

Water Management Scenarios and Clams - Jan Thompson, U.S.G.S.
Water Management Scenarios and Clams - Jan Thompson, U.S.G.S.
Water Management Scenarios and Clams - Jan Thompson, U.S.G.S.

You also want an ePaper? Increase the reach of your titles
YUMPU automatically turns print PDFs into web optimized ePapers that Google loves.
Chlorophyll a (μg/L)<br />
60<br />
50<br />
40<br />
30<br />
20<br />
10<br />
0<br />
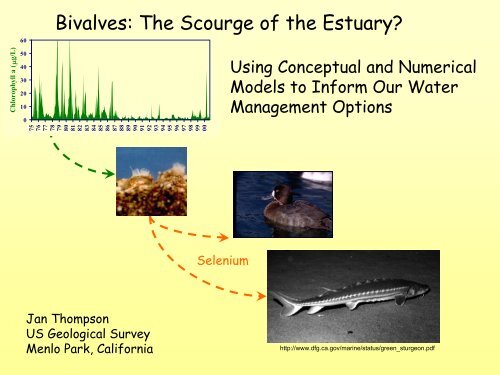
Bivalves: The Scourge of the Estuary?<br />
75<br />
76<br />
77<br />
78<br />
79<br />
80<br />
81<br />
82<br />
83<br />
84<br />
85<br />
86<br />
87<br />
88<br />
89<br />
90<br />
91<br />
92<br />
93<br />
94<br />
95<br />
96<br />
97<br />
98<br />
99<br />
00<br />
<strong>Jan</strong> <strong>Thompson</strong><br />
US Geological Survey<br />
Menlo Park, California<br />
Selenium<br />
Using Conceptual <strong>and</strong> Numerical<br />
Models to Inform Our <strong>Water</strong><br />
<strong>Management</strong> Options<br />
http://www.dfg.ca.gov/marine/status/green_sturgeon.pdf
My charge<br />
Any good models on the shelf? No<br />
How can we use conceptual models like those<br />
developed in DRERIP? Strengths? Weaknesses?<br />
How do conceptual models differ from numerical<br />
models being developed in CASCADE? Strengths?<br />
Weaknesses?<br />
What’s the future for bivalve models?<br />
Status:<br />
Invaded 1986 <strong>and</strong> limited laboratory studies<br />
DRERIP conceptual model<br />
STELLA population model – not dynamically linked<br />
Invaded 1940 <strong>and</strong> more laboratory studies<br />
DRERIP conceptual model<br />
STELLA population model – not dynamically linked
Bivalve models are connected to many submodels<br />
but must be connected to hydrodynamic models<br />
at a minimum to<br />
Distribute gametes <strong>and</strong> young<br />
Maintain/change water quality<br />
Deliver food to the sedentary animals
Conceptual models such as DRERIP<br />
models are critical for:<br />
Designing numerical models – particularly<br />
valuable in making connections between bivalves<br />
<strong>and</strong> other factors (physical transport & mixing,<br />
phytoplankton etc)<br />
Highlighting data needs <strong>and</strong> prioritizing<br />
research needs <strong>and</strong> dollars<br />
Examining processes which may not be modeled<br />
in the near term
DRERIP: We have developed Corbul <strong>and</strong> Corbicula Life<br />
Cycle Models, the first step in building conceptual models<br />
Reproductive<br />
Adult<br />
(5mm long)<br />
Recruit<br />
(135 µm long)<br />
17-19 days<br />
Corbula<br />
Pelagic<br />
Veliger<br />
Larvae*<br />
Spawning &<br />
Fertilization<br />
Pelagic<br />
Trochophore<br />
Larvae*<br />
*From Nicolini, MH <strong>and</strong> DL Penry. 2000. Spawning, fertilization, <strong>and</strong> larval development of Potamocorbula amurensis (Mollusca: Bivalvia) from San Francisco Bay, California. Pacific Science:<br />
54(4):377-388 NO PERMISSION FOR PUB.<br />
24 hours<br />
Gametogenesis<br />
24 hours
DRERIP sub-models have been connected to each Life<br />
Cycle Model <strong>and</strong> coded for habitat <strong>and</strong> critical processes<br />
physical processes dominate<br />
Adult<br />
Transport<br />
Model<br />
Mortality<br />
Model<br />
Reproductive<br />
Adult<br />
(5mm long)<br />
Growth<br />
Model<br />
Recruit<br />
(135 µm long)<br />
Larval<br />
Recruit<br />
Model<br />
benthic habitat<br />
17-19 days<br />
pelagic habitat<br />
Pelagic<br />
Veliger<br />
Larvae*<br />
Mortality<br />
