

Pollen and Stigma Structure and Function: The Role of Diversity in ...
Pollen and Stigma Structure and Function: The Role of Diversity in ...
Pollen and Stigma Structure and Function: The Role of Diversity in ...

You also want an ePaper? Increase the reach of your titles
YUMPU automatically turns print PDFs into web optimized ePapers that Google loves.
S90<br />
<strong>The</strong> Plant Cell<br />
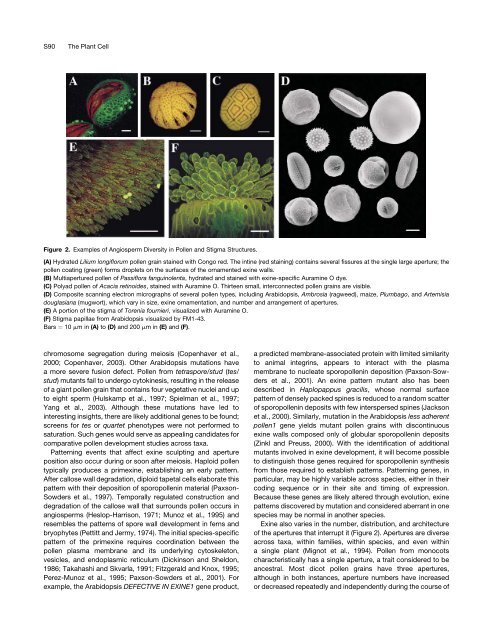
Figure 2. Examples <strong>of</strong> Angiosperm <strong>Diversity</strong> <strong>in</strong> <strong>Pollen</strong> <strong>and</strong> <strong>Stigma</strong> <strong>Structure</strong>s.<br />
(A) Hydrated Lilium longiflorum pollen gra<strong>in</strong> sta<strong>in</strong>ed with Congo red. <strong>The</strong> <strong>in</strong>t<strong>in</strong>e (red sta<strong>in</strong><strong>in</strong>g) conta<strong>in</strong>s several fissures at the s<strong>in</strong>gle large aperture; the<br />
pollen coat<strong>in</strong>g (green) forms droplets on the surfaces <strong>of</strong> the ornamented ex<strong>in</strong>e walls.<br />
(B) Multiapertured pollen <strong>of</strong> Passiflora fangu<strong>in</strong>olenta, hydrated <strong>and</strong> sta<strong>in</strong>ed with ex<strong>in</strong>e-specific Auram<strong>in</strong>e O dye.<br />
(C) Polyad pollen <strong>of</strong> Acacia ret<strong>in</strong>oides, sta<strong>in</strong>ed with Auram<strong>in</strong>e O. Thirteen small, <strong>in</strong>terconnected pollen gra<strong>in</strong>s are visible.<br />
(D) Composite scann<strong>in</strong>g electron micrographs <strong>of</strong> several pollen types, <strong>in</strong>clud<strong>in</strong>g Arabidopsis, Ambrosia (ragweed), maize, Plumbago, <strong>and</strong> Artemisia<br />
douglasiana (mugwort), which vary <strong>in</strong> size, ex<strong>in</strong>e ornamentation, <strong>and</strong> number <strong>and</strong> arrangement <strong>of</strong> apertures.<br />
(E) A portion <strong>of</strong> the stigma <strong>of</strong> Torenia fournieri, visualized with Auram<strong>in</strong>e O.<br />
(F) <strong>Stigma</strong> papillae from Arabidopsis visualized by FM1-43.<br />
Bars ¼ 10 mm <strong>in</strong>(A) to (D) <strong>and</strong> 200 mm <strong>in</strong>(E) <strong>and</strong> (F).<br />
chromosome segregation dur<strong>in</strong>g meiosis (Copenhaver et al.,<br />
2000; Copenhaver, 2003). Other Arabidopsis mutations have<br />
a more severe fusion defect. <strong>Pollen</strong> from tetraspore/stud (tes/<br />
stud) mutants fail to undergo cytok<strong>in</strong>esis, result<strong>in</strong>g <strong>in</strong> the release<br />
<strong>of</strong> a giant pollen gra<strong>in</strong> that conta<strong>in</strong>s four vegetative nuclei <strong>and</strong> up<br />
to eight sperm (Hulskamp et al., 1997; Spielman et al., 1997;<br />
Yang et al., 2003). Although these mutations have led to<br />
<strong>in</strong>terest<strong>in</strong>g <strong>in</strong>sights, there are likely additional genes to be found;<br />
screens for tes or quartet phenotypes were not performed to<br />
saturation. Such genes would serve as appeal<strong>in</strong>g c<strong>and</strong>idates for<br />
comparative pollen development studies across taxa.<br />
Pattern<strong>in</strong>g events that affect ex<strong>in</strong>e sculpt<strong>in</strong>g <strong>and</strong> aperture<br />
position also occur dur<strong>in</strong>g or soon after meiosis. Haploid pollen<br />
typically produces a primex<strong>in</strong>e, establish<strong>in</strong>g an early pattern.<br />
After callose wall degradation, diploid tapetal cells elaborate this<br />
pattern with their deposition <strong>of</strong> sporopollen<strong>in</strong> material (Paxson-<br />
Sowders et al., 1997). Temporally regulated construction <strong>and</strong><br />
