Teil VIII - Tired Joe
Teil VIII - Tired Joe
Teil VIII - Tired Joe

Sie wollen auch ein ePaper? Erhöhen Sie die Reichweite Ihrer Titel.
YUMPU macht aus Druck-PDFs automatisch weboptimierte ePaper, die Google liebt.
Grundstrukturen und Leistungen des Nervensystems<br />
Neurophysiologie<br />
1. Rezeptoren (Sinneszellen)<br />
Einteilung der Rezeptoren nach Art des Reizes:<br />
Optischer Reiz Photorezeptoren (Lichtsinneszellen im Auge)<br />
Mechanischer Reiz Mechanorezeptoren (Tastkörperchen der Hand)<br />
Thermischer Reiz Termorezeptoren (Wärme-, Kälterezeptoren der Haut)<br />
Chemischer Reiz Chemorezeptoren (Geruchs- Geschmackszellen)<br />
Elektrischer Reiz Elektrorezeptoren z.B. bei Haien<br />
Die Reizart kann auch adäquat oder inadäquat sein!<br />
Beispiel Photorezeptoren: Photorezeptoren reagieren auch auf Druck „Sternchen sehen“<br />
Farbwahrnehmung<br />
<br />
Einteilung der Rezeptoren nach der Lage im Organismus:<br />
Exterorezeptoren: nach außen zur Umwelt gerichtet, Beispiel: Photorezeptoren<br />
Enterorezeptoren: im Körperinneren, Infos über den Innenzustand z.B. CO 2 -Rezeptor im arteriellen Blut<br />
(Halsschlagader)<br />
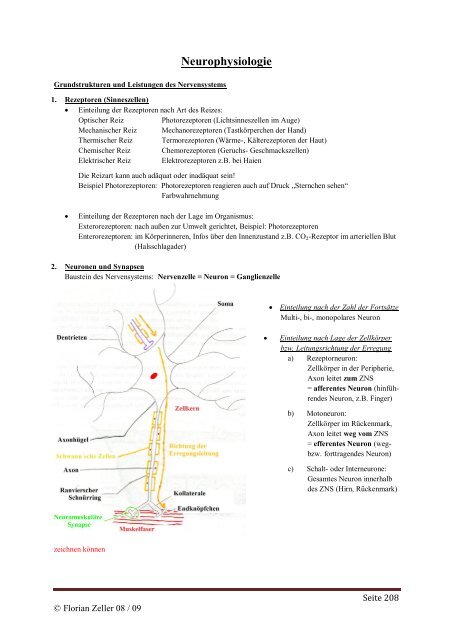
2. Neuronen und Synapsen<br />
Baustein des Nervensystems: Nervenzelle = Neuron = Ganglienzelle<br />
Einteilung nach der Zahl der Fortsätze<br />
Multi-, bi-, monopolares Neuron<br />
<br />
Einteilung nach Lage der Zellkörper<br />
bzw. Leitungsrichtung der Erregung<br />
a) Rezeptorneuron:<br />
Zellkörper in der Peripherie,<br />
Axon leitet zum ZNS<br />
= afferentes Neuron (hinführendes<br />
Neuron, z.B. Finger)<br />
b) Motoneuron:<br />
Zellkörper im Rückenmark,<br />
Axon leitet weg vom ZNS<br />
= efferentes Neuron (wegbzw.<br />
forttragendes Neuron)<br />
c) Schalt- oder Interneurone:<br />
Gesamtes Neuron innerhalb<br />
des ZNS (Hirn, Rückenmark)<br />
zeichnen können<br />
© Florian Zeller 08 / 09<br />
Seite 208
Bau eines Motoneurons:<br />
Die Axone sind vom Gliazellen umgeben, die das Axon umhüllen, stützen und schützen, es ernähren. Bei den<br />
markhaltigen = myelinisierten Nervenfasern der Wirbeltiere heißt die Gliazelle Schwann´sche Zelle. Sie ist<br />
ganz platt und mehrfach um das Axon gewickelt (siehe Abbildung), so dass viele Membranen aufeinander zu<br />
liegen kommen = Myelinscheide (Myelin ist das Material, z.B. auch weiße Substanz im ZNS im Gegensatz zu<br />
den „kleinen grauen“ Zellkörpern). Sie isoliert das Axon elektrisch, daher hohe Geschwindigkeit der Erregungsleitung<br />
möglich!!<br />
Bei unmyelinisierten = marklosen Nervenfasern ist die Gliazelle nur eine einfache Hülle für 1 oder mehrere<br />
Axone. Leitungsgeschwindigkeit gering. Vorkommen bei Wirbellosen.<br />
Formenvielfalt von Neuronen<br />
Nur zum anschauen!<br />
© Florian Zeller 08 / 09<br />
Seite 209
Synapsen<br />
Neuronale Versschaltung bzw. Möglichkeiten von Synapsen<br />
a) Zentrale Synapsen zwischen Nervenzellen<br />
- Axo - somatische Synapse , d.h. Synapse zwischen Axon und Soma<br />
- Axo – dentritische Synapse, d.h. Synapse zwischen Axon und Dentrit<br />
b) Synapse zwischen Axon und Muskel = neuromuskuläre Synapse = motorische Endplatte<br />
c) Synapse zwischen Axon und Drüsenzellen<br />
Neuromuskuläre Synapse = motorische Endplatte<br />
zeichnen können!!<br />
© Florian Zeller 08 / 09<br />
Seite 210
Die unerregte Nervenfaser - Ruhepotential<br />
Versuchsanordnung zur Messung des Ruhepotentials<br />
Extracelluläre Ableitung,<br />
Potentialdifferenz = 0<br />
intracelluläre Ableitung<br />
Potentialdifferenz - 50 mV bis<br />
- 90 mV bei Wirbeltieren<br />
= RUHEPOTENTIAL<br />
Bei Wirbeltiernervenzellen misst man eine Potentialdifferenz von ca. -70 mV, d.h. Zellinneres negativ im Vergleich<br />
zu Außen.<br />
Weitere Beobachtung:<br />
Die Potentialdifferenz tritt bereits beim Durchdringen der Membran auf, sie ist also an der Membran angelegt<br />
deswegen spricht man von Membranpotential.<br />
Die Potentialdifferenz ist an allen Punkten der Ruhenden Nervenzelle messbar!<br />
Darstellung im Diagramm:<br />
© Florian Zeller 08 / 09<br />
Seite 211
Entstehung (Aufbau) des Ruhepotentials<br />
Untersuchungen über die Art der Ionen innen und außen an der Axonmembran ergaben folgende Ergebnisse:<br />
1. Konzentrationsunterschiede (ungleiche Ionenverteilung)<br />
großer Buchstabe = viel vorhanden<br />
kleiner Buchstabe = wenig vorhanden<br />
sehr wichtig und zeichnen können!<br />
2. Selektive Permeabilität<br />
Die Nervenzellmembran ist für diese Ionen unterschiedlich durchlässig.<br />
Selektive Permeabilität für K + = 1<br />
Cl - = 0,1<br />
Na + = 0,01<br />
A - = 0<br />
Vorstellung: in der Membran sind kleine Poren, die die leinen K + -Ionen durchtreten lassen,<br />
auf die K + -Ionen wirken zwei Kräfte:<br />
a) Bestreben nach Konzentrationsausgleich: wegen guter Permeabilität diffundieren sie nach<br />
außen<br />
b) Die nun zunehmend unkompensierte negative Ladung (Org - !!) innen hält weitere K +<br />
durch elektrostatische Anziehung zurück, es kehren sogar manche K + nach innen zurück,<br />
so dass es nicht zu einem Konzentrationsausgleich der Ionen kommt, sondern zu einem<br />
