24-hour metabolism, Vad händer? Inledning
24-hour metabolism, Vad händer? Inledning
24-hour metabolism, Vad händer? Inledning

You also want an ePaper? Increase the reach of your titles
YUMPU automatically turns print PDFs into web optimized ePapers that Google loves.
2003-06-03<br />
<strong>24</strong>-<strong>hour</strong> Metabolism<br />
Ett arbete i Biokemi kursen vt. 2003<br />
Sofia Bertolino<br />
Annlouise Mickelsen<br />
Handledare: Tom Taylor<br />
Supervisor: Stefan Knight<br />
Inlämnat: Den 3 juni 2003
<strong>24</strong>-<strong>hour</strong> <strong>metabolism</strong>, <strong>Vad</strong> händer<br />
<strong>Inledning</strong><br />
Vi ska nu presentera ett arbete om fettsyror och dess <strong>metabolism</strong>. Vi kommer att ta upp<br />
hur fett bryts ner i kroppen och hur det syntetiseras. Nedbrytningen kan ta olika vägar<br />
beroende på hur de övriga näringsämnena i kroppen är balanserade. Vi kommer sedan<br />
att gå igenom de större övergripande stegen i <strong>metabolism</strong>en under ett dygn. Detta<br />
representeras av tre stadium the fed state, the early fasting state och the refed state. I den<br />
sistnämnda förklarar vi vad som händer i kroppen när magen får mat igen efter en natts<br />
fasta. Men vad händer om vi inte får mat igen Svar på detta ges vidare utifrån ett<br />
metaboliskt perspektiv. Arbetet avslutas med en beskrivning av ett huvudenzym i<br />
fettsyra <strong>metabolism</strong>en, vid namn Carnitin acetyltransferas, som hjälper till i<br />
nedbrytningen av fettsyror med långa kolkedjor.<br />
Bakgrund<br />
När vi till vardags pratar om fett är det oftast triacylglycerol vi menar. I detta arbetet<br />
kommer vi att benämna triacylglyceroler för ett lite enklare namn nämligen<br />
triglycerider. Byggstenarna i fett består av en glycerol molekyl och tre fettsyra<br />
molekyler. En vatten molekyl försvinner för varje fettsyra som binds till glycerolet. Fett<br />
används av kroppen som bla lagring av energi. För att komma åt fettet i fettcellerna och<br />
omvandla detta till energi, krävs vissa åtgärder. För det första måste fettet mobiliseras, i<br />
den här processen omvandlas triacylglycerol till glycerol och fria fettsyror, som sen<br />
frigörs från fettcellerna och transporteras till de energi krävande vävnaderna. Fettsyrorna<br />
måste också aktiveras och transporteras till mitokondrierna för vidare nedbrytning.<br />
Fettsyrorna bryts ner till acetyl CoA, som sen går in i citron syra cykeln.<br />
Bild 1. Schematisk bild på de olika reaktionsvägarna för fettsyror i muskler.<br />
Triglyceridens väg genom kroppen<br />
Fettet vi får i oss genom maten består ofta av triglycerider. Dessa hydrolyseras ner till<br />
glycerol och fria fettsyror, innan de kan absorberas av mag och tarmkanalen. När de väl
har blivit absorberade av tarmmembranen, syntetiseras dem igen och sedan böjar<br />
transporten av triglycerider till övriga celler i kroppen. Innan de tas upp av tex fett eller<br />
muskelvävnad måste de sönderdelas igen och sen sker transporten av dessa in till cellen.<br />
Även här sker en syntetisering av fettsyror och glycerol till triclycerider.<br />
Oxidation av fettsyror<br />
Det kan komma tider då kroppen behöver extra energi i form av fett. Musklerna oxiderar<br />
då dessa fettsyror. Denna process beskrivs vidare i oxidation av fettsyror.<br />
Fettsyrenedbrytningen sker i mitokondrierna och startar med en oxidation. Vid denna<br />
