

transcript of the animation - Harvard University
transcript of the animation - Harvard University
transcript of the animation - Harvard University

Create successful ePaper yourself
Turn your PDF publications into a flip-book with our unique Google optimized e-Paper software.
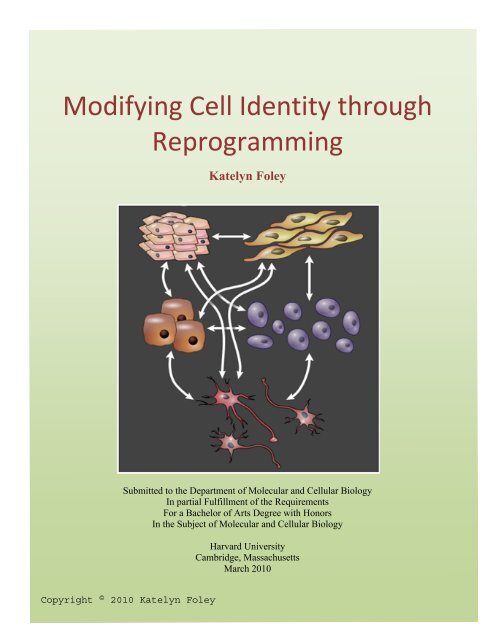
Modifying Cell Identity through <br />
Reprogramming <br />
Katelyn Foley<br />
Submitted to <strong>the</strong> Department <strong>of</strong> Molecular and Cellular Biology<br />
In partial Fulfillment <strong>of</strong> <strong>the</strong> Requirements<br />
For a Bachelor <strong>of</strong> Arts Degree with Honors<br />
In <strong>the</strong> Subject <strong>of</strong> Molecular and Cellular Biology<br />
<strong>Harvard</strong> <strong>University</strong><br />
Cambridge, Massachusetts<br />
March 2010
I. Development and Differentiation<br />
An embryo’s development into an adult organism is a unidirectional process associated<br />
with increasingly limited cell potential. The fusion <strong>of</strong> egg and sperm leads to <strong>the</strong> formation <strong>of</strong> a<br />
single-cell embryo called a zygote, which has <strong>the</strong> potential to form an entire organism, as well as<br />
extraembryonic tissues. The zygote and its progeny cells divide several times to form a<br />
blastocyst, or a pre-implantation embryo structured like a hollow ball <strong>of</strong> cells with an inner cell<br />
mass (ICM). Cells in <strong>the</strong> ICM are pluripotent, meaning that <strong>the</strong>y can give rise to all cell types in<br />
an organism (Fig. 1) (Surani et al. 2007).<br />
The process by which less specialized cells become increasingly specialized is called cell<br />
differentiation. During development, pluripotent inner cell mass cells give rise to multipotent<br />
stem cells that are more committed to specific lineages. These multipotent stem cells <strong>the</strong>n give<br />
rise to progenitor cells, which have limited potential and self-renewal. Finally, differentiated<br />
cells are generated from committed progenitors (Fig. 2). In some cases, tissue-specific stem cells<br />
are present in an adult organism in order to maintain and repair certain types <strong>of</strong> tissues. The<br />
process <strong>of</strong> cell differentiation can be mimicked in culture when pluripotent embryonic stem (ES)<br />
cells are isolated from <strong>the</strong> inner cell mass <strong>of</strong> a blastocyst.<br />
II. Epigenetic Changes Drive Differentiation<br />
As cells become more specialized, <strong>the</strong>y undergo epigenetic changes that reinforce a new<br />
cell identity. Epigenetic changes are non-genetic changes that affect <strong>the</strong> reading <strong>of</strong> DNA without<br />
altering <strong>the</strong> DNA sequence (Kouzarides, 2007). Covalent chemical modifications <strong>of</strong> DNA and<br />
histones influence gene expression by affecting a gene’s physical accessibility to <strong>transcript</strong>ion<br />
factors. Catalyzed by <strong>the</strong> DNA methyltransferase family <strong>of</strong> enzymes, <strong>the</strong> addition <strong>of</strong> methyl<br />
groups to certain cytosine bases hinders physical contact between a promoter and <strong>the</strong><br />
<strong>transcript</strong>ion factors required for <strong>transcript</strong>ion initiation (Fig. 3) (Attwood, 2002).<br />
Posttranslational histone modifications also play a role in gene expression by causing histones to<br />
loosen or tighten DNA and by recruiting proteins that initiate chromatin remodeling. At least<br />
eight different classes <strong>of</strong> modifications have been observed. For example, acetylation <strong>of</strong> histone<br />
amino termini can lead to activation <strong>of</strong> gene expression by decreasing <strong>the</strong> affinity <strong>of</strong> histones for<br />
DNA (Fig. 4) (Kouzarides 2007). Histone methylation is a versatile regulator that has been<br />