GGW-Zustand der treibenden Kräfte! D.h. bei einer bestimmten Spannung stellt sich ein<br />
GGW ein zwischen Konzentrationsgefälle und Potentialgefälle, siehe Abbildung!<br />
Das Ruhepotential (RP) ist ein<br />
„Kaliumpotential“<br />
© Florian Zeller 08 / 09<br />
Seite 212
Zum Thema Leckströme<br />
Die Nervenzellmembran ist etwas durchlässig für Natriumkationen und Chloridionen.<br />
Folgen:<br />
- Geringer Chloridioneneinstrom stützt das Membranpotential.<br />
- Ständiges Einsickern von Natriumkationen ins Zellinnere (aufgrund des Konzentrationsgefälles und des<br />
Potentialgefälles) müsste eigentlich das Membranpotential schmälern. Kaliumionen können leichter nach<br />
außen treten, das Ruhepotential müsste irgendwann zusammenbrechen.<br />
Dies ist jedoch am intakten Axon nicht der Fall !!<br />
Ursache: gegenläufig arbeitende Natrium-Kalium-Ionenpumpe,<br />
ein aktiver Ionentransportmechanismus. Die Pumpe ist eine Membranstruktur , die unter Energieverbrauch<br />
(ATP-Verbrauch) Natriumionen aus dem zellinneren gegen Kaliumionen aus dem Außenmedium<br />
austauscht. Der Transport erfolgt gegen den jeweiligen Konzentrationsgradienten, also „bergauf“.<br />
Die erregte Nervenfaser – Auslösung und Entstehung von Aktionspotentialen<br />
Prinzip:<br />
Überschwelliger Reiz<br />
(gereizte Sinneszelle, Stromstoß)<br />
Erregung der Nervenzelle<br />
Folge: Membran wird kurzfristig umgepolt, die Umpolungswelle<br />
Pflanzt sich über das Axon fort. Diese Potentialänderung<br />
bezeichnet man als<br />
AKTIONSPOTENTIAL<br />
© Florian Zeller 08 / 09<br />
Seite 213
Um Näheres über die Auslösebedingungen und die charakteristischen Eigenschaften eines Aktionspotentials zu<br />
erfahren, verwendet man die Versuchsanordnung zur gleichzeitigen intrazellulären Reizung und Ableitung von<br />
einem Axon.<br />
Auslösung von Aktionspotentialen<br />
1. Bei nicht aktivem Reizgerät erfasst das Messgerät das Ruhepotential von – 70 mV.<br />
2. Ist die Reizelektrode mit dem negativen Pol des Reizgerätes verbunden, so wird dem Axoninneren eine<br />
gewisse Menge negativer Ladung zugeführt. Diese bewirkt eine entsprechende Verschiebung des<br />
Membranpotentials in den Bereich stärker negativer Werte. Die „Antwort“ der Membran ist passiv.<br />
Diese Verstärkung des Ruhepotentials wird als Hyperpolarisation bezeichnet.<br />
3. Ebenfalls passive Antwort; die Größe der Hyperpolarisation ist der Reizspannung proportional.<br />
4. Ist die Reizelektrode mit dem positiven Pol des Reizgerätes verbunden, so wird dem Axoninneren eine<br />
gewisse menge positiver Ladung zugeführt. Diese bewirkt eine entsprechende Verschiebung des<br />
Membranpotentials in den Bereich weniger stark negativer Werte. Die „Antwort“ der Membran ist wie<br />
bei 2. und 3. passiv. Diese Abschwächung des Ruhepotentials wird als Depolarisation bezeichnet.<br />
5. & 6. Ebenfalls passive Antwort; die Größe der Depolarisation ist der Reizspannung proportional.<br />
© Florian Zeller 08 / 09<br />
Seite 214
7. – 9. Ab einer bestimmten Reizschwelle kommt es zur Auslösung von Aktionspotentialen. Die „Antwort“<br />
der Membran ist aktiv. Dabei gilt die „Alles-oder-Nichts-Regel“. Sie gesagt, dass sämtliche überschwelligen<br />
Reize die Bildung eines Aktionspotentials bewirken, sämtliche Reize unterhalb der<br />
Schwelle hingegen nicht. Das Aktionspotential läuft in immer gleicher, starrer Weise ab.<br />
10. Am Ende jedes Aktionspotentials kann es zu sog. Nachpotentialen kommen.<br />
Also: Nur ein überschwelliger Reiz bewirkt eine überproportionale Depolarisation (aktive „Antwort“) der<br />
Axonmembran, und damit eine Potentialumkehr:<br />
Das Aktionspotential schnellt auf + 30 mV hoch.<br />
Die Höhe (+30 mV) des Aktionspotentials ist unabhängig von der Größe des überschwelligen Reizes<br />
Alles-oder-Nichts-Signal !!!<br />
Nur ein Aktionspotential wird über das Axon weitergeleitet !!<br />
Die Phasen eines Aktionspotentials<br />
Depolarisation: Sehr rasche positive Potentialänderung in 0,2 – 0,5 ms, negative Ruheladung geht verloren;<br />
positiver Anteil des Aktionspotential = Überschuss.<br />
Repolarisation: Normale Polarisation der Zellmembran wird wieder hergestellt, Prozess verläuft gegen Ende<br />
langsamer.<br />
Nachpotentiale: Bei der Rückkehr des Potentials von der Aktionspotentialspitze, schwingt es über den normalen<br />
Wert des Ruhepotential hinaus, erst nach hyper- & depolarisierten Nachpotential wird der<br />
Ruhewert erreicht.<br />
Ein Aktionspotential ist ein für jede Nervenzelle konstanter Ablauf von Depolarisation und Repolarisation,<br />
der immer dann selbsttätig auftritt, wenn die Membran über den Schwellenwert hinaus depolarisiert<br />
wird.<br />
Nachpotentiale: Bei der Rückkehr des Aktionspotentials von der Spitze stellen sich noch „Nachschwingungen“<br />
ein, danach wird der Wert des Ruhepotentials wieder erreicht!<br />
Hinweis: Wenn Sie die Phasen eines Aktionspotentials skizzieren, achten Sie darauf, dass Sie die Depolarisation<br />
steiler zeichnen als die Repolarisation!!<br />
© Florian Zeller 08 / 09<br />
Seite 215
Refraktärzeiten<br />
