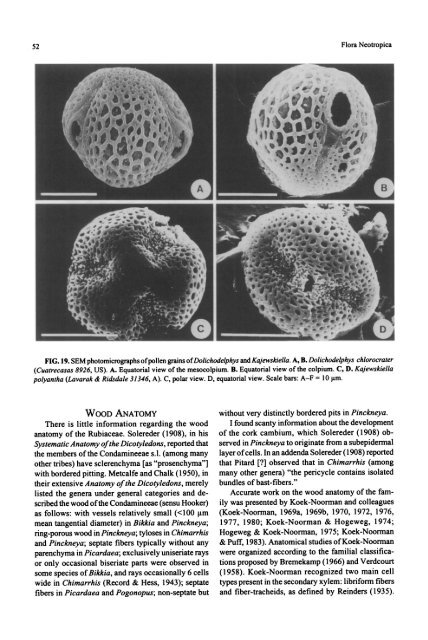
ass 52 Flora Neotropica I *:J ~Zl:, ,Mt ... A ............ .. +_ B ..9, I _ ' FI.1.SM htmcorah a Ka n eskiela foln A, riso olcoepy oihdlhsclrcae 896 qao S.A ial~ temsolium viwo theclpu.C, .Eqaoiliwo I.Kewila polyanth (Lvrk&Rddl 1346,: A. polrviwD,eutria cl iw as - C, 0Im FIG. 19. SEM photomicrographs ofvpollen grains ofDolichodelphys and Kajewskiella. A, B.Dolichodelphys chiorocrater (Cuatrecasas 8926, US). A. Equatorial view of the mesocolpium. B. Equatorial view of the colpium. C, D. Kajewskiella polyantha (Lavarak & Ridsdale 31346, A). C, polar view. D, equatorial view. Scale bars: A-F = 10 gtm. WOOD ANATOMY There is little information regarding the wood anatomy of the <strong>Rubiaceae</strong>. Solereder (1908), in his Systematic Anatomy ofthe Dicotyledons, reported that the members of the Condamineeae s.l. (among many other tribes) have sclerenchyma [as "prosenchyma"] with bordered pitting. Metcalfe and Chalk (1950), in their extensive Anatomy of the Dicotyledons, merely listed the genera under general categories and described the wood of the Condamineeae (sensu Hooker) without very distinctly bordered pits in Pinckneya. I found scanty information about the development of the cork cambium, which Solereder (1908) observed in Pinckneya to originate from a subepidermal layer of cells. In an addenda Solereder (1908) reported that Pitard [?] observed that in Chimarrhis (among many other genera) "the pericycle contains isolated bundles of bast-fibers." Accurate work on the wood anatomy of the family was presented by Koek-Noorman and colleagues as follows: with vessels relatively small (
Introduction to the Neotropical Genera Studied 53 Koek-Normann (1970, 1972), Koek-Noorman and transferred to the Condamineeae by Andersson and Hogeweg (1974), and Koek-Noorman and Puff(1983) Persson (1991). Coutarea has recently been transreported that the tribes Gardenieae, Ixoreae, Vangueri- ferred to the Catesbaeeae (Delprete, 1995a, 1996d), and eae, Isertieae, Naucleae, Mussaendeae, Hedyotideae, Exostema was treated as a closely related taxon (Delprete, Anthospermeae, and Rubieae are all homogeneous in 1995a, 1996d; McDowell, 1996, and pers. comm.). having only fiber-tracheids, while Psychotrieae, Pae- Koek-Noorman and Hogeweg (1974) performed derieae, Morindeae, Guettardeae, Hillieae, and Hame- a cluster analysis using 125 wood anatomical characlieae are homogeneous in having only libriform fibers. ters, but a complete data matrix of such characters was Koek-Noorman (1969b), in her review of the Ru- not published. Three dendrograms for the Cinchoneae, biaceae of Suriname, discussed the wood anatomy and <strong>Rondeletieae</strong>, and Condamineeae were produced, and taxonomy of Chimarrhis as follows: "Schumann was they all displayed a basal "partition in two groups, one not sure whether C. turbinata was rightly placed in of which shows libriform fibers, the other one fiber this genus. Ducke (1922) referred it to a new genus, tracheids." In the Condamineeae they reported that viz. Pseudochimarrhis. According to Bremekamp "the parenchyma pattern seems to be less connected (1934) and Steyermark (1965) Chimarrhis and with the fiber type, the arrangement of the vessels, and Pseudochimarrhis are not separable. Record and Hess the presence of crystal sand." Bikkia, Morierina, and (1943) report that the wood.of C. turbinata greatly Portlandia [which were transferred to the Catesbaeeae resembles C. longistipulata [= C. microcarpa], the by Delprete (1996d)] formed one cluster. Bikkia camplace of which is not in doubt. They differ only in panulata is reported as "dissimilar in the