Model<br />
Gametogenesis<br />
physical processes critical<br />
Spawning &<br />
Fertilization<br />
Pelagic<br />
Trochophore<br />
Larvae*<br />
Larval<br />
Transport<br />
Model<br />
Reproduction<br />
Model<br />
*From Nicolini, MH <strong>and</strong> DL Penry. 2000. Spawning, fertilization, <strong>and</strong> larval development of Potamocorbula amurensis (Mollusca: Bivalvia) from San Francisco Bay, California. Pacific Science:<br />
54(4):377-388 NO PERMISSION FOR PUB.<br />
24 hours<br />
24 hours
Each DRERIP sub-model includes triggers or forcing<br />
factors <strong>and</strong> stressors<br />
Food Quantity<br />
Food Quality<br />
<strong>Water</strong> Quality<br />
Toxics<br />
Adult Health<br />
<strong>Water</strong> Quality<br />
Adult Health<br />
<strong>Water</strong> Quality<br />
Gametogenesis<br />
Internal<br />
Fertilization<br />
Release Brood<br />
el.erdc.usace.army<br />
Gametogenesis<br />
Broadcast<br />
Spawning<br />
External<br />
Fertilization<br />
Pelagic<br />
Trochophore<br />
Larvae<br />
Food Quantity<br />
Food Quality<br />
<strong>Water</strong> Quality<br />
Toxics<br />
Adult Health<br />
Stress/ no food<br />
<strong>Water</strong> Quality<br />
<strong>Water</strong> Quality<br />
Transport
DRERIP models help us build numerical models AND tell<br />
us what we know, how well we know it, <strong>and</strong> what critical<br />
data are lacking<br />
Importance<br />
High<br />
Medium<br />
Low<br />
Underst<strong>and</strong>ing<br />
High<br />
Medium<br />
Low<br />
Predictability<br />
High<br />
Medium<br />
Low<br />
Food Quantity<br />
Food Quality<br />
<strong>Water</strong> Quality<br />
Toxics<br />
Adult Health<br />
<strong>Water</strong> Quality<br />
Adult Health<br />
<strong>Water</strong> Quality<br />
Gametogenesis<br />
Internal<br />
Fertilization<br />
Release Brood<br />
el.erdc.usace.army
A similar exercise for Corbula shows different data gaps<br />
<strong>and</strong> needs<br />
Importance<br />
High<br />
Medium<br />
Low<br />
Underst<strong>and</strong>ing<br />
High<br />
Medium<br />
Low<br />
Predictability<br />
High<br />
Medium<br />
Low<br />
Gametogenesis<br />
Broadcast<br />
Spawning<br />
External<br />
Fertilization<br />
Pelagic<br />
Trochophore<br />
Larvae<br />
Food Quantity<br />
Food Quality<br />
<strong>Water</strong> Quality<br />
Toxics<br />
Adult Health<br />
Stress/ no food<br />
<strong>Water</strong> Quality<br />
<strong>Water</strong> Quality<br />
Transport
Salinity<br />
Food<br />
35<br />
30<br />
25<br />
20<br />
15<br />
10<br />
5<br />
0<br />
Year Class 1 Gametogenesis<br />
underst<strong>and</strong>ing of controls on a species distribution<br />
0<br />
Spawn<br />
Fertilization<br />
Days<br />
1 2 17-19 80+ Fall<br />
Spring Extended Life Cycle Models further our<br />
Pelagic Larva<br />
Pelagic Swimming Larva<br />
Settled Juvenile<br />
Some times &<br />
places >1/season<br />
Pelagic Infaunal<br />
Reproductively Mature Adult<br />
Year Class 2 Gametogenesis
Temperature<br />
Food<br />
35<br />
30<br />
25<br />
20<br />
15<br />
10<br />
5<br />
0<br />
Year Class 1 Oogenesis<br />
And allow us to contrast controls on different<br />
species<br />
Always<br />
Spring<br />
Spermatogenesis<br />
0<br />
Self -Fertilization<br />
Pediveliger<br />
Days<br />
Fall<br />
5 7 12 17 19<br />
90+<br />
Spermatogenesis<br />
Self -Fertilization<br />
Pediveliger<br />
Spermatogenesis<br />
Stop when ≈26°C<br />
or insufficient food<br />
Epibenthic/Pelagic Infaunal<br />
Juveniles Reproductively Mature<br />