Kommen zwei Reize zeitlich knapp hintereinander, so beobachtet man Refraktärzeiten.<br />
Absolute Refraktärzeit: Bei sehr enger zeitlicher Aufeinanderfolge (2 ms und weniger) löst der eigentlich<br />
überschwellige zweite Reiz kein Aktionspotential aus.<br />
Relative Refraktärzeit:<br />
In der Zeitspanne zwischen 5 und 2 ms nach der Auslösung eines Aktionspotentials<br />
Kann nur ein abgeschwächtes Aktionspotential ausgelöst werden.<br />
Die ursprüngliche Ionenverteilung wird durch die Na-K-Pumpe wiederhergestellt.<br />
Hinweis: Die Zahl der Ionen, die während eines Aktionspotentials die Membran passieren ist im Vergleich zur<br />
Gesamtzahl relativ gering. Deshalb können auch bei blockierter Na-K-Pumpe noch ca. 1000 Aktionspotentiale<br />
ausgelöst werden. Die Na-K-Pumpe hat also keinen Einfluss auf das Aktionspotential.<br />
Alles-oder-Nichts-Regel:<br />
Alle überschwelligen Reize lösen ein Aktionspotential aus, das für eine bestimmte<br />
Nervenzelle immer die gleiche Größe besitzt.<br />
Verschlüsselung der Reizstärke:<br />
Starke Reize lösen pro Zeiteinheit eine größere Zahl von Aktionspotentialen<br />
aus als schwache Reize.<br />
„Die Reizstärke ist durch die Frequenz der Aktionspotentiale codiert“.<br />
(max. Frequenz ca. 500 Aktionspotentiale pro Sekunde, begrenzt durch Refraktärzeit)<br />
© Florian Zeller 08 / 09<br />
Seite 216
Die Rolle der Ionen beim Aktionspotential<br />
Erinnerung: Das Ruhepotential ist ein „Kaliumpotential“<br />
Das Aktionspotential ist ein „Natriumpotential“!<br />
Sie müssen jetzt wissen, dass die Nervenzellmembran neben den stets offenen Kaliumionenporen auch spannungsgesteuerte<br />
Ionenporen (Ionenkanäle) für Natriumkationen enthält !!!<br />
Und auch spannungsgesteuerte K + -Poren !<br />
Was passiert bei der Depolarisationsphase?<br />
Ein überschwelliger Reiz depolarisiert die Membran über den Schwellenwert hinaus,<br />
Spannungsgesteuerte Natriumionenporen öffnen sich schlagartig,<br />
Natriumkationen strömen schlagartig in das Axon ein<br />
(wegen Konzentrationsgefälle und Potentialgefälle (innen - )),<br />
führt zur Potentialumkehr an der Membran,<br />
die Membraninnenseite wird positiv geladen.<br />
Bei +30 mV schließen sich die Natriumionenkanäle.<br />
(Ein Ausgleich der Natriumionenkonzentration auf beiden Seiten der Membran wird dabei nicht erreicht.)<br />
Jetzt setzt die Repolarisation ein: Spannungsgesteuerte K + -Poren<br />
Kaliumionen diffundieren verstärkt aus der Zelle durch die Membran,<br />
(wegen Konzentrationsgefälle und negativer Ladung außen),<br />
bis das Ruhepotential (nach den Schwankungen der Nachpotentiale) wieder hergestellt ist.<br />
Tatsächlich ist experimentell eine unterschiedliche Membranleitfähigkeit für die Ionen während des Aktionspotentials<br />
messbar.<br />
Die Dauer eines Aktionspotentials (Depolarisation, Umpolung, Repolarisation, Nachpotential) beträgt 4 – 5 ms.<br />
Während der Depolarisation und der Repolarisation (ca. 2 ms) kann auch durch einen überschwelligen Reiz kein<br />
weiteres Aktionspotential ausgelöst werden (absolute Refraktärphase). Der Natriumionenporen-Öffnungsmechanismus<br />
ist ganz einfach nicht funktionsfähig.<br />
Während der relativen Refraktärzeit (von ca. 2 – 5 ms) ist sowohl die Natriumionen- als auch die Kaliumionenpermeabilität<br />
gebremst.<br />
© Florian Zeller 08 / 09<br />
Seite 217
Zum Anschauen !<br />
Ionenverteilung nach einem Aktionspotential<br />
mehr Na + innen<br />
mehr K + außen<br />
als ursprünglich<br />
Doppelte Aufgabe der K + /Na + - Pumpe<br />
Sie transportiert die während eines Aktionspotentials<br />
eingeströmten Na + wieder<br />
nach außen und die K + nach innen.<br />
Mit dem PORENMODELL lassen sich weitere Sachverhalte für die Auslösung von Aktionspotentialen erklärt.<br />
Alles-oder-Nichts-Regel<br />
unterschwelliger Reiz:<br />
überschwelliger Reiz:<br />
Zahl der geöffneten Na + - Poren reicht nicht aus, um Entladung der Membran zu<br />
bewirken.<br />
schlagartige Öffnung aller Na + - Poren konstante Amplitudenhöhe von Aktionspotential.<br />
Eine noch höhere Reizstärke kann keine Amplitudenerhöhung bewirken, da<br />
die Zahl der Na + - Poren begrenzt ist.<br />
Refraktärzeiten<br />
absolute Refraktärzeit: Nach einem ausgelösten Aktionspotential ist der Na + - Porenöffnungsmechanismus<br />
1 – 2 ms nicht funktionsfähig.<br />
relative Refraktärzeit:<br />
Weitere 3 ms ist die Erregbarkeit der Nervenfaser noch herabgesetzt. Nur bei erhöhter<br />
Reizschwelle öffnen aller Poren.<br />
© Florian Zeller 08 / 09<br />
Seite 218
Latenzzeit ≠ Reflektärzeit<br />
Weiterleitung von Aktionspotentialen<br />
Erregungsleitung in einer marklosen Nervenfaser<br />
skizzieren<br />
können<br />
<br />
<br />
<br />
<br />
<br />
(1) An der Stelle, an der gerade ein Aktionspotential auftritt, wird das Membranpotential durch einen starken<br />
Einstrom von Na + - Ionen umgepolt.<br />
(2) An beiden Seiten der Membran stoßen damit positive und negative Ladungen direkt aufeinander.<br />
(3) Da sich entgegengesetzte Ladungen anziehen, kommt es zu einer Verschiebung der Ionen in Längsrichtung<br />