frequency minor features." In the same article Koek-Noorman of radial vessel multiples, relatively large intervascu- (1969b) described the wood of Chimarrhis as having lar pits, uniseriate rays, relatively large ray-vessel septate fibers, narrowly vasicentric parenchyma, with pitting, presence of crystal sand, and vague growth 6-12 vessels per mm2 and up to 150 pm wide, upright rings." But this set of characters could be influenced ray cells up to 50 um, without sheath cells nor broad by environmental factors. rays. As a conclusion she states that the wood of The monotypic Pinckneya was pointed out by Chimarrhis resembles that of the Cinchoneae (of Koek-Noorman and Hogeweg (1974) as exceptional which she studied Capirona, Ferdinandusa, and Cin- to the Condamineeae (sensu Hooker) in having semichona) more than that of the <strong>Rondeletieae</strong> (of which ring-porous wood, with tangential pore chains and she studied only Elaeagia and Warszewiczia). concentric parenchyma bands. They did not mention Koek-Noorman and Hogeweg (1974) presented a that this is the only non-tropical genus analyzed (ensurvey of some representative taxa of the tribes Van- demic to Georgia and South Carolina, North America) guerieae, Cinchoneae, Condamineeae, and <strong>Rondeletieae</strong> and that its wood anatomy is certainly influenced by (all sensu Bremekamp, 1966), which is the only study the seasonal climate to which this genus is exposed. in which representative genera of the Condamineeae Considering the dry-seasonal forests to which Pogonwere analyzed. The genera of the Condamineeae opus is endemic, I would also expect its wood to be (sensu Bremekamp, 1966) analyzed by Koek-Noorman ring-porous (not reported by Koek-Noorman). and Hogeweg (1974) were Condaminea (C. corym- Pogonopus and <strong>Rustia</strong> (with Calycophyllum and bosa), <strong>Rustia</strong> (R. formosa), Chimarrhis (C. cymosa, Mussaendopsis) were reported to have fibers clearly C. microcarpa [as C. longistipulata], C. turbinata, C. septate and with "scanty" pits on their tangential walls. hookeri), Pogonopus (P. speciosus), Pinckneya (P. bracteata [as P. pubens]), Bikkia (B. campanulata, B. palauensis), Portlandia (P. grandiflora), and Morierina (M. montana). They reported that the Cinchoneae, <strong>Rondeletieae</strong>, and Condamineeae were rather heterogeneous Finally, Koek-Noorman and Hogeweg (1974) performed two cluster analyses of the Vanguerieae, Cinchoneae, <strong>Rondeletieae</strong>, and Condamineeae combined, one with equal character weighting and the other with preferential character weighting. Both in having both libriform fibers and fiber-tracheids. analyses also showed a basal bipartition so that gen- This finding is probably due to the poor delimitation era of various tribes were scattered on the dendroof the three tribes at that time. Unfortunately, the au- grams, and sometimes species of the same genus were thors discussed only the characteristics of the genera even placed on remote branches. According to these exceptional to the Condamineeae, and the descriptions two analyses, the authors concluded that none of the of wood anatomical characters for most genera stud- characters studied has reliable diagnostic value, and ied (<strong>Rustia</strong>, Condaminea, Pogonopus) are only frag- they were persuaded "not to pursue the attempt to mentary. Koek-Noormann and Hogeweg (1974) re- define the taxonomic tribes under discussion [Ronported Exostema and Coutarea to be exceptional for deletieae, Condamineeae, and Cinchoneae as defined the tribe Cinchoneae. The two genera were subsequently at that time] on the base of wood anatomy."