oxidation bildas en dubbelbindning mellan _- och _-kolatomerna. Coenzym i det enzym<br />
som katalyserar denna reaktion är FAD. Med hjälp av ett annat enzym adderas vatten till<br />
dubbelbindningen (hydratisering). Vattnets hydroxidgrupp binds till _-kolatomen och<br />
vätet till _-kolatomen. Nästa steg i nedbrytningen innebär ytterligare en oxidation med<br />
NAD + som coenzym, varvid syreatomen binds med en dubbelbindning till _-kolatomen.<br />
Slutprodukt vid _-oxidationen är en _-ketoförening av den aktiverade fettsyran, R-CH 2 -<br />
CO~S-CoA. Denna reagerar med ytterligare en molekyl coenzym A, varvid acetyl-CoA<br />
och en aktiverad fettsyra, två kolatomer kortare än den ursprungliga bildas. Denna<br />
avkortade fettsyra-CoA oxideras på nytt, reagerar med ännu en CoA-SH så att acetyl-<br />
CoA samt en ny avkortad fettsyra-CoA bildas osv tills hela fettsyran har överförts till<br />
acetyl-CoA. Den fortsatta nedbrytningen av acetyl-CoA sker i citronsyracykeln.<br />
Glycerol överförs genom en serie reaktioner till glyceraldehyd-3-fosfat. Nedbrytning av<br />
glyceraldehyd-3-fosfat fortsätter sedan i glykolysen.<br />
Biosyntes av fettsyror<br />
Biosyntesen av fettsyror sker i cytoplasman. Den är i princip omvändningen av _-<br />
oxidationen men katalyseras av helt andra enzymer. Fettsyresyntesen är en cyklisk<br />
process bestående av fyra reaktioner. Två av dessa är reduktioner och i båda fungerar<br />
coenzymet NADPH som reduktionsmedel. I fettsyresyntesen adderas två kol per varv,<br />
vilket förklarar varför de flesta biologiskt förekommande fettsyrorna innehåller ett jämnt<br />
antal kolatomer. Slutprodukten i fettsyresyntesen är palmitinsyra, som sedan kan<br />
förlängas i andra reaktionssekvenser. Även bildning av dubbelbindningar sker i andra<br />
processer, men vi människor kan inte bilda fleromättade fettsyror (essentiella). De<br />
syntetiserade fettsyrorna kan sedan komma att ingå antingen i neutrallipider eller i<br />
fosfolipider. I båda fallen sker en förestring av glycerol med fettsyrorna, och dessa<br />
reaktioner sker i endoplasmatiska nätverket.
<strong>24</strong>-<strong>hour</strong> <strong>metabolism</strong> cykeln.<br />
Under en dag ändras de biokemiska processerna flera gånger. Från nedbrytning<br />
till uppbyggning. De metaboliska vägarna inhiberas och aktiveras till följd av<br />
fysiologiska förhållanden. Vi ska nu gå igenom tre stora händelser som utspelas<br />
under ett dygn. Det börjar med the fed state. Detta representerar leverns respons<br />
på en intagen måltid. Vidare undernatten tar glukoslagren slut och levern byter<br />
”mode”. Detta representeras i the early fasting state. Åter på morgonen får magen<br />
i sig mat igen och detta representeras i The refed state.<br />
1.Well fed state.<br />
- Maten är klar ropar pappa från köket. Redan i hallen känner jag lukten av nybakad<br />
pizza. Ohh, calzone! Jag sätter mig vid bordet och framför mig ligger denna stora<br />
flottiga pizza som bara väntar på att få bli uppäten….<br />
Lukten, synen och smaken skickar signaler till parasympatiska nervsystemet om att det<br />
är dags att äta. Jag ska ta det lugnt, njuta.<br />
Den första tuggan är underbar. Proteinerna och glukosen transporteras sedan från magen<br />
till blodet. När glukosen närmar sig bukspottkörteln stimuleras speciella B- celler. Dessa<br />
celler börjar utsöndra hormonet insulin. Insulin signalerar till kroppen att den ska lagra<br />