des Axons. Diese Ionenströme („Kreisströmchen“) bewirken eine Veränderung des Membranpotentials<br />
in der unmittelbaren Nachbarschaft der gerade umgepolten Stelle.<br />
(4) Die Nachbarstelle wird auf diese Weise überschwellig depolarisiert, und es bildet sich an ihr ein Aktionspotential<br />
aus.<br />
(5) An der ursprünglich umgepolten Stelle stellt sich wieder das Ruhepotential ein.<br />
© Florian Zeller 08 / 09<br />
Seite 219
(6 & 7) Ionenströme bewirken wiederum die überschwellige Depolarisation der Membran in der unmittelbaren<br />
Nachbarschaft der aktuell umgepolten Stelle.<br />
(8) An der Nachbarstelle entsteht erneut ein Aktionspotential, das in der beschriebenen Form auf die neue<br />
Nachbarstelle einwirkt.<br />
Das Aktionspotential breitet sich unter Beibehaltung seiner vollen Stärke entlang der gesamten Nervenfaser<br />
aus (kontinuierliche Erregungsleitung).<br />
Die Leitungsgeschwindigkeit (ca. 10 m s ) ist abhängig von der Dicke des Axons.<br />
Aufgabe:<br />
In welcher Richtung kann die Erregung<br />
a) bei künstlicher Reizung eines Axons laufen?(vom Soma zum Endknöpfchen)<br />
b) im Körper nehmen? (in beide)<br />
Erregungsleitung in einer markhaltigen Nervenfaser<br />
Bei der markhaltigen Nervenfaser sind weite Bereiche des Axons durch eine Markscheide isoliert. Nur an einem<br />
RANVIER-Schnürring können sich Ionen durch die Membran bewegen.<br />
<br />
<br />
<br />
<br />
<br />
<br />
(1) An der Einschnürung, an der gerade ein Aktionspotential auftritt, wird das Membranpotential durch den<br />
Einstrom von Na + - Ionen umgepolt.<br />
(2) Die Ionenströme (Kreisströmchen) können wegen der isolierend wirkenden Markscheide erst am<br />
nächsten Schnürring eine Depolarisation bewirken.<br />
(3) Dort wird ein Aktionspotential gebildet, während sich an der ursprünglich umgepolten Stelle wieder das<br />
Ruhepotential einstellt.<br />
(4) Die Ionenströme führen dann zur Weitergabe des Aktionspotentials an den nächstfolgenden Schnürring.<br />
Die Erregung springt gleichermaßen von Schnürring zu Schnürring. Man spricht deshalb auch von saltatorischer<br />
Erregungsleitung.<br />
Die Leitungsgeschwindigkeit beträgt ca. 100 m s.<br />
Aufgabe:<br />
Geben Sie drei Gründe an, weshalb sich markhaltige Nervenfasern gegenüber marklosen Nervenfasern im Verlauf<br />
der Evolution durchgesetzt haben?<br />
- Materialersparnis bei gleicher Leistung (Platzersparnis)<br />
- ATP-Verbrauch geringer weniger Na + / K + - Pumpen werden benötigt (Energieersparnis)<br />
- Schneller (Zeitersparnis)<br />
© Florian Zeller 08 / 09<br />
Seite 220
Vorgänge an der neuromuskulären Synapse, chemische Erregungsübertragung<br />
durch Transmitter<br />
1. Ein Aktionspotential kommt an der präsynaptischen Membran an, löst über Calciumioneneinstrom die Ausschüttung<br />
des Transmitterstoffs Acetylcholin, das in den synaptischen Bläschen gespeichert ist, aus.<br />
2. Die Acetylcholinmoleküle diffundieren über den synaptischen Spalt (Dauer: max. 1 ms).<br />
3. Die Acetylcholinmoleküle setzen sich an der subsynaptischen Membran weist wie die Membran der Nervenzelle<br />
ein Ruhepotential auf.<br />
4. Durch eine Konformationsänderung der Rezeptormoleküle kommt es zu einer Permeabilitätsänderung der<br />
Membran für Natrium- und Kaliumkationen.<br />
5. Als Folge wird die subsynaptische Membran depolarisiert, es entsteht ein sog. „Endplattenpotential“, das<br />
sich in den postsynaptischen Membranbereich der Muskelfasermembran hinein ausbreitet und letzendlich<br />
die Kontraktion einer Muskelfaser auslöst.<br />
6. Ein an der subsynaptischen Membran befindliches Enzym – die Cholinesterase – löst die Acetylcholinmoleküle<br />
augenblicklich wieder ab, indem es sie in Cholin und Essigsäure spaltet.<br />
7. Diese Moleküle diffundieren zur präsynaptischen Membran zurück, werden ins Endknöpfchen aufgenommen,<br />
wieder zu Acetylcholin verbunden und in die synaptischen Bläschen verpackt!<br />
© Florian Zeller 08 / 09<br />
Seite 221
Störung der Synapsenfunktion durch Gifte<br />
© Florian Zeller 08 / 09<br />
Seite 222
Beeinflussung der synaptischen Erregungsübertragung<br />
1. Hemmung der Erregungsübertragung durch Giftstoffe<br />
Hemmstoff Angriffsort Wirkungsart<br />
Botulinus-Toxin<br />
Curare und<br />
Derivate<br />
E 606<br />
Sarin<br />
präsynaptische<br />
Membran<br />
subsynaptische<br />
Membran<br />
Acetylcholinesterase<br />
Hemmt die Acetylcholinfreisetzung<br />
Blockade der Erregung Lähmung<br />
Besetzt Ach-Rezeptoren, ohne eine Erregung auszulösen,<br />
kann nicht von Enzymen gespalten werden Lähmung<br />
Hemmung der Acetylcholinesterase, erst Erregung<br />
Krämpfe, dann Lähmung (Kompetitiver Hemmstoff)<br />
2. Wirkung von Psychopharmaka und Drogen<br />
Substanzen, die die Aktivität des Nervensystems, insbesondere des Gehirns beeinflussen. Sie wirken<br />
dadurch auf psychische Prozesse ein.<br />
(nur zum anschauen)<br />
© Florian Zeller 08 / 09<br />
Seite 223
Das Gehirn<br />
(Wirbeltiere)<br />
Bildung der Neurula mit Verschluss des Neuralrohres.<br />
V = Vorder-<br />
M = Mittel-<br />
H = Hinterhirnbläschen<br />
Die Entwicklung des menschlichen Gehrins<br />
© Florian Zeller 08 / 09<br />