associated with gene activation and repression, depending on <strong>the</strong> locations <strong>of</strong> methylated lysine<br />
residues (Kouzarides 2007; Mikkelsen et al. 2007; Vakoc et al. 2005; Vakoc et al. 2006).<br />
Epigenetic marks represent <strong>transcript</strong>ional activity and gene expression for a genomic<br />
region (Aoto et al. 2006). Chromatin is found in two forms in interphase nuclei: <strong>transcript</strong>ionally<br />
active euchromatin and highly condensed, inactive heterochromatin. Both chromatin variants<br />
have a specific epigenetic signature based on <strong>the</strong> histone code, or <strong>the</strong> global chemical pr<strong>of</strong>ile <strong>of</strong><br />
histone amino termini (Jenuwein & Allis 2001). While euchromatin is characterized by<br />
hyperacetylation <strong>of</strong> histones H3 and H4 and by methylation <strong>of</strong> H3K4, H3K36, and H3K79,<br />
heterochromatin displays low acetylation and methylation <strong>of</strong> H3K9, H3K27, and H4K20 (Aoto<br />
et al. 2006; Kouzarides 2007).<br />
The differentiation <strong>of</strong> stem cells is driven by epigenetic changes that reinforce a new<br />
<strong>transcript</strong>ional program. Previous research indicates that stem cells are characterized by global<br />
acetylation <strong>of</strong> histone H3 (Kimura et al. 2004; Lee et al. 2004) and by global hypomethylation<br />
(Jackson et al. 2004; Zvetkova et al. 2005), suggesting that chromatin might be in a more<br />
2
permissive state in undifferentiated cells. In addition, a deficiency in DNA methyltransferase<br />
greatly hinders stem cell differentiation, resulting in abnormal development and embryonic<br />
lethality (Li et al. 1992; Okano et al. 1999). Aligned with <strong>the</strong>se findings are data showing that ES<br />
cells express 40 to 60% <strong>of</strong> <strong>the</strong> genes in a genome, whereas only 10 to 20% are expressed in<br />
differentiated cells (Abeyta et al. 2004). From this perspective, differentiation may involve <strong>the</strong><br />
conversion <strong>of</strong> active euchromatin into stably silenced heterochromatin (Gasser 2002).<br />
On a gene-specific level, ES-cell-specific genes are associated with activating epigenetic<br />
marks and <strong>transcript</strong>ion machinery in ES cells. Oct-3/4 and Nanog, which have been implicated<br />
in <strong>the</strong> maintenance <strong>of</strong> stem cell characteristics, are unmethylated and associated with histone<br />
acetylation in ES cells (Delcuve et al. 2009). Although RNA Polymerase-II binding to <strong>the</strong> Oct-<br />
3/4 promoter has been observed in ES cells, it is lost in neural precursor cells and post-mitotic<br />
neurons (Aoto et al. 2006). These findings suggest that differentiation status is based on an<br />
epigenetic signature determined by DNA methylation patterns and histone modifications.<br />
III. Nuclear Reprogramming<br />
The 220 cell types in <strong>the</strong> mammalian body are established and maintained by specific<br />
patterns <strong>of</strong> gene expression. Although cells do not naturally express genes that are specific to<br />
unrelated cell types, experimental procedures have been identified that allow dramatic switches<br />
in gene expression and cell identity (Fig. 5). The cloning <strong>of</strong> amphibians and mammals from<br />
differentiated cells (Briggs & King 1957; Gurdon 1962; Wilmut et al. 1997) has shown that cell<br />
differentiation is caused by reversible epigenetic changes (Hochedlinger & Jaenisch 2006). With<br />
<strong>the</strong> exception <strong>of</strong> antibody-producing cells, all cells in <strong>the</strong> body are genetically identical, even<br />
though <strong>the</strong>ir distinct morphology and behavior is determined by a specific pattern <strong>of</strong> gene<br />
expression (Fig. 6). Nuclear reprogramming describes a change in <strong>the</strong> nuclear gene expression <strong>of</strong><br />