glukos och bygga upp olika proteiner.<br />
Levern hjälper till att sänka halten glukos i blodet när glukoshalten är hög, genom att<br />
absorbera glukos. Glukosen lagras sedan i levern i form utav glykogen, för användning<br />
när kroppen är i behov av glukos.<br />
Insulin har också en generellt stimulerande effekt på proteinsyntesen. Detta i sin tur<br />
skapar gynnsammare förutsättningar för uppbyggnaden av muskelprotein.<br />
Alltså, Efter en måltid är kroppens huvudsakliga mål att lagra glukos i form av glykogen<br />
och upptaget av aminosyror för uppbyggnad av protein.<br />
2. The early fasting state.<br />
Några timmar efter en måltid sänks nivån glukos i blodet. Detta leder i sin tur till att<br />
utsöndringen av insulin avtar. Den låga nivån av glukos i blodet aktiverar a-celler i<br />
bukspottkörteln och utsöndrar istället ett annat hormon vid namn glukagon. Glukagon<br />
stimulerar glykogen nedbrytning och inhiberar glykogensyntesen.<br />
Glukagon inhiberar också fettsyrasyntesen genom att minska produktionen av pyruvat<br />
och genom att sänka aktiviteten av Acetyl CoA carboxylase.<br />
Glukagon stimulerar glukoneogenes i levern och blockerar glykolysen.<br />
Den stora mängden glukos som är formad genom hydrolys av glukos-6-fosfat från<br />
glykogen tas upp av blodet. Upptaget av glukos till muskel- och fett vävnad sänks som<br />
ett svar till den sjunkande insulinnivån.<br />
Huvud målet för glukagon är att öka utsöndringen av glukos från levern.<br />
När blodglukosnivån sänks använder muskler och levern fettsyror för att få energi. Pga<br />
detta borde nivån glukos i blodet hållas över 80 mg/dl. Detta görs av tre huvudfaktorer :
1. mobiliseringen av glykogen och utsöndringen av glukos i levern, 2, utsöndring av<br />
fettsyror från fettvävnad, 3, muskler och levern utvinner energi från fettsyror istället för<br />
glukos.<br />
När glukosen tar slut i levern, aktiveras glukoneogenesen. Denna är i behov av en<br />
kolkälla som kan förse syntesen av glukos med kol. En av dessa kol källor är glycerol<br />
som är produkt vid lipolysen. Resten av kolet kommer från hydrolysen av muskel<br />
protein.<br />
Alltså, När blodsocker nivån sänks, utsöndrar bukspottkörteln glukagon. Detta hormon<br />
har som uppgift att öka utsöndringen av glukos i levern. Fettsyror utsöndras från<br />
fettvävnad och en ändring i energi utvinning i muskler och lever sker, från glukos till<br />
fettsyror.<br />
3. The refed state.<br />
Denna fas representerar frukosten efter en natts fasta.<br />
Fett lagras som det gör i the fed state, men detta gäller inte för glukos. Levern börjar inte<br />
att uppta glukos från blodet, utan lämnar glukosen för periferal vävnad. Levern fortsätter<br />
att syntetisera glukos. Denna används för att fylla på det minskade glykogen lagren.<br />
Under tiden blodglukosnivån börjar öka slutför levern påfyllningen av sitt glykogen<br />
lager och börjar bearbeta det glukos som finns kvar för fettsyra syntes.<br />
Alltså, Levern tar inte upp glukos direkt, utan fortsätter med glukoneogenesen för att<br />
återfylla glykogen lagren. När blodsocker nivån igen är hög sätts nedbrytningen igång.<br />
3. <strong>Vad</strong> händer om det inte kommer mer mat<br />
The refed state visar vad som händer när glukos återigen blir tillgängligt för<br />
nedbrytning. Men vad händer om man inte äter igen Jo, Du kan dö såklart. Men innan<br />
dess….<br />
En 70 kg man har en energi reserv på ungefär 161 000 kcal. På en dag behöver denna<br />
