

AULA 6 Músculo Liso Biofísica Molecular 2012-2013.pdf
AULA 6 Músculo Liso Biofísica Molecular 2012-2013.pdf
AULA 6 Músculo Liso Biofísica Molecular 2012-2013.pdf

Create successful ePaper yourself
Turn your PDF publications into a flip-book with our unique Google optimized e-Paper software.
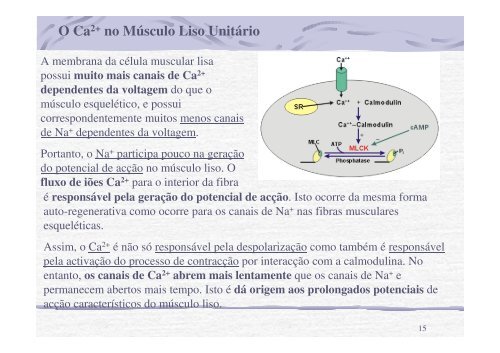
O Ca 2+ no <strong>Músculo</strong> <strong>Liso</strong> Unitário<br />
A membrana da célula muscular lisa<br />
possui muito mais canais de Ca 2+<br />
dependentes da voltagem do que o<br />
músculo esquelético, e possui<br />
correspondentemente muitos menos canais<br />
de Na + dependentes da voltagem.<br />
Portanto, o Na + participa pouco na geração<br />
do potencial de acção no músculo liso. O<br />
fluxo de iões Ca 2+ para o interior da fibra<br />
é responsável pela geração do potencial de acção. Isto ocorre da mesma forma<br />
auto-regenerativa como ocorre para os canais de Na + nas fibras musculares<br />
esqueléticas.<br />
Assim, o Ca 2+ é não só responsável pela despolarização como também é responsável<br />
pela activação do processo de contracção por interacção com a calmodulina. No<br />
entanto, os canais de Ca 2+ abrem mais lentamente que os canais de Na + e<br />
permanecem abertos mais tempo. Isto é dá origem aos prolongados potenciais de<br />
acção característicos do músculo liso.<br />
15
Potenciais no <strong>Músculo</strong> <strong>Liso</strong> Unitário<br />
No estado de repouso, o potencial intracelular<br />
da célula muscular lisa é de cerca de –50 a<br />
–60 mV, ou seja, cerca de –30 mV menos<br />
negativo que o da célula muscular<br />
esquelética.<br />
Os potenciais de acção no músculo liso<br />
unitário ocorrem em duas formas:<br />
A e B. Potencial em ponta (Spike Potentials)<br />
Este tipo de potenciais, que são similares aos<br />
que se observam no músculo esquelético,<br />
ocorrem na maioria das células do músculo<br />
liso unitário. Duram de cerca de 10 a 50 ms.<br />
Podem ser estimuladas de várias maneiras, por<br />
exemplo, por estimulação eléctrica, por acção<br />
de hormonas no músculo liso, por acção<br />
de neurotransmissores libertados pelas fibras nervosas, por estiramento, e ainda, como<br />
resultado da despolarização da fibra muscular mediada pelas células intersticiais de<br />
Cajal (células pacemaker).<br />
16
Potenciais no <strong>Músculo</strong> <strong>Liso</strong> Unitário<br />
B. Potencial de onda lenta (Slow Wave<br />
Potentials).<br />
O potencial de alguns tecidos musculares lisos<br />
oscila, formando uma onda lenta na zona<br />
adjacente à membrana plasmática, especialmente<br />
nos músculos lisos da parede dos intestinos. Esta<br />
onda lenta é produzida por uma rede especializada<br />
de células não musculares: as células intersticiais<br />
de Cajal que comunicam com o músculo liso<br />
através de junções de hiato. A onda lenta não é um<br />
potencial de acção. Ou seja, não é um processo<br />
auto-regenerativo que se propaga ao longo da<br />
fibra muscular. Mas quando a onda lenta é<br />
suficientemente positiva pode iniciar potenciais de<br />
acção no músculo liso, mediados pela abertura de<br />
canais iónicos de Ca 2+ dependentes da voltagem.<br />
As ondas lentas são também conhecidas por ondas<br />
marca-passo (pacemaker waves).<br />
17
Potenciais no <strong>Músculo</strong> <strong>Liso</strong> Unitário<br />
Figura do livro Integrative Action of the Autonomic Nervous System: Neurobiology of Homeostasis (2006) By Wilfrid<br />
Jänig, Cambridge University Press.