degradation <strong>of</strong> the callose wall that surrounds pollen occurs <strong>in</strong><br />
angiosperms (Heslop-Harrison, 1971; Munoz et al., 1995) <strong>and</strong><br />
resembles the patterns <strong>of</strong> spore wall development <strong>in</strong> ferns <strong>and</strong><br />
bryophytes (Pettitt <strong>and</strong> Jermy, 1974). <strong>The</strong> <strong>in</strong>itial species-specific<br />
pattern <strong>of</strong> the primex<strong>in</strong>e requires coord<strong>in</strong>ation between the<br />
pollen plasma membrane <strong>and</strong> its underly<strong>in</strong>g cytoskeleton,<br />
vesicles, <strong>and</strong> endoplasmic reticulum (Dick<strong>in</strong>son <strong>and</strong> Sheldon,<br />
1986; Takahashi <strong>and</strong> Skvarla, 1991; Fitzgerald <strong>and</strong> Knox, 1995;<br />
Perez-Munoz et al., 1995; Paxson-Sowders et al., 2001). For<br />
example, the Arabidopsis DEFECTIVE IN EXINE1 gene product,<br />
a predicted membrane-associated prote<strong>in</strong> with limited similarity<br />
to animal <strong>in</strong>tegr<strong>in</strong>s, appears to <strong>in</strong>teract with the plasma<br />
membrane to nucleate sporopollen<strong>in</strong> deposition (Paxson-Sowders<br />
et al., 2001). An ex<strong>in</strong>e pattern mutant also has been<br />
described <strong>in</strong> Haplopappus gracilis, whose normal surface<br />
pattern <strong>of</strong> densely packed sp<strong>in</strong>es is reduced to a r<strong>and</strong>om scatter<br />
<strong>of</strong> sporopollen<strong>in</strong> deposits with few <strong>in</strong>terspersed sp<strong>in</strong>es (Jackson<br />
et al., 2000). Similarly, mutation <strong>in</strong> the Arabidopsis less adherent<br />
pollen1 gene yields mutant pollen gra<strong>in</strong>s with discont<strong>in</strong>uous<br />
ex<strong>in</strong>e walls composed only <strong>of</strong> globular sporopollen<strong>in</strong> deposits<br />
(Z<strong>in</strong>kl <strong>and</strong> Preuss, 2000). With the identification <strong>of</strong> additional<br />
mutants <strong>in</strong>volved <strong>in</strong> ex<strong>in</strong>e development, it will become possible<br />
to dist<strong>in</strong>guish those genes required for sporopollen<strong>in</strong> synthesis<br />
from those required to establish patterns. Pattern<strong>in</strong>g genes, <strong>in</strong><br />
particular, may be highly variable across species, either <strong>in</strong> their<br />
cod<strong>in</strong>g sequence or <strong>in</strong> their site <strong>and</strong> tim<strong>in</strong>g <strong>of</strong> expression.<br />
Because these genes are likely altered through evolution, ex<strong>in</strong>e<br />
patterns discovered by mutation <strong>and</strong> considered aberrant <strong>in</strong> one<br />
species may be normal <strong>in</strong> another species.<br />
Ex<strong>in</strong>e also varies <strong>in</strong> the number, distribution, <strong>and</strong> architecture<br />
<strong>of</strong> the apertures that <strong>in</strong>terrupt it (Figure 2). Apertures are diverse<br />
across taxa, with<strong>in</strong> families, with<strong>in</strong> species, <strong>and</strong> even with<strong>in</strong><br />
a s<strong>in</strong>gle plant (Mignot et al., 1994). <strong>Pollen</strong> from monocots<br />
characteristically has a s<strong>in</strong>gle aperture, a trait considered to be<br />
ancestral. Most dicot pollen gra<strong>in</strong>s have three apertures,<br />
although <strong>in</strong> both <strong>in</strong>stances, aperture numbers have <strong>in</strong>creased<br />
or decreased repeatedly <strong>and</strong> <strong>in</strong>dependently dur<strong>in</strong>g the course <strong>of</strong>