Seite 224
© Florian Zeller 08 / 09<br />
Seite 225
Längsschnitt durch das Gehirn des Menschen<br />
1 = Großhirn = Vorderhirn<br />
2 = Balken<br />
3 = Zwischenhirn<br />
4 = Mittelhirn<br />
5 = Kleinhirn = Hinterhirn<br />
6 = Hypophyse (Hirnanhangsdrüse)<br />
7 = Nachhirn = verlängertes Mark<br />
Zwischen-, Mittel- & Nachhirn wird als<br />
Stammhirn bezeichnet.<br />
Das Gehirn des Menschen<br />
„Die Größe des Gehirns sagt nichts über die Intelligenz aus!“<br />
‣ Das Gehirn ist <strong>Teil</strong> des Zentralnervensystems, der bei Wirbeltieren<br />
im Schädel liegt. Beim Menschen ist es etwa eine 1,3 kg schwere<br />
Masse aus rosa-grauem Gewebe, mit ca. 10 Milliarden Nervenzellen,<br />
die untereinander verknüpft sind.<br />
1. Großhirn (= Vorderhirn)<br />
Es besteht aus 2 Hälften, der linken und der rechten Großhirnhemisphäre, und macht etwa 85 % der gesamten<br />
Hirnmasse aus. Hier liegt der Sitz der Gedächtnisleistung (Motivation, Lernen/Speicher) und Intelligenzleistung,<br />
sowie des Bewusstseins und des Willens.<br />
2. Balken (Corpus Callosum)<br />
Ein Faserbündel, das die beiden Großhirnhemisphären miteinander verbindet und Informationsaustausch<br />
untereinander ermöglicht.<br />
3. Zwischenhirn<br />
Es umfasst den Thalamus, Hypothalamus und die Hypophyse.<br />
Thalamus: spielt bei allen Sinnesreizen außer dem Geruchssinn eine Rolle. Vielerlei für die Großhirnrinde<br />
bestimmte Signale durchlaufen diese Struktur. Verbindung zwischen Sinnesorganen und Großhirn<br />
(Filter!)<br />
Hypothalamus: steuert die Anpassung von Funktionen wie Herzfrequenz und Körpertemperatur. Darüber<br />
hinaus bestimmt er Schlafrhythmus und Hormonkonzentration. Er ist das Steuerzentrum für das vegetative<br />
Nervensystem.<br />
4. Mittelhirn<br />
Leitet sensorische Impulse aus dem Rückenmark in andere Hirnabschnitte und steuert Reflexantworten<br />
auf bestimmte Reize. Steuerung von Aktivität und Ruhe.<br />
© Florian Zeller 08 / 09<br />
Seite 226
5. Kleinhirn (= Hinterhirn)<br />
Ist für Gleichgewicht und Haltung verantwortlich, es verarbeitet Informationen über das Raumgefühl<br />
und koordiniert alle Bewegungen des Menschen.<br />
6. Hypophyse (Hirnanhangsdrüse)<br />
Setzt unter der Kontrolle des Hypothalamus Hormone frei, die im Körper zirkulieren und wiederum die<br />
Tätigkeit anderer Drüsen steuern.<br />
7. Nachhirn (verlängertes Mark)<br />
Unterstützt die Regulation von Atmung, Schlucken, Blutdruck und Reaktionen wie Niesen ebenso wie<br />
den Schlaf. Sitz vieler Reflexzellen.<br />
8. Rückenmark<br />
Übermittelt Impulse aus dem Gehirn an andere Körperabschnitte und leitet von dort aus Nachrichten<br />
zurück an verschiedene Hirnstrukturen.<br />
„ Das menschliche Gehirn ist eine großartige Sache. Es funktioniert vom Moment der Geburt an – bis zu dem<br />
Zeitpunkt, wo du aufstehst, um eine Rede zu halten.“<br />
(Mark Twain)<br />
© Florian Zeller 08 / 09<br />
Seite 227
Rückenmark: Querschnitt (im Bereich zwischen zwei Wirbeln)<br />
Rückenmarkwurzeln:<br />
Hinterwurzel: alle afferenten Nervenfasern treten über sie ein; Zellkörper der afferenten Nervenfasern<br />
befinden sich im Hinterwurzel- oder Spinalganglion.<br />
Vorderwurzel: alle efferenten Nervenfasern treten über sie aus! Zellkörper der efferenten Nervenfasern<br />
liegen im vorderem <strong>Teil</strong> der grauen Substanz des Rückenmarks.<br />
Spinalnerv: beide Wurzeln vereinigen sich zum „gemischten“ Nerv, teilt sich aber bald wieder auf<br />
und versorgt somatisches und vegetatives Nervensystem.<br />
Das Rückenmark<br />
© Florian Zeller 08 / 09<br />
Seite 228
Das Nervensystem der Wirbeltiere und des Menschen<br />
Zentrales<br />
Nervensystem<br />
Gehirn<br />
Rückenmark<br />
Nervensystem<br />
Peripheres<br />
Nervensystem<br />
somatisches<br />
Nervensystem<br />
vegetatives<br />
Nervensystem<br />
Motorische Nervenbahnen<br />
Afferenzen<br />
Sensorische Nervenbahnen<br />
Efferenzen<br />
Versorgt die inneren Organe<br />
animales Nervensystem<br />
vegetatives Nervensystem<br />
Autonomes = vegetatives Nervensystem<br />
Das vegetative Nervensystem innerviert die glatte Muskulatur der inneren Organe und zwar antargonistisch. Es<br />
regelt unter anderem die Funktion von:<br />
- Atmung<br />
- Kreislauf<br />
- Verdauung<br />
- Stoffwechsel<br />
- Drüsen<br />
Die zentralen Bereiche liegen im Gehirn, Rückenmark und dem sog. Grenzstrang. Sie sind über den peripheren<br />
Bereich mit den inneren Organen verbunden.<br />
2 Systeme<br />
Sympathikus<br />
Parasympathikus<br />
- Mobilisiert den Körper - fördert Erholung + Verdauung<br />
(Angriff, Kampf, Flucht)<br />
- Ausgleich des inneren Gleichgewichts<br />
- tagaktiv - nachtaktiv<br />
- „aggressiv“ - „friedlich“<br />
Transmitter:<br />
- Adrenalin Acetylcholin<br />
- Noradrenalin<br />
© Florian Zeller 08 / 09<br />
Seite 229
Wenn wir schnell viele Treppen hochsteigen, kommen wir leicht „außer Atem“. Unser Herz klopft und wir atmen<br />
sehr hastig. Nach wenigen Minuten haben sich Herz- und Lungentätigkeit wieder normalisiert. Unser Wille<br />
