />
11
MECCANISMI DI ENTRATA<br />
Figura 33.1 Meccanismi <strong>di</strong> entrata per <strong>di</strong>verse classi <strong>di</strong> virus. I virus con pericapside<br />
possono entrare o me<strong>di</strong>ante fusione (1 e 2) oppure me<strong>di</strong>ante endocitosi seguita da un<br />
processo <strong>di</strong> fusione con la membrana endosomiale (3). I virus nu<strong>di</strong>, come i picornavirus<br />
23<br />
(4), penetrano all’interno della cellula me<strong>di</strong>ante un processo <strong>di</strong> endocitosi.<br />
SCAPSIDAZIONE<br />
Gli eventi che seguono l’entrata del virus e precedono<br />
la replicazione del genoma virale costituiscono il<br />
processo <strong>di</strong> scapsidazione, attraverso il quale il<br />
genoma virale si separa dalle proteine che lo<br />
rivestono.<br />
Questo evento, nel caso del poliovirus, potrebbe<br />
essere strettamente associato alla fase <strong>di</strong> penetrazione,<br />
dal momento che, potrebbe essere solo<br />
l’acido nucleico a traslocare all’interno del<br />
citoplasma.<br />
24<br />
12
SCAPSIDAZIONE<br />
Altri virus si ritrovano all’interno della cellula ospite<br />
come complesso nucleoproteico, e <strong>di</strong> conseguenza<br />
richiedono una fase <strong>di</strong> <strong>di</strong>sassemblaggio per liberare<br />
il proprio genoma.<br />
Per molti virus le proteine si <strong>di</strong>saggregano spontaneamente<br />
o per l’intervento <strong>di</strong> proteasi cellulari; in<br />
alcuni virus che penetrano per endocitosi, la<br />
scapsidazione può avvenire per azione <strong>di</strong> enzimi<br />
lisosomiali che si riversano all’interno dei vacuoli<br />
fagocitari.<br />
25<br />
SITI <strong>di</strong> REPLICAZIONE<br />
Dopo la fase <strong>di</strong> scapsidazione, il genoma virale può<br />
rimanere a livello del citoplasma o essere trasportato<br />
nel nucleo.<br />
La maggior parte dei virus a RNA si replicano nel<br />
citoplasma della cellula ospite, perché la loro replicazione,<br />
che avviene me<strong>di</strong>ante l’RNA polimerasi-RNA<br />
<strong>di</strong>pendente associata all’RNA virale, non richiede<br />
fattori nucleari.<br />
Al contrario, la maggior parte dei virus a DNA, i<br />
retrovirus, gli hepadnavirus e alcuni virus a RNA,<br />
come il virus influenzale, devono entrare nel nucleo<br />
della cellula poiché la loro replicazione richiede l’uso<br />
<strong>di</strong> enzimi e altri fattori che sono presenti nel nucleo<br />
della cellula.<br />
26<br />
13
SITI <strong>di</strong> REPLICAZIONE<br />
I virus che presentano una fase nucleare devono<br />
attraversare non solo la membrana plasmatica e/o<br />
endosomiale, ma debbono superare anche la membrana<br />
nucleare.<br />
Quest’ultima è costituita da una doppia membrana<br />
all’interno della quale si trova il complesso del poro,<br />
che presenta più <strong>di</strong> 50 <strong>di</strong>versi tipi <strong>di</strong> nucleoporine<br />
che regolano il trasporto <strong>di</strong> macromolecole verso<br />
l’interno ed esterno del nucleo.<br />
Questi meccanismi <strong>di</strong> trasporto, così come i segnali<br />
che li regolano, sono utilizzati da molti virus per far<br />
penetrare il proprio complesso ribonucleoproteico nel<br />
nucleo della cellula<br />
27<br />
SITI <strong>di</strong> REPLICAZIONE<br />
Particelle virali con massimo 39 nm <strong>di</strong> <strong>di</strong>ametro<br />
possono essere trasportate attraverso il complesso<br />
del poro.<br />
Virus e capsi<strong>di</strong> <strong>di</strong> piccole <strong>di</strong>mensioni possono<br />
teoricamente attraversare il complesso del poro<br />
senza essere necessariamente “<strong>di</strong>sassemblati” a<br />
livello citoplasmatico.<br />
Il parvovirus (<strong>di</strong>ametro 18-24 nm) e il capside del<br />
virus dell’epatite B (<strong>di</strong>ametro 36 nm) sono, con molta<br />
probabilità, importati intatti nel nucleo.<br />
28<br />
14
ESPRESSIONE E REPLICAZIONE DEL GENOMA<br />