Potenciais no <strong>Músculo</strong> <strong>Liso</strong> Unitário<br />
C. Potencial de acção com plateau (Action<br />
Potentials with Plateaus)<br />
O começo deste tipo de potencial é similar ao<br />
potencial em ponta. No entanto, em vez de<br />
ocorrer uma rápida repolarização da célula, a<br />
repolarização atrasa-se várias centenas de ms<br />
(atraso que pode chegar até 1000 ms). Este<br />
plateau pode explicar as contracções<br />
prolongadas que ocorrem em alguns tipos de<br />
músculo liso, tal como o ureter, o útero e<br />
alguns tipos de músculos lisos vasculares.<br />
19
Potenciais no <strong>Músculo</strong> <strong>Liso</strong> Multiunitário<br />
As fibras musculares lisas multiunitárias contraem-se normalmente somente em<br />
resposta a estímulos nervosos (sistema nervoso autónomo).<br />
Não se desenvolvem tipicamente potenciais de acção. Isto deve-se ao facto das fibras<br />
serem constituídas por células muito pequenas que não estão conectadas umas às<br />
outras (o número de junções de hiato é muito reduzido ou nulo), pelo que cada célula<br />
contrái independentemente das restantes.<br />
O tempo requerido para<br />
activação da fibra é de cerca de<br />
200 a 300 ms, e chama-se<br />
período latente, que decorre<br />
antes da contracção começar.<br />
Este período latente é cerca de<br />
50× maior do que o período<br />
latente do músculo esquelético.<br />
20
Acção do Sistema Nervoso Autónomo no <strong>Músculo</strong> <strong>Liso</strong><br />
As fibras nervosas do sistema nervoso<br />
autónomo que inervam o músculo liso<br />
geralmente ramificam-se duma forma difusa no<br />
topo da camada de fibras musculares lisas.<br />
Na maioria dos casos, as fibras nervosas não<br />
fazem contacto directo com as fibras musculares<br />
lisas, formando junções difusas que secretam o<br />
seu neurotransmissor na matriz que rodeia o<br />
músculo liso, a alguns nm a µm das fibras. O<br />
neurotransmissor difunde-se até às células<br />
musculares.<br />
Quando existem muitas camadas de células musculares lisas, só a camada externa é que<br />
é inervada. A excitação muscular propaga-se entre as camadas por propagação interna<br />
do potencial de acção de uma célula muscular para outra e ainda por difusão externa do<br />
neurotransmissor.<br />
Os botões terminais das fibras nervosas têm múltiplas varicosidades distribuídas ao<br />
longo dos seus eixos. Nestes pontos, não existem células de Schwann, permitindo a<br />
secreção de NT através das paredes das varicosidades.<br />
21
Acção do Sistema Nervoso Autónomo no <strong>Músculo</strong> <strong>Liso</strong><br />
Em algumas situações, especialmente no músculo<br />
liso multiunitário, as varicosidades encontram-se<br />
directamente sobre a membrana da fibra<br />
muscular, separados desta membrana por cerca<br />
de 20 a 30 nm, a mesma espessura da fenda<br />
sináptica da junção neuromuscular. Estas zonas<br />
designam-se junções de contacto e funcionam da<br />
mesma forma que no músculo esquelético. Como<br />
consequência estas fibras contraem-se mais<br />
rapidamente do que as fibras estimuladas pelas<br />
junções difusas.<br />
Os neurotransmissores mais importantes secretados pelas fibras nervosas do<br />
sistema nervoso autónomo são a acetilcolina e a norepinefrina. Nunca são<br />
secretadas pelas mesmas fibras nervosas. A acetilcolina e a norepinefrina são<br />
substâncias excitatórias em alguns órgãos e inibitórias noutros órgãos.<br />
22
Acção do Sistema Nervoso Autónomo no <strong>Músculo</strong> <strong>Liso</strong><br />
Quando a acetilcolina excita uma fibra muscular, a<br />
norepinefrina tipicamente inibi-a, e vice-versa.<br />
Isto acontece porque a acetilcoolina e a<br />
norepinefrina actuam sobre as fibras musculares<br />
lisas por interacção com receptores proteicos<br />
membranares. Alguns destes receptores são<br />
excitatórios, enquanto que outros são inibitórios.<br />
Todos eles conduzem à activação de segundos<br />
mensageiros metabotrópicos.<br />
Exemplos da actuação de receptores muscarínicos<br />