Year Class 2 Oogenesis<br />
Spermatogenesis
DRERIP can help us discover critical processes<br />
before we are ready to model them. Eg. Eg.<br />
Why do<br />
the clams disappear in winter? Does it matter?<br />
Net Outflow (m 3 /s)<br />
600000<br />
500000<br />
400000<br />
300000<br />
200000<br />
100000<br />
0<br />
Grizzly Bay Corbula<br />
88 89 90 91 92 93 94 95 96 97 98 99 00 01 02 03 04 05 06<br />
Samples courtesy of DWR (D7)<br />
0.7<br />
0.6<br />
0.5<br />
0.4<br />
0.3<br />
0.2<br />
0.1<br />
0.0<br />
Corbula Biomass<br />
(g AFDW/m 2 )
Migratory ducks heavily prey upon<br />
shallow water Corbula Corbula <strong>and</strong> they are<br />
bio-accumulating bio accumulating Se.<br />
Mortality<br />
Benthic Predation<br />
Pelagic Larvae<br />
Predation<br />
Starvation<br />
Burial<br />
<strong>Water</strong> Quality Stress<br />
Natural Death - Age<br />
Fish<br />
Resident Birds<br />
Migratory Birds<br />
Invertebrates<br />
Corbula<br />
Mortality<br />
Model<br />
White Sturgeon<br />
Sacramento Splittail<br />
Ducks<br />
Opisthobranchs<br />
Dungeness Crab<br />
physical processes<br />
dominate<br />
physical processes<br />
critical<br />
biological variables
However DRERIP models can’t can t explain the inter-<br />
annual variability in biomass (<strong>and</strong> thus grazing)<br />
peaks<br />
Net Outflow (m 3 /s)<br />
600000<br />
500000<br />
400000<br />
300000<br />
200000<br />
100000<br />
0<br />
Grizzly Bay Corbula<br />
88 89 90 91 92 93 94 95 96 97 98 99 00 01 02 03 04 05 06<br />
Samples courtesy of DWR (D7)<br />
0.7<br />
0.6<br />
0.5<br />
0.4<br />
0.3<br />
0.2<br />
0.1<br />
0.0<br />
Corbula Biomass<br />
(g AFDW/m 2 )
DRERIP models also can’t can t tell us why Corbicula Corbicula<br />
juveniles occur throughout the Delta but only<br />
grow-up grow up in the central Delta.
To do that we need to look at growth<br />
models <strong>and</strong> it becomes obvious that we<br />
need more than a conceptual model.<br />
Tissue Growth<br />
Energy for<br />
Growth<br />
Carbon Available<br />
for Assimilation<br />
Growth<br />
(increase in<br />
Carbon)<br />
Shell Growth<br />
Energy for<br />
Reproduction<br />
Energy for<br />
Respiration<br />
Energy for<br />
Excretion<br />
Consumption<br />
Rate<br />
Physiological<br />
Stressors<br />
Food Quality<br />
Filtration Efficiency<br />
Filtration Rate<br />
<strong>Water</strong> Quality<br />
Food Quantity<br />
Suspended Sediment<br />
Population Size of<br />
Filter Feeders<br />
Current Speed<br />
Behavior<br />
Pumping Rate<br />
Zooplankton<br />
Phytoplankton<br />
Bacteria<br />
DOM<br />
Zooplankton<br />
Phytoplankton<br />
Bacteria<br />
DOM<br />
Corbula<br />
Growth<br />
Model<br />
<strong>Water</strong> Quality<br />
physical processes<br />
dominate<br />
physical processes<br />
critical<br />
biological variables<br />
Species<br />
Size<br />
Species<br />
Size<br />
Attached/Free?<br />
Size
In Summary, conceptual models such<br />
as DRERIP models do not<br />
Tell us the magnitude of the bivalves ecological<br />
functions (eg. consuming phytoplankton)<br />
Tell us the temporal <strong>and</strong> spatial variability of<br />
those ecological functions, the causes of which<br />
might be exploited by water management<br />
Tell us if the model is wrong – surprises are when<br />
we learn<br />
Allow us to dynamically link <strong>and</strong> manipulate<br />
variables
Numerical models within CASCADE<br />
We are using STELLA models until we know<br />
more – adapted to be spatially variable <strong>and</strong><br />