Una volta che il genoma virale si è liberato del capside<br />
e ha raggiunto la propria sede, inizia la sintesi delle<br />
macromolecole virali.<br />
Il risultato fi nale <strong>di</strong> questo processo è la produzione <strong>di</strong><br />
numerose copie <strong>di</strong> componenti strutturali e genomici<br />
necessari per la formazione della nuova progenie.<br />
Per raggiungere questo obiettivo i virus devono sfruttare<br />
l’apparato biosintetico cellulare.<br />
29<br />
SINTESI PROTEICA<br />
La funzione che più <strong>di</strong>pende dalla cellula è la sintesi proteica,<br />
che è effettuata integralmente dagli apparati cellulari proteinosintetici.<br />
I virus devono presentare ai ribosomi cellulari i propri mRNA in<br />
modo tale che questi vengano riconosciuti e subito tradotti.<br />
La cellula impone delle limitazioni al virus, in quanto sintetizza i<br />
propri messaggeri solo nel nucleo e solo su stampi <strong>di</strong> DNA.<br />
I virus a DNA che sono in grado <strong>di</strong> raggiungere il nucleo possono<br />
sfruttare gli enzimi cellulari per produrre gli mRNA;<br />
gli altri virus presentano un genoma che può esso stesso<br />
funzionare da messaggero oppure debbono avere i propri enzimi<br />
capaci <strong>di</strong> sintetizzare gli mRNA.<br />
Inoltre l’apparato biosintetico cellulare può tradurre solo mRNA<br />
monocistronici, in quanto i ribosomi non sono in grado <strong>di</strong><br />
riconoscere segnali <strong>di</strong> inizio interni alla molecola <strong>di</strong> mRNA.<br />
30<br />
15
SINTESI PROTEICA<br />
La fase che più <strong>di</strong>pende dalla cellula ospite è la sintesi<br />
proteica, che viene effettuata integralmente dagli apparati<br />
proteinosintetici cellulari.<br />
I virus devono presentare ai ribosomi cellulari i propri<br />
mRNA in modo tale che questi vengano riconosciuti e<br />
tradotti.<br />
La cellula impone delle limitazioni al virus, in quanto<br />
sintetizza i propri messaggeri solo nel nucleo e solo su<br />
stampi <strong>di</strong> DNA:<br />
i virus a DNA che sono in grado <strong>di</strong> raggiungere il nucleo<br />
possono sfruttare gli enzimi cellulari per produrre mRNA;<br />
gli altri virus presentano un genoma «messaggero» o<br />
debbono avere i propri enzimi capaci <strong>di</strong> sintetizzare gli<br />
mRNA.<br />
31<br />
SINTESI PROTEICA<br />
Ne consegue che i virus debbono sintetizzare:<br />
un mRNA per ogni singola proteina, come avviene per il<br />
virus infl uenzale,<br />
un mRNA contenente l’informazione <strong>di</strong> <strong>di</strong>versi geni, che<br />
viene tradotto sotto forma <strong>di</strong> precursore poliproteico<br />
successivamente scisso da proteasi virali e/o cellulari<br />
nelle singole proteine.<br />
32<br />
16
ESPRESSIONE DEL GENOMA VIRALE<br />
Nella cellula infetta l’espressione del genoma virale è<br />
in competizione con una miriade <strong>di</strong> geni cellulari che<br />
devono essere trascritti e tradotti.<br />
Per poter superare questo ostacolo molti virus<br />
inibiscono la sintesi delle proteine cellulari<br />
<strong>di</strong>rettamente o in<strong>di</strong>rettamente.<br />
il virus dell’influenza blocca lo splicing e<br />
l’esportazione degli mRNA cellulari dal nucleo al<br />
citoplasma;<br />
altri virus, invece, producono un’enorme quantità <strong>di</strong><br />
messaggeri che sono favoriti nel processo <strong>di</strong><br />
traduzione.<br />
33<br />
Principali strategie replicative dei virus a RNA<br />
L’RNA virale monofilamento (ssRNA)<br />
a polarità positiva viene imme<strong>di</strong>atamente<br />
tradotto;<br />
l’RNA virale a polarità negativa<br />
necessita <strong>di</strong> essere trascritto da una<br />
RNA-polimerasi RNA <strong>di</strong>pendente.<br />
Nei retrovirus la trascrittasi inversa<br />
(RT) retrotrascrive l’RNA virale in una<br />
doppia catena <strong>di</strong> DNA (dsDNA).<br />
34<br />
17
REPLICAZIONE VIRALE: CLASSIFICAZIONE<br />
35<br />
18