(acetilcolina) (Eglen, 2006, Autonomic & Autacoid Pharmacology, 26: 219–233): M3 medeia a contracção de<br />
inúmeros músculos lisos nos tractos respiratório, gastrointestinal e genito-urinário; M1<br />
e M3 medeiam a relaxação do músculo liso vascular.<br />
Exemplos da actuação de receptores adrenérgicos (norepinefrina). Receptores α1,<br />
excitatórios, envolvidos na contracção do músculo liso vascular, tracto genito-urinário e<br />
esfíncteres e os receptores β2, com efeitos relaxantes no músculo liso dos tractos<br />
respiratório, gastrointestinal, e genito-urinário (Kandel et al, 2001, Principles of Neuroscience, NY: MacGraw-<br />
Hill).<br />
23
Estimulação mediada por hormonas<br />
Cerca de metade das contracções do músculo liso são iniciadas por factores<br />
estimulatórios que actuam directamente sobre a maquinaria de contracção do<br />
músculo liso, sem recurso a potenciais de acção. Os factores mais frequentemente<br />
envolvidos neste processo são: (1) hormonas e (2) agentes químicos locais.<br />
Muitas das hormonas em circulação no<br />
organismo afectam a contracção do músculo liso,<br />
em maior ou menor grau. São elas a<br />
norepinefrina, epinefrina, acetilcolina,<br />
angiotensina, endotelina, vasopressina, oxitocina,<br />
serotonina e histamina. Estas hormonas irão<br />
provocar excitação ou inibição da fibra muscular<br />
lisa dependendo da acção do receptor a que se<br />
ligam na célula.<br />
Os receptores hormonais apresentam dois tipos de resposta: (1) Alteram o<br />
potencial da célula, aumentam ou diminuem o potencial, por abertura e/ou fecho<br />
selectivo de canais iónicos; (2) Não alteram o potencial da célula, promovendo,<br />
por exemplo, a libertação de Ca<br />
24<br />
2+ do retículo sarcoplasmático (excitatório) ou a<br />
activação das Ca2+ Mg-ATPase que bombeiam iões cálcio para o retículo<br />
sarcoplasmático e espaço extracelular (inibitório).
Estimulação do músculo liso por neurotransmissores e hormonas<br />
Hilgers et al, 2005, Exp Biol Med, 230: 829-835.<br />
25
Relaxamento do músculo liso por neurotransmissores e hormonas<br />
Yuan et al, 2007, BMC Pregnancy and Childbirth, 7(Suppl 1):S10.<br />
26
Estimulação mediada por agentes químicos locais<br />
A contracção das arteríolas,<br />
metarteríolas e esfíncteres précapilares,<br />
é mediada pelo<br />
músculo liso. Estes pequenos<br />
vasos tem pouco ou nenhum<br />
controlo nervoso. No entanto,<br />
respondem rapidamente a<br />
alterações nas condições<br />
químicas locais do fluído<br />
intersticial. Alguns dos agentes<br />
químicos locais de controlo são:<br />
(1) O 2 - diminuição da concentração do O 2 nos tecidos locais faz com que o<br />
músculo liso relaxe, e portanto provoca vasodilatação.<br />
(2) CO 2 - excesso de CO 2 também provoca vasodilatação.<br />
(3) H + - aumento da concentração de H + também provoca vasodilatação.<br />
O aumento da concentração em adenosina, ácido láctico, iões K + , a diminuição da<br />
concentração em iões Ca 2+ e o aumento da temperatura corporal também<br />
provocam vasodilatação.<br />
27
Estimulação mediada por estiramento das membranas<br />
Quando o músculo liso é suficientemente estirado, gera potenciais de acção<br />
espontaneamente. Eles resultam duma combinação de: (1) potenciais de onda<br />
lenta e (2) despolarização membranar devida ao estiramento. Este tipo de<br />
resposta permite que os músculos lisos do intestino, quando suficientemente<br />
esticados por alimentos, se contraiam automática e ritmicamente.<br />
http://www.mfi.ku.dk/ppaulev/chapter22/kap%2022.htm<br />
Nota: Esta imagem é<br />
ilustrativa da acção do<br />
SNA e do estiramento<br />
nos movimentos<br />
peristálticos do<br />
intestino.<br />
28
Comparação dos mecanismos de contracção dos músculos<br />
esquelético e liso<br />
Para além dos aspectos, já referidos, sobre a acção do complexo Ca 2+ - calmodulina,<br />
da miosina cinase e da miosina fosfafatase no processo de contracção muscular,<br />
existem outras diferenças nos dois mecanismos de contracção muscular.<br />