responsive to stressors<br />
Will tell us about processes <strong>and</strong> limits on<br />
populations – I will discuss some today<br />
Allow us to dynamically link <strong>and</strong> manipulate some<br />
variables<br />
We expect <strong>and</strong> look forward to surprises
We are simplifying STELLA models to operate with<br />
statistical relationships instead of using energetic<br />
principles.<br />
St<strong>and</strong>ard Stella<br />
Energetic Growth<br />
Model<br />
Statistical Stella<br />
Growth Model<br />
Adapted from Grant <strong>and</strong> Bacher 1998
Spatial resolution <strong>and</strong> physical variables known to<br />
dynamically interact with clam functions will be run<br />
sequentially to produce a “look up” table for other<br />
models.<br />
habitat<br />
1<br />
.<br />
.<br />
.<br />
.<br />
.<br />
.<br />
.<br />
20<br />
1 . . . . . 5<br />
phytoplankton<br />
biomass<br />
1 . . . . . 4<br />
temperature<br />
Output: Temporally <strong>and</strong> spatially<br />
varying growth rate, biomass,<br />
grazing rate for phytoplankton <strong>and</strong><br />
contaminant models
Step 1: Initial Conditions: Where are the two species in<br />
space? Corbula recruits settle downstream of X2<br />
(Corbicula juveniles settle upstream of X2!)<br />
X2 at 4.1 # Recruits/0.05m2<br />
physical processes dominate<br />
# Adults/0.05m 2<br />
1000<br />
900<br />
800<br />
700<br />
600<br />
500<br />
400<br />
300<br />
200<br />
100<br />
0<br />
200<br />
180<br />
160<br />
140<br />
120<br />
100<br />
80<br />
60<br />
40<br />
20<br />
0<br />
station is at X2 of 72; as<br />
long as X2 meets or exceeds<br />
that level we get recruits<br />
40 45 50 55 60 65 70 75 80 85 90 95 100<br />
Older year class shows wider<br />
spread which we expect<br />
because they are tolerant of<br />
lower salinities<br />
40 45 50 55 60 65 70 75 80 85 90 95 100
In order to establish some statistical bounds to our<br />
results, we will use an ensemble of salinity <strong>and</strong><br />
temperature distributions in time <strong>and</strong> space to establish<br />
a range of distributions for each scenario<br />
Δ<strong>Water</strong> Temp / ΔAir Temp<br />
0.65<br />
0.6<br />
0.55<br />
0.5<br />
0.45<br />
0.4<br />
-122.3 -122.2 -122.1 -122 -121.9 -121.8<br />
Longitude<br />
-121.7 -121.6 -121.5 -121.4 -121.3<br />
Grantline Tracy<br />
SJ Antioch<br />
Middle Tracy<br />
SJ Prisoner<br />
Wickl<strong>and</strong> Pier<br />
Sac Martinez<br />
Stockton Burns<br />
Old Tracy<br />
SJ Navy Bridge<br />
Sac Mallard<br />
SJ Mossdale<br />
Sac Rio Vista<br />
Sac Hood<br />
Preliminary data from Wayne Wagner & Mark Stacey
physical processes critical<br />
biological variables<br />
Initial Conditions: Number of recruits is<br />
partially dependent on the biomass of adults<br />
present; there is a physical <strong>and</strong> biological basis<br />
for this relationship<br />
# Recruits/0.05m 2<br />
1400<br />
1200<br />
1000<br />
800<br />
600<br />
400<br />
200<br />
0<br />
0 0.2 0.4 0.6 0.8<br />
Biomass Adults/0.05m 2
Net Outflow (m 3 /s)<br />
600000<br />
500000<br />
400000<br />
300000<br />
200000<br />
100000<br />
We assign Corbula recruit abundance based on<br />
biomass of adults <strong>and</strong> antecedent outflow<br />
events – note the grouping of abundances<br />
around 100, 200 <strong>and</strong>
# Recruits ( ≤2.5 mm/0.05m 2 )<br />
# Recruits ( ≤2.5 mm/0.05m 2 )<br />
80<br />
70<br />
60<br />
50<br />
40<br />
30<br />
20<br />
10<br />
0<br />
350<br />
300<br />
250<br />
200<br />