one cell type to that <strong>of</strong> an unrelated cell type or embryo (Fig. 7) (Gurdon & Melton 2008). The<br />
genetic equivalence across cell types makes nuclear reprogramming feasible.<br />
At <strong>the</strong> molecular level, nuclear reprogramming is driven by epigenetic changes that alter<br />
gene expression and cell identity. For example, <strong>the</strong> conversion <strong>of</strong> a differentiated cell into a stem<br />
cell is associated with histone modifications that reverse cell differentiation. As somatic cells<br />
return to a pluripotent state, lysine residues <strong>of</strong> histones H3 and H4 become hyperacetylated, and<br />
lysine 4 <strong>of</strong> H3 becomes hypermethylated, corresponding to <strong>the</strong> permissive state <strong>of</strong> chromatin in<br />
ES cells (Kimura et al. 2004). O<strong>the</strong>r work shows that this transition also involves changes in<br />
histone and DNA methylation patterns (Maherali et al. 2007; Okita et al. 2007; Wernig et al.<br />
2007). Complete epigenetic reprogramming <strong>of</strong> cancer cells underscores <strong>the</strong> important role that<br />
epigenetic changes play in generating cell heterogeneity (Hochedlinger et al. 2004). Taken<br />
toge<strong>the</strong>r, <strong>the</strong>se data support <strong>the</strong> epigenetic basis <strong>of</strong> cell identity and reveal a high level <strong>of</strong><br />
epigenetic plasticity.<br />
There are currently three methods for achieving reprogramming: somatic cell nuclear<br />
transfer, cell fusion, and direct reprogramming.<br />
IV. Significance<br />
Nuclear reprogramming not only presents a framework for investigating <strong>the</strong> mechanisms<br />
that govern epigenetic memory and <strong>the</strong> maintenance <strong>of</strong> cell identity but it also provides a model<br />
for <strong>the</strong> generation <strong>of</strong> patient-specific cells for transplantation (Gurdon & Melton 2008). Somatic<br />
3
cells from a patient can be reprogrammed into immune-compatible cells that behave like<br />
pluripotent stem cells. These cells can be expanded and induced to differentiate into a needed<br />
cell type in vitro, providing patient-specific cells for replacement <strong>of</strong> diseased or damaged tissues<br />
(Fig. 8). A needed cell type may also be directly generated from patient cells without <strong>the</strong> need<br />
for intermediate pluripotent cells (Fig. 9). In addition, nuclear reprogramming enables <strong>the</strong><br />
generation <strong>of</strong> diseased cell lines, allowing observation <strong>of</strong> disease progression and drug screening.<br />
V. Somatic Cell Nuclear Transfer<br />
Early experiments in amphibians showed that cells across <strong>the</strong> differentiation spectrum<br />
can be reprogrammed when <strong>the</strong>ir nuclei are injected into enucleated eggs (Briggs & King 1957;<br />
Gurdon 1962). In somatic cell nuclear transfer (SCNT), <strong>the</strong> nucleus <strong>of</strong> a somatic cell is injected<br />
into an egg that has had its own chromatin removed. Although some <strong>of</strong> <strong>the</strong> somatic proteins are<br />
transferred with <strong>the</strong> nucleus, <strong>the</strong> large volume <strong>of</strong> egg cytoplasm dilutes <strong>the</strong> somatic<br />
<strong>transcript</strong>ional program, allowing <strong>the</strong> embryonic <strong>transcript</strong>ional program to dominate (Fig. 10)<br />
(Gurdon & Melton 2008). The reprogramming <strong>of</strong> somatic chromatin is associated with<br />
epigenetic changes that allow reactivation <strong>of</strong> early embryonic genes, such as Oct-3/4 (Simonsson<br />
& Gurdon 2004).<br />
In mammals, metaphase II oocytes (Wilmut et al. 1997) and mitotic zygotes (Egli et al.<br />
2007) have been shown to reprogram somatic nuclei. Somatic cell nuclear transfer can lead to <strong>the</strong><br />
