lezione 3
lezione 3
lezione 3

You also want an ePaper? Increase the reach of your titles
YUMPU automatically turns print PDFs into web optimized ePapers that Google loves.
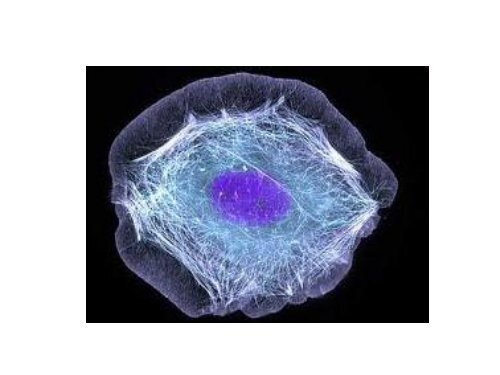
Per funzionare correttamente le cellule devono organizzarsi<br />
nello spazio e devono interagire meccanicamente con il loro<br />
ambiente.<br />
Devono, inoltre, avere una forma corretta, essere robuste e<br />
strutturate in modo corretto.<br />
Tali proprietà della cellula le sono conferite dal<br />
CITOSCHELETRO
Funzioni del CITOSCHELETRO<br />
Fornisce un supporto strutturale dinamico che determina la<br />
forma della cellula (es.: i microvilli di cellule epiteliali)<br />
Determina la posizione degli organelli citoplasmatici (es.<br />
polarità degli organelli nelle cellule ghiandolari esocrine)<br />
Direziona le molecole, gli organelli e le vescicole lungo specifici<br />
percorsi (es.: trasporto di vescicole di neurotrasmettitori lungo<br />
l’assone di una cellula nervosa<br />
Presiede al movimento cellulare (es.: Movimento flagellare di<br />
uno spermatozoo)<br />
Forma il macchinario per la divisione cellulare (fuso mitotico,<br />
anello contrattile della citodieresi)
CITOSCHELETRO<br />
Il citoscheletro è un sistema di filamenti proteici<br />
FILAMENTI INTERMEDI: forniscono forza meccanica<br />
MICROFILAMENTI: determinano la forma della superficie della<br />
cellula e sono necessari per la locomozione della cellula stessa<br />
MICROTUBULI: determinano le posizioni degli organelli e sono<br />
necessari per la locomozione della cellula<br />
PROTEINE ACCESSORIE: permettono l’assemblaggio regolato<br />
dei filamenti del citoscheletro, convertono l’energia dell’ATP in<br />
forza meccanica
Actina<br />
Actina-G (monomerica) ed Actina-F (Filamentosa)<br />
Actina lega l’ATP che durante la fase di polimerizzazione viene<br />
convertito in ADP
Proteine che legano l’Actina<br />
Nelle cellule i microfilamenti si organizzano in svariati modo, ad esempio in<br />
fasci, in complessi gelificati tridimensionali. La loro organizzazione è regolata<br />
da una miriade (oltre 100) di proteine che legano l’actina.<br />
Proteine di nucleazione (Arp, formina)<br />
La formina nuclea<br />
l’assemblaggio e resta<br />
associata all’estremità<br />
+ in crescita<br />
Il complesso Arp<br />
nuclea l’assemblaggio<br />
per formare una rete e<br />
resta associato alle<br />
estremità -
La polimerizzazione dell’actina è regolata da alcune proteine<br />
che legano l’Actina<br />
Timosina si lega alle G-Actina e ne impedisce l’assemblaggio<br />
Profilina si lega alle G-Actina e ne favorisce l’assemblaggio<br />
Cofilina si lega alla F-Actina e ne favorisce il disassemblaggio<br />
Gelsolina taglia i filamenti e si lega all’estremità +<br />
Proteina del cappuccio impedisce l’assemblaggio ed il<br />
disassemblaggio all’estremità +<br />
Tropomiosina lega e stabilizza il filamento
Disposizione dell’actina in una cellula<br />
I filopodi sono proiezioni appuntite della membrana plasmatica<br />
che permettono ad una cellula di esplorare il suo ambiente. Sono<br />
caratterizzati da stretti fasci paralleli di actina
Struttura di un microvillo<br />
E’ costituito da fasci stretti paralleli di<br />
actina uniti dalle proteine villina e<br />
fimbrina. Braccia laterali di miosina +<br />
calmodulina connettono i lati del fascio<br />
alla membrana plasmatica<br />
Microvillo<br />
delle cellule<br />
dell’epitelio di<br />
assorbimento<br />
dell’intestino
Microtubuli<br />
I microtubuli sono elementi cavi orientati con una estremità positiva e l’altra<br />
negativa. Sono polimeri di un eterodimero, l’ α- e β-tubulina. Il diametro<br />
esterno è di 25 nm, quello interno di 15 nm. In sezioni trasversali si osservano<br />
13 sub-unità. Pertanto i microtubuli sono costituiti da 13 protofilamenti<br />
affiancati.