150<br />
100<br />
50<br />
0<br />
Apr<br />
<strong>Jan</strong>-98<br />
May<br />
Apr-98<br />
We observe Corbicula recruit abundance at<br />
≈20 recruits/0.05 m2 throughout the year at<br />
monitoring stations<br />
Franks Tract Near Old River<br />
Jun<br />
Jul-98<br />
Jul<br />
Oct-98<br />
Aug<br />
<strong>Jan</strong>-99<br />
Sep<br />
Apr-99<br />
Oct<br />
Jul-99<br />
Nov<br />
Oct-99<br />
Dec<br />
<strong>Jan</strong>-00<br />
<strong>Jan</strong><br />
Sacramento River Near Rio Vista<br />
No data<br />
Apr-00<br />
Feb<br />
Jul-00<br />
Mar<br />
Oct-00<br />
We are determining if transport<br />
of larvae from non-local sites,<br />
where temperature regimes may<br />
be staggered, accounts for the<br />
near continuous pattern of<br />
recruitment observed in areas like<br />
the central delta where many<br />
sources of water are mixed.<br />
If so changing temperatures<br />
throughout the tributaries <strong>and</strong><br />
Delta could alter this population<br />
distribution<br />
physical processes critical
Corbicula’s present <strong>and</strong> potential biomass<br />
distribution is likely based on food limitation so a<br />
growth relationship is critical to our model<br />
Feb-Dec 1981<br />
Foe & Knight 1985<br />
Corbicula growth is food limited at<br />
20 mg/L chlorophyll a at 15°C <strong>and</strong> 47<br />
mg/L at 20-24°C. Growth was limited<br />
in the San Joaquin River except for two<br />
months in 1981 so this relationship is<br />
probably as good as is available for this<br />
system<br />
Chlorophyll a<br />
(mg/L)<br />
30<br />
25<br />
20<br />
15<br />
10<br />
5<br />
0<br />
San Joaquin River at Antioch Ship Channel<br />
<strong>Jan</strong> Feb Mar Apr Ma<br />
y<br />
Jun Jul Aug Sep Oct Nov Dec<br />
physical processes critical
Shell Length<br />
Growth Rate<br />
(mm/d)<br />
We believe that Corbula is also food limited in<br />
this system but we do not have laboratory data<br />
similar to that for Corbicula to verify our beliefs.<br />
0.08<br />
0.07<br />
0.06<br />
0.05<br />
0.04<br />
0.03<br />
0.02<br />
0.01<br />
0<br />
y = 0.0121e 0.6072x<br />
R 2 = 0.9791<br />
Recruits in Grizzly Bay<br />
0 0.5 1 1.5 2 2.5 3<br />
Average Chlorophyll a (mg/L)<br />
Our preliminary analyses of field data confirm food<br />
limitation in bivalves in the shallow water.
Summary<br />
Our skill with both bivalve models is limited by<br />
data<br />
Data gaps <strong>and</strong> priorities are well summarized by<br />
conceptual models (DRERIP)<br />
Developing relationships for use in the numerical<br />
models is ongoing <strong>and</strong> is helping us define important<br />
processes <strong>and</strong> thresholds<br />
Given the best data <strong>and</strong> models, ecologists are<br />
still struggling with multi-generation, spatially<br />
explicit population models of benthic animals. We<br />
also expect to have problems so we are taking it<br />
slow, linking sub-models one step at a time
Net Outflow (m 3 /s)<br />
Net Outflow (m 3 /s)<br />
600000<br />
500000<br />
400000<br />
300000<br />
200000<br />
100000<br />
0<br />
600000<br />
500000<br />
400000<br />
300000<br />
200000<br />
100000<br />
0<br />
800+<br />
87 88 89 90 91 92 93 94 95 96 97 98 99 00 01 02 03 04 05 06<br />
87 88 89 90 91 92 93 94 95 96 97 98 99 00 01 02 03 04 05 06<br />
500<br />
400<br />
300<br />
200<br />
100<br />
0<br />
0.7<br />
0.6<br />
0.5<br />
0.4<br />
0.3<br />
0.2<br />
0.1<br />
0.0<br />
# Recruits (/0.05m 2 )<br />
Adult Corbula Biomass<br />
(g AFDW/m 2 )