man ca 1600 till 6000 kcal, beroende på grad fysisk aktivitet. Den lagrade energi som<br />
han redan har kan därför räcka upp till 1 till 3 månader. Kolhydrat lagren däremot är slut<br />
på bara en dag. Blodglukosnivån måste vara på minst 1,2 mM. Första prioriteten för<br />
<strong>metabolism</strong>en är att ge tillräckligt med glukos till hjärnan och andra vävnader som är<br />
beroende av denna energiform. Fettsyror kan inte omvandlas till glukos pga att acetyl<br />
CoA inte kan omvandlas till pyruvat. Glycerol kan omvandlas till glukos, men bara en<br />
viss bestämd mängd finns tillgänglig. Den andra hälften av glukos kommer från<br />
aminosyror från nedbrytningen av protein.<br />
Den andra prioriteten i svält <strong>metabolism</strong>en är att bevara proteinerna genom att ändra<br />
energiutvinningen från glukos till fettsyror och ketokroppar. De metaboliska<br />
förändringarna är som en övernatts fasta. Den låga blodsockernivån leder till en<br />
sänkning av insulin utsöndringen, men en ökning av glukagon utsöndringen. De större<br />
metaboliska processerna är mobilisering av triglycerol i fettvävnaden och<br />
glukoneogenesen i levern. Levern får energi genom att oxidera fettsyror från fettvävnad.<br />
Upptaget av glukos av muskler avtar pga den låga insulin nivån medan fettsyror upptas<br />
fritt. Detta betyder att muskeln ändrar energi utvinningen från glukos till fettsyror. B-<br />
oxidation av fettsyror sätter stopp för omvandlingen av pyruvat till acetyl CoA. Pyruvat,<br />
laktat och alanine används av levern för att producera glukos. Vid klyvning av<br />
triglyceroler bildas glycerol som sedan kan användas till syntes av glukos. Ett djurs
överlevnad kan vara beroende av att kunna röra sig. Detta betyder att djuret måste<br />
bevara sin muskel massa och minimera nedbrytningen av denna.<br />
Efter tre dagars svält har levern producerat stora mängder acetoacetat och D-3-<br />
hydroxybutyrat (även kallat ketokroppar).Så här går det till:<br />
Kolhydraternas uppgift är att se till att citronsyracykeln kan ta hand om alla<br />
acetylgrupper som bildas vid fettets nedbrytning. Acetylgrupperna kommer in i<br />
citronsyracykeln genom att reagera med oxalättiksyra till citrat. Cellerna behöver<br />
påfyllning av oxalättiksyra, det bildas av pyruvatjoner som i sin tur kommer från<br />
glykolysen. Om oxaloättiksyra saknas kommer acetygrupperna att reagera med varandra<br />
och bilda acetoacetat och 3-hydroxybutyrat – ketoföreningar. Ketoföreningarna bildas i<br />
levermitokondrierna, och släpps ut i blodet där de transporteras till kroppens vävnader.<br />
Vid denna tidpunkt börjar hjärnan att erhålla energi från acetoacetat istället för glukos.<br />
Efter tre dagars svält utvinner hjärnan en tredjedel av energin från ketokroppar. Hjärtat<br />
använder också ketokroppar som energi.<br />
Efter några veckors svält blir ketokropparna huvudkällan för energi utvinning. Nu<br />
används bara 40 gram glukos per dag av hjärnan istället för 120 gram den första dagen.<br />
Att använda ketokroppar istället för glukos sänker behovet av glukos för hjärnan. Detta<br />
leder till att mindre muskelmassa bryts ner nu än efter bara någon dag i svältperioden.<br />
20 gram muskelmassa bryts ner per dag i svält jämfört med 75 gram per dag i början av<br />
en svältperiod. Hur länge en människa kan överleva beror helt och hållet på hur mycket<br />
triglycerider som är lagrade.