I microtubuli sono nucleati da complessi contenenti la �-Tubulina<br />
I microtubli sno nucleati al livello del Centro Organizzatore dei Microtubuli<br />
(MTOC). I microtubuli sono nucleati al livello delle estremità -, con l’estremità<br />
+ che cresce verso l’esterno.
Il centrosoma è l’MTOC principale delle cellule animali<br />
Posto nel citoplasma vicino al nucleo,<br />
consiste di una matrice amorfa di<br />
proteine<br />
Immersa nel centrosoma si trova una<br />
coppia di strutture cilindriche,<br />
disposte fra loro ad angolo retto,<br />
chiamati centrioli
Attiva il<br />
disassemblaggio<br />
dei microtubuli<br />
all’estremità +<br />
Proteine associate con i microtubuli<br />
Taglia i microtubuli<br />
all’estremità +<br />
Fasci stretti Fasci a maglie<br />
larghe<br />
Legano e<br />
stabilizzano i<br />
microtubuli<br />
Legano l’estremità<br />
+ e ne accelerano<br />
l’assemblaggio
Ciglia e Flagelli<br />
I microtubuli sono alla base di due importanti sistemi di motilità cellulare: le<br />
ciglia e flagelli. Sono sottili espansioni della membrana plasmatica. Sono<br />
presenti in tutti gli organismi animali. Costituiscono il mezzo di locomozione<br />
di due importanti classi di protozoi, a cui devono il nome: i Ciliati e i<br />
Flagellati.<br />
Ciglia e Flagelli sono strutturalmente simili e differiscono per le dimensioni. I<br />
flagelli sono più grandi delle Ciglia<br />
Ciglia delle cellule<br />
dell’epitelio bronchiale<br />
Flagelli di organismi<br />
Flagellati
Struttura del Flagello<br />
La parte che emerge dal citoplasma è detto assonema, mentre<br />
la parte infissa nel citoplasma è detta corpuscolo basale o<br />
blefaroblasto.
Nell’assonema, perifericamente, sono presenti 9 coppie di microtubuli (A e B),<br />
legate tra loro da ponti di nexina. Dalla placca basale, che è un MTOC, emerge<br />
la coppia centrale dei microtubuli. La coppia centrale è connessa ai microtubuli<br />
periferici da fibre radiali.I microtubuli A delle 9 coppie esterne presentano due<br />
braccia di una proteina motrice la Dineina assonemale, che è alla base del<br />
movimento ciliare e flagellare.
Il battito ciliare dipende dalle braccia di dineina, le quali si attaccano al<br />
microtubulo B della coppia adiacente, idrolizzando ATP. Se i tubuli non<br />
fossero infissi nel citoplasma, si avrebbe lo scorrimento reciproco delle<br />
coppie. Essendo bloccati nel citoplasma il movimento di scorrimento si<br />
traduce in piegamento laterale, ovvero in battito ciliare. Cessato<br />
l’impulso il ciglio ritorna passivamente alla posizione iniziale.
I filamenti intermedi sono una componente abbondante del<br />
citoscheletro delle cellule animali. Si distinguono dai<br />
microfilamenti sottili e dai microtubuli per il loro spessore che di<br />
8-12 nm e pertanto intermedio tra quello dei microfilamenti di<br />
actina (6 nm) e dei microtubuli (25 nm)
I Filamenti intermedi costituiscono le lamine nucleari<br />
Le lamine nucleari presentano caratteristiche peculiari rispetto agli altri<br />
filamenti intermedi:<br />
non formano cordoni solidi, ma si assemblano a formare una solida rete,<br />
che riveste la membrana interna dei nuclei;<br />
sono presenti in tutte le cellule eucariotiche.