war an diesen Vorgängen nicht beteiligt. Die Steuerung erfolgte über selbstständig arbeitende Nervenzentren und<br />
Nervenbahnen, die man unter der Bezeichnung „vegetatives Nervensystem“ zusammenfasst.<br />
Oberstes Steuerungszentrum des vegetativen<br />
Nervensystems ist ein <strong>Teil</strong> des Zwischenhirns,<br />
der Hypothalamus. Von hier aus ziehen zwei<br />
Nervenstränge links und rechts der Wirbelsäule<br />
entlang, die man als Grenzstränge bezeichnet.<br />
Neben jedem Wirbelkörper verdicken sie sich<br />
zu Nervenknoten, von denen Nervenfasern zu<br />
den inneren Organen ziehen. Im Bauchraum<br />
und im Unterbauch werden ganze Geflechte von<br />
Nervenknoten gebildet. Bekannt ist der „Solarplexus“<br />
(„Sonnengeflecht“) im Bauchraum, bei<br />
Boxern ein Ziel für einen möglichen K.O.-<br />
Schlag.<br />
Man nennt den bisher beschriebenen <strong>Teil</strong> des<br />
vegetativen Nervensystems den Sympathikus.<br />
Ein zweiter <strong>Teil</strong> des vegetativen Nervensystems<br />
besteht aus mehreren Einzelnerven, die im Hypothalamus<br />
und anderen <strong>Teil</strong>en des Stammhirns<br />
entspringen und zu den inneren Organen<br />
führen. Der bekannteste Nerv in diesem System<br />
ist der Vagus. Man nennt das System den Parasympathikus.<br />
Wieso besteht das vegetative Nervensystem<br />
aus zwei <strong>Teil</strong>en?<br />
Jedes innere Organ ist sowohl mit dem Sympathikus<br />
als auch mit dem Parasympathikus<br />
verbunden. Der Sympathikus treibt z. B. den<br />
Herzschlag an, der Parasympathikus verlangsamt ihn. Andererseits regt der Vagus (<strong>Teil</strong> des Parasympathikus)<br />
die Magendrüsen zur Sekretion an, und der Sympathikus bremst diesen Prozess. Die beiden Systeme sind also<br />
Gegenspieler und regeln so die Tätigkeit der inneren Organe unabhängig vom Großhirn. Der Sympathikus fördert<br />
die Aktivierung des Körpers; er bringt den Körper „in Schwung“. Der Parasympathikus bremst dagegen<br />
viele Organe, er fördert die Erholung und damit diejenigen Organe, die in der Erholungsphase aktiv sind, wie<br />
z. B. der Verdauungstrakt.<br />
Wieso läuft es uns bei Angst „kalt den Rücken hinunter“?<br />
Jeder kennt diese Redensart, doch kaum einer weiß, dass sie ihre Ursache in der Sympathikus-Tätigkeit hat. Das<br />
vegetative Nervensystem ist nicht völlig unabhängig vom Großhirn. Wenn wir plötzlich erschrecken, wird über<br />
das Großhirn der Sympathikus aktiviert. Die Impulse breiten sich relativ langsam (1 bis 2 m s) über das Rückenmark<br />
nach unten aus und bewirken von oben nach unten absteigend eine Kontraktion der peripheren Blutgefäße.<br />
Dies wird von Nervenendigungen in der Haut registriert.<br />
© Florian Zeller 08 / 09<br />
Seite 230
+ = aktiv - = nicht aktiv<br />
Beispiele:<br />
Alle Emotionen z.B. Angst, Aggression, Freude usw. finden immer mit Beteiligung des vegetativen Nervensystems<br />
statt.<br />
Jäger in der Steinzeit liegt am Lagerfeuer, als er plötzlich ein Knacken hört:<br />
Knacken löst Erbrecken und Stress aus; es kommt zur Denkblockade; die Wahrnehmung „Angst“ wird an das<br />
Zwischen (Hypothalamus) gesendet; der Sympathikus wird aktiviert – Nebenniere schüttet Adrenalin und Noradrenalin<br />
aus; dies bewirkt eine Beschleunigung des Herzschlags, der Blutdruck steigt, der Kreislauf verändert<br />
sich; Jäger spring auf!<br />
Zucker und Fettreserven werden angezapft, um die Muskelversorgung zu decken; Jäger rennt davon oder ist nun<br />
auch bereit zu kämpfen; (= akute Phase). Danach folgt Ruhezustand = sog. Erholungsphase.<br />
Ein Mensch aus der heutigen Zeit befindet sich in einem Dauerstresszustand;<br />
Der permanente Stress ist nicht abbaubar, dies bedeutet einen nicht lösbaren Widerstand; auf die akute Phase<br />
kann keine Erholungsphase folgen, daher kommt es zur Erschöpfung (Magengeschwüre, Herzinfarkt)<br />
Lösungsmöglichkeit: z.B. durch Entspannungstraining = eine willentliche Einflussnahme auf das vegetative<br />
Nervensystem. Besonders Atemübungen haben eine gute Wirkung auf den Parasympathikus („Abschalten“ üben,<br />
Verspannungen lösen)<br />
Jedoch erfolgt dadurch keine Ursachenänderung, sondern nur eine Anpassung an den Stress. Ebenso wichtig eine<br />
Veränderung der Ursachen, soweit als möglich.<br />
© Florian Zeller 08 / 09<br />
Seite 231
Reflexbogen<br />
= Bezeichnung für die Nervenschaltung eines Reflexes<br />
Eine über Nervenzellen laufende Verbindung zwischen Sinnesorganen und Muskeln bezeichnet man als Reflexbogen,<br />
die durch den Reiz ausgelöste Reaktion als Reflex.<br />
Reflexe<br />
Reflexe sind einfache Reaktionen, die dann in immer gleicher, stereotyper und rascher Weise auf spezifische<br />
Reize hin eintreten, wenn der Reiz eine bestimmte Stärke aufweist. Reflexe sind stets funktionsbereit. Es scheint<br />
für Reflexe ein genetisch fixiertes Programm zu geben, daher bezeichnet man sie als angeboren.<br />
Zu den Schutzreflexen zählt man z.B. den Kratz-, Rückzieh-, Liedschluss-, Nieß-, Husten-, Brechreflex und<br />
Kniesehnenreflex.<br />
Im Zusammenhang mit der Ernährung wirkt der Schluck- und Speichelflussreflex. Lebenserhaltender Reflex ist<br />
z.B. der Atemreflex.<br />