development <strong>of</strong> a normal blastocyst, from which embryonic stem cells can be derived for<br />
transplantation <strong>the</strong>rapies. This process is termed <strong>the</strong>rapeutic cloning. In some cases, SCNT can<br />
support <strong>the</strong> full-term development <strong>of</strong> an adult organism. Cloned from an adult mammary gland<br />
cell, Dolly <strong>the</strong> sheep is a famous example <strong>of</strong> reproductive cloning (Wilmut et al. 2002). At this<br />
point in time, a number <strong>of</strong> ethical concerns and technical limitations surround human SCNT (see<br />
Jaenisch 2004 for review).<br />
Nuclear transfer experiments have yielded valuable information about <strong>the</strong> nature <strong>of</strong> <strong>the</strong><br />
epigenetic changes that drive differentiation. Thirty percent <strong>of</strong> nuclear transfer experiments give<br />
rise to a normal adult when <strong>the</strong> donor nucleus is embryonic, but only 1 to 2% are successful<br />
when <strong>the</strong> donor nucleus is from a differentiated cell (Tecirlioglu et al. 2006). However, even<br />
terminally differentiated cells, such as certain post-mitotic neurons (Eggan et al. 2004; Li et al.<br />
2004) and mature B and T cells (Hochedlinger & Jaenisch 2002), have been shown to support<br />
<strong>the</strong> full-term development <strong>of</strong> mammalian clones. These data indicate that even <strong>the</strong> most<br />
differentiated states are reversible but that <strong>the</strong> epigenetic modifications that characterize highly<br />
specialized cells are more difficult to undo.<br />
VI. Cell Fusion<br />
Fusion <strong>of</strong> unrelated cell types has been used to investigate cell plasticity for decades<br />
(Blau & Blakely 1999). When two cells are fused, a heterokaryon with two distinct nuclei is<br />
formed. In some cases, nuclei from fused partner cells merge, leading to a stable hybrid cell that<br />
is directed by <strong>the</strong> <strong>transcript</strong>ional program <strong>of</strong> one <strong>of</strong> <strong>the</strong> partner cells (Fig. 11). Previous work has<br />
shown that a partner cell’s ability to impose its pattern <strong>of</strong> gene expression on a heterokaryon or<br />
hybrid cell is dependent on its cell division activity and size (Gurdon & Melton 2008).<br />
Unspecialized cells, which generally have active cell cycles, have been implicated in<br />
<strong>transcript</strong>ional dominance after fusion with specialized cells (Hochedlinger & Jaenisch 2006).<br />
4
ES cells can reprogram somatic nuclei when fused with somatic cells, in both <strong>the</strong> mouse<br />
(Tada et al. 2001) and human (Cowan et al. 2005) models. ES-somatic fusion can lead to <strong>the</strong><br />
demethylation and reactivation <strong>of</strong> ES-cell-specific genes from somatic chromosomes.<br />
Reprogrammed cells that display embryonic stem cell morphology, growth, and gene expression<br />
have been isolated by using a genetic marker that indicates reactivation <strong>of</strong> somatic Oct-3/4 (Fig.<br />
12). In addition, ES-somatic hybrids can form teratomas that express genes from all three germ<br />
layers, a hallmark <strong>of</strong> pluripotency (Cowan et al. 2005; Tada et al. 2003). However, it is important<br />
to note that global gene expression analysis has revealed differences between <strong>the</strong> ES cell and ESfibroblast<br />
hybrid <strong>transcript</strong>omes, although major regulators <strong>of</strong> pluripotency are similarly<br />
expressed (Ambrosi et al. 2007).<br />
The fusion <strong>of</strong> differentiated cell types has also suggested that <strong>the</strong> <strong>transcript</strong>ional program<br />
<strong>of</strong> one partner cell can dominate a heterokaryon or hybrid. When hepatocytes are fused with<br />
myotubes, non-muscle nuclei in <strong>the</strong> resulting heterokaryons express muscle