Huvudenzymet Carnitin Acetyltransferas<br />
I svältstadiet omvandlar levern fettsyror till ketokroppar i mitokondriens matrix. Detta<br />
är beroende av om fettsyrorna kommer in i matrix eller ej. Fettsyror med långa<br />
kolkedjor kan endast komma in i matrix om de är esterfierade till carnitin. Det är<br />
enzymet carnitin acyltransferas 1 (också kallat carnitin palmitoyl transferase 1, CPT1)<br />
som katalyserar denna omvandling.<br />
Den här reaktionen kan inhiberas av malonyl CoA, (ett intermediat i fettsyrasyntesen).<br />
När malonyl CoA finns närvarnde kan inte fettsyrorna med de långa kolkedjorna komma<br />
in i matrix, där ß-oxidation och ketokroppsbildningen sker. Istället transporteras<br />
fettsyrorna till fett vävnaderna för att omvandlas till triglycerider. När bränsletillgången<br />
är låg är också malonyl CoA nivån låg och då frisätts fettsyrorna från fettvävnaderna<br />
och går vidare in till matrix.<br />
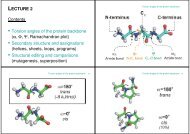
Bild 2. Humant peroxisomalt carnitine acetyltransferase (hpCAT).<br />
Vi fick i uppgift att beskriva ett enzym som relaterar till fettsyra <strong>metabolism</strong>en. Vi valde<br />
därför ett liknande enzym som tillhör samma familj som CPT1. Det enzym som vi nu<br />
ska presentera katalyserar samma reaktion, men detta enzymet är aktivt i peroxisomerna.<br />
Enzymet kallas humant peroxisomalt carnitine acetyltransferase (hpCAT). HpCAT är<br />
det namn som vidare kommer användas i denna presentation.
Bild 3. Humant peroxisomalt carnitine acetyltransferase (hpCAT), katalyserar Acetyl<br />
carnitine bildningen från Acetyl CoA och Carnitin.<br />
Detta är ett monomert protein som består av två lika stora _ / _ domän. Mellan dessa<br />
domän bildas en tunnel som korsar proteinet. Denna tunnel är åtkomlig för lösning och i<br />
mitten hittar man den aktiva ytan.<br />
I den katalytiska tunnel finns en His 322 , som föreslås delta i katalysen. En funktionellt<br />
konserverad Glu 326 påverkar His 322 och verkar vara nödvändig för katalys. Utifrån detta<br />
ges förslaget om en katalytisk diad. I nästan alla de andra medlemmarna från denna<br />
familj (Carnitin acyltransferaser) hittar man en aspartat i denna position, som<br />
funktionellt skulle kunna substitueras för glutamat. På enzym i dessa familjer har man<br />
bytt ut aspartatet mot alanin, vilket har inaktiverat enzymet totalt. Detta visar betydelsen<br />
av en negativ laddning i katalysen.<br />
I bindningsfickorna tillåts varje substrat att närma sig den aktiva ytan från var sin sida.<br />
Förslaget är att några sidokedjor bla Tyr 431 och Thr 444 vätebinder till syret på carnitin.<br />
Lys 398 ligger nära fosfat gruppen på CoA och Leu 142 binder CoA genom hydrofoba<br />
interaktioner med metylgrupper på ena sidan av substratet. Ser 407 och Asp 487 vätebinder<br />
till CoA.<br />
His 322 fungerar som en bas och deprotonerar primär alkoholen av carnitin. Detta<br />
resulterar i en positivt laddad histidin. Glu 326 polariserar histidinet för att öka den<br />
katalytiska aktiviteten, genom att stabilisera den positiva laddningen som utvecklas efter<br />
deprotoneringen av carnitin. Carnitinanjonen som bildas attackerar tioesterbindningen i<br />
acetyl CoA. Produkten blir sedan Acetylcarnitin och fritt Coenzyme A.
Bild 3. Sidokedjor som associeras med carnitin bindning och Co A bindning indikeras<br />
med mörka klot på banden. De ljusare kloten (gröna) visar sidokedjor som är essentiella<br />
för aktiviteten för acyltransferaser.<br />
Referenser<br />
Fettsyra<strong>metabolism</strong> :<br />
Biochemistry 5:th ed., Berg J, Tymoczko J, Stryer L, 2002 W.H Freeman & Co<br />
Acetyltransferaser:<br />
http://www.jbc.org/cgi/content/full/278/15/13159#F5