Reflexe kann man unterteilen in Fremdreflexe und Eigenreflexe<br />
Fremdreflex bedeutet, dass der Weg von einem Rezeptororgan über normalerweise mehrere afferente und efferente<br />
Nervenbahnen zum Effektororgan führt. Rezeptor und Effektor liegen in verschiedenen Organen, meist<br />
polysynaptisch, z.B. Rückziehreflex bei heißer Herdplatte<br />
Eigenreflex bedeutet, dass von der Muskelspindel als Rezeptor der Weg zum gleichen Muskel als Effektor zurükführt.<br />
(Rezeptor und Effektor im selben Organ, meist monosynaptisch!)<br />
Ein beliebtes Beispiel ist der Kniesehnenreflex<br />
Dieser monosynaptische Reflex wird an einem „Dehnungsfühler“, den Muskelspindel der Wade wahrgenommen.<br />
Er wird über afferente Nervenbahnen durch elektrische Impulse (Rezeptorneuron) zum zentralen Nervensystem<br />
übertragen und dort verarbeitet. Auf efferenten Nervenbahnen gelangt die Antwort auf diesen Reiz zu<br />
einem Effektor (Muskel, Drüse), der dadurch aktiviert wird und eine Reaktion bedingt.<br />
Den gesamten Weg bezeichnet man als Reflexbogen<br />
Reflexe von Neugeborenen sind z.B. Lippen-, Saug-, Hand- bzw. Fußgreifreflex; Augenschluss-, Stütz- und<br />
Gehreflex.<br />
Einige dieser Neugeborenen Reflexe verlieren sich in den ersten Wochen bis Monaten.<br />
© Florian Zeller 08 / 09<br />
Seite 232
= Interneuron<br />
Verschaltungen in Reflexbögen<br />
© Florian Zeller 08 / 09<br />
Seite 233
Sarko = Fleisch<br />
(Sarkorak = Fleischfresser)<br />
© Florian Zeller 08 / 09<br />
Seite 234
© Florian Zeller 08 / 09<br />
Seite 235
Bau der Quergestreiften Muskulatur<br />
Eine Muskelfaser ist aus vielen Myofibrillen zusammengesetzt.<br />
Bau einer Myofibrille: Die Myofibrille wird durch sog. Z-Scheiben in ca. 2,5 μm lange Abschnitte gegliedert,<br />
die Sarkomere. Die Sarkomere sind die kontraktilen Einheiten des Muskels. Sie bestehen<br />
aus Eiweißfilamenten:<br />
dicke Eiweißfilamente = Myosin<br />
dünne Eiweißfilamente = Actin<br />
Die besondere Anordnung der Myosin- und Actinfilamente ergibt eine Streifung<br />
quergestreifte Muskulatur.<br />
Aktinfilamente<br />
Myoisnfilamente<br />
(zeichnen können!)<br />
Mechanismus der Muskelkontraktion<br />
Eine Muskelzelle besitzt wie jede Zelle Organellen, besonders auffällig sind jedoch<br />
- viele Mitochondrien (hoher Stoffwechselumsatz)<br />
- zwei Hohlraumsysteme, die nicht ineinander übergehen und verschiedene Aufgaben erfüllen:<br />
a) Endoplasmatisches Reticulum, hier Sarkoplasmatisches Reticulum genannt, mit Ca 2+ -Speicher<br />
b) T-System = Transversales Tubuli-System, die tubuli verlaufen quer auf Höhe der Z-Scheiben,<br />
leiten Depolarisation ins Innere<br />
Feinstruktur des Sarkomers:<br />
Aktinfilamente: Aktin: zwei Keten von Nonomeren<br />
Tropomyosin: stäbchenförmiges Molekül<br />
Troponin: am Ende des Tropomyosins<br />
Myosinfilamente: langgestreckte Moleküle mit Köpfchen, die seitlich herausragen<br />
Molekularer Mechanismus der Kontraktion<br />
Die Länge der Actin- und Myosinfilamente ändert sich dabei nicht!!!<br />
© Florian Zeller 08 / 09<br />
Seite 236
Ruhender Muskel:<br />
Die Interaktion zwischen Myosinköpfchen und Aktin ist durch das Troponin behinder.<br />
Aktiver Muskel:<br />
Über die Muskelfasermembran trifft eine Erregung ein, das T-System leitet die Depolarisation ins Innere<br />
Ca 2+ -Permeabilität der Sarkomer-Membran wird erhöht, Ca 2+ diffundiert aus dem Sarkomer zu<br />
den Filamenten und startet den Bewegungsmechanismus = Gleitfilamentmechanismus:<br />
Die hundertfach erhöhte Ca 2+ -Konzentration bewirkt eine Konformationsänderung des Troponins, das<br />
eine Strukturänderung des Tropomyosins, nun können Myosin und Aktin Kontakt aufnehmen.<br />
Die Myosinköpfchen heften am Aktin an, winkeln sich um 45° ab (ATP notwendig !!) und ziehen dadurch<br />
das Aktin über sich hinweg. Unter ATP-Verbrauch und mit Hilfe von Mg 2+ lösen sich die Köpfchen<br />
und schwingen zurück 1. „Ruderschlag“ ist beendet.<br />
Solange die Ca 2+ -Konzentration erhöht ist, erfolgten viele „Ruderschläge“ das Sarkomer verkürzt<br />
sich der Muskel verkürzt sich.<br />
Nach Reizende werden die Ca 2+ wieder ins Sarkomer zurückgepumpt, das Troponin-Hemmsystem wird wieder<br />
wirksam.<br />
Weichmacherwirkung von ATP / Totenstarre:<br />
Die Bindung der Myorinköpfchen ans Aktin ist starr! ATP löst diese starre Bindung! Fehlt ATP oder sinkt der<br />
ATP-Spiegel bleiben die Muskeln hart. Das ist auch nach dem Tod der Fall.<br />
© Florian Zeller 08 / 09<br />
Seite 237
Biologische Regelung<br />
Begriffe zum Regelkreisschema (am Beispiel des Kühlschranks)<br />
Wer kennt das nicht? Sie reißen die Kühlschranktüre auf, danach brummt der Motor!!!<br />
Führungsgröße: bestimmt den Sollwert (das sind Sie, sie stellen den Thermostat ein)<br />
Sollwert:<br />
ist die Information über den Endzustand der Regelgröße<br />
(also wie es sein soll, z.B. wenn Sie unentschlossen die Kühlschranktüre aufreißen und während Ihrer kulinarischen<br />
Überlegungen die Temperatur im Kühlschrank auf 15 °C ansteigt, dann soll nachher die Temperatur im Kühlschrank<br />
Doch wieder ca. 8°C sein, oder?)<br />