A velocidade do ciclo das cabeças cruzadas é muito menor no músculo liso, cerca<br />
de 1/10 a 1/300 da frequência com que ocorre no músculo esquelético. Uma das razões<br />
para esta lentidão é a lenta actividade ATPásica das cabeças de miosina, que afecta a<br />
velocidade de degradação do ATP e consequentemente a velocidade de colocação das<br />
cabeças de miosina num estado conformacional de alta-energia. Por outro lado, a<br />
fracção de tempo em que as cabeças de miosina permanecem ligadas à actina é maior<br />
no músculo liso.<br />
A energia requerida para suster a contracção do músculo liso é cerca de 1/10 a 1/300<br />
da energia necessária para suster a mesma tensão de contracção no músculo<br />
esquelético. Pensa-se que isto resulta da lenta velocidade do ciclo das cabeças<br />
cruzadas e do facto de só se despender 1 molécula de ATP por ciclo,<br />
independentemente da sua duração. Este baixo dispêndio de energia é importante<br />
porque órgãos como os intestinos, a bexiga, a vesícula biliar, e outras vísceras mantém<br />
um estado de contracção tónica quase indefinidamente.<br />
29
Comparação dos mecanismos de contracção dos músculos<br />
esqueléticos e liso<br />
O começo da contracção e a duração total da contracção demoram em média<br />
mais tempo no músculo liso. O músculo liso começa a contrair cerca de 50 a 100<br />
ms após ter sido excitado, atinge a força máxima cerca de 500 ms depois,<br />
declinando de seguida até ao repouso em 1 a 2 s. O tempo total de contracção é de<br />
1 a 3 segs. Isto é em média 30× mais lento do que uma contracção dum músculo<br />
esquelético. Devido à grande variedade de músculos lisos, a contracção pode durar<br />
tão pouco como 0.2 segs ou tanto como 30 segs. São duas as razões para esta<br />
lentidão: (1) a baixa velocidade do ciclo das cabeças cruzadas; (2) a baixa<br />
velocidade do processo de iniciação da contracção.<br />
A força de contracção muscular é em média maior do que a do músculo<br />
esquelético – 4 a 6 Kg/cm 2 em comparação com o 3 a 4 Kg/cm 2 do músculo<br />
esquelético. Esta grande força de contracção resulta do período prolongado de<br />
ligação das cabeças cruzadas de miosina aos filamentos de actina.<br />
30
Comparação dos mecanismos de contracção dos músculos<br />
esqueléticos e liso<br />
O mecanismo latch permite que o<br />
músculo liso permaneça em contracção<br />
tónica durante horas, com pouco<br />
consumo de energia ou necessidade de<br />
sinais excitatórios.<br />
Quando a estimulação é persistente e<br />
os níveis citosólicos de Ca 2+<br />
permanecem relativamente elevados,<br />
a velocidade de hidrólise do ATP no<br />
ciclo das pontes cruzadas diminui.<br />
Nestas circunstâncias, a desfosforilação, mediada pela miosina fosfatase, das unidades<br />
regulatórias da miosina dá-se, mesmo com a miosina ligada à actina. As miosinas assim<br />
desfosforiladas permanecem muito mais tempo agarradas à actina, atrasando o ciclo da<br />
cabeças cruzadas enquanto mantém a tensão muscular. Como consequência, o número<br />
de cabeças ligadas aos filamentos de actina num dado momento permanece elevado,<br />
mantendo a contracção. Pouca energia está a ser utilizada nestas circunstâncias (cerca<br />
de 1/300 da energia que é utilizada para manter a contracção no músculo esquelético)<br />
porque o ATP não é degradado a ADP (Widmaier et al, 2006, Vander´s Human Physiology, McGraw-Hill).<br />
31
Bibliografia<br />
• Crimi E, Taccone FS, Infante T, Scolletta S, Crudele V, Napoli C (<strong>2012</strong>) Effects<br />
of intracellular acidosis on endothelial function: an overview J Crit Care 27: 108-<br />
118-<br />
• Guyton & Hall, 2005, Textbook of Medical Physiology, Pennsylvania: Elsevier.<br />
• Widmaier et al, 2006, Vander´s Human Physiology: The Mechanisms of Body<br />
Funcion, NY: McGraw-Hill.<br />
• Hilgers et al, 2005, <strong>Molecular</strong> Aspects of Arterial Smooth Muscle Contraction:<br />
Focus on Rho, Exp Biol Med, 230: 829-835.<br />
• Gunst, 2000, The contractile apparatus and mechanical properties of airway<br />
smooth muscle, Eur Respir J, 15: 600-616.<br />
32