© Florian Zeller 08 / 09<br />
Seite 238
Regelgröße: ist ein konstant zu haltender Vorgang oder Zustand (also was soll geregelt werden, hier die 8°C)<br />
Regelglied: vergleicht Sollwert und Istwert und löst eine Steuerung aus<br />
(der Thermostat im Kühlschrank vergleicht 8°C mit 15°C und löst letztendlich den Kühlschrankmechanismus aus)<br />
Istwert:<br />
Fühler:<br />
Störgröße:<br />
Stellwert:<br />
augenblicklicher Zustand der Regelgröße<br />
(im Moment Temperatur von 15°C)<br />
Messeinrichtung für den Istwert, leitet den Istwert an das Regelglied weiter<br />
(Temperaturfühler im Kühlschrank)<br />
störender Außeneinfluss auf die Regelgröße<br />
(Erwärmung der normalen Kühlschranktemperatur)<br />
Information, was das Stellglied zu tun hat, oder welche Tätigkeit das Stellglied auszulösen hat<br />
(Info, um welchen Betrag die Temperatur verstellt werden muss)<br />
Stellglied: ist der Korrekturmechanismus (der Motor des Kühlschranks)<br />
Stellgröße:<br />
Regelstrecke:<br />
Tätigkeit des Korrekturmechanismus (Laufzeit des Motors des Kühlschranks)<br />
best. Bereich, innerhalb dessen die Regelgröße verändert werden kann<br />
(Der Kühlschrank kann innerhalb eines bestimmten Temperaturbereiches kühlen, aber Sie können ihn nicht als<br />
Backofen verwenden)<br />
© Florian Zeller 08 / 09<br />
Seite 239
Regelung der Pupillenöffnung<br />
Durch die Beleuchtungsstärke<br />
Pfeildiagramm:<br />
Lichteinfall (Ursache)<br />
-<br />
+<br />
Verengung der Pupille (Wirkung)<br />
Reitz und Reaktion bzw. Ursache und Wirkung sind zu einem Kausalkreis mit negativer Rückkoppelung<br />
geschlossen<br />
Regelung über das Nervensystem<br />
1. Regelung der Atemgaskonzentration im Blut<br />
Pfeildiagramm:<br />
+<br />
CO 2 -Gehalt im Blut<br />
-<br />
Atemtätigkeit<br />
Kausalkreis mit negativer Rückkoppelung<br />
Regelstrecke Blut<br />
© Florian Zeller 08 / 09<br />
Seite 240
2. Regelung der Körpertemperatur<br />
Pfeildiagramm:<br />
zweiseitige Regelung (Körpertemp. Erhöhen & senken)<br />
Muskelzittern zur<br />
Wärmeproduktion<br />
Verengung der<br />
Blutgefäße<br />
(Grundumsatz<br />
erhöht sich)<br />
-<br />
+<br />
Außentemperatur<br />
+<br />
Körpertemperatur<br />
-<br />
+<br />
Schweißabsonderung<br />
Erweiterung der Blut-<br />
Kapillaren<br />
(Senkung des Grundumsatzes<br />
(GU) )<br />
© Florian Zeller 08 / 09<br />
Seite 241
Hormone<br />
Hormone sind Botenstoffe, die der Körper selbst erzeugt. Sie sind bereits in geringsten Mengen wirksam.<br />
Sie stellen neben dem Nervensystem ein zweites Informationssystem dar.<br />
Chemischer Bau<br />
1. Steroide (Sexualhormone)<br />
2. Aminosäurederivate (Thyroxin, Adrenalin) uneinheitlich<br />
3. Peptide / Proteine (Insulin, Glucagon)<br />
Orte der Produktion:<br />
1. Endokrine (innersekretorische) Drüsen<br />
(Hypophyse, Schilddrüse, Thymus, Pankreas,<br />
Nebennieren, Keimdrüsen)<br />
2. Gewebshormone<br />
Hypothalamus bildet Releasing-Hormone<br />
Haut bildet Histamine<br />
Drüsen Hormon Funktion<br />
Hypophyse Tyhreotropin (TSH) Stimuliert Schilddrüse<br />
Schilddrüse Thyroxin Stimuliert Stoffwechselprozesse<br />
Wirkt auf Grundumsatz<br />
Nebennierenrinde<br />
Glucocorticoide<br />
heben glucosespiegel im Blut<br />
Nebennierenmark<br />
Pankreas<br />
Adrenalin<br />
Insulin<br />
Glucagon<br />
Stresshormon, erhöht Stoffwechsel<br />
senkt Glucosespiegel im Blut<br />
hebt Glucosespielgel im Blut<br />
Zusammenhang zwischen Hormonsystem und Nervensystem<br />
Hormonsystem und Nervensystem sind bei der Aufrechterhaltung der Homöostase (inneres Gleichgewicht), der<br />
Entwicklung und der Fortpflanzung oft untrennbar verwoben.<br />
1. Strukturelle Beziehungen: Der Hypothalamus und die Hypophyse bestehen aus Nervengewebe, das<br />
Nebennierenmark hat sich aus Nervengewebe entwickelt.<br />
2. Chemische Beziehung: Mehrere Hormone werden auch als Neurotransmitter verwendet (z.B. Adrenalin).<br />
3. Funktionelle Beziehung: Viele Körperfunktion werden von beiden Systemen reguliert (z.B. Stressreaktionen,<br />
Säugen der Jungen).<br />
© Florian Zeller 08 / 09<br />
Seite 242
Regelung über das Hormonsystem<br />
Regelung des Grundumsatzes bzw. des Schilddrüsen-Hormons Thyroxin<br />
Pfeildiagramm:<br />
Hypothalamus<br />
Neurohormon TRH<br />
-<br />
-<br />
+<br />
Hypophyse<br />
Steuerhormon TSH<br />
+<br />
-<br />
Thyroxinkonzentration<br />
im Blut<br />
+<br />
Grundumsatz, Höhe<br />
des Energiestoffwechsels<br />
und der<br />
Wärmeproduktion<br />
+<br />
SCHILDDRÜSE<br />
THYROXIN<br />
Regelung der Zuckerkonzentration im Blut<br />
durch Adrenalin Insulin: Glycogenaufbau (holt Zucker aus dem Blut)<br />
Glycogenabbau zu Glucose<br />
Glucagon<br />
Pfeildiagramm:<br />
Glucagonfreisetzung -<br />
+<br />
Glycogenzerlegung<br />
Muskeltätigkeit<br />
-<br />
+<br />
Glucosespiegel<br />
im Blut<br />
Kohlenhydratverdauung<br />
(Zufuhr von Glucose)<br />
+<br />
-<br />
+<br />
Insulinfreisetzung<br />
+<br />
Glycogenaufbau<br />
2 Regler beteiligt: 1. Hypothalamus + Hypophyse<br />
2. Bauchspeicheldrüse<br />
© Florian Zeller 08 / 09<br />
Seite 243
© Florian Zeller 08 / 09<br />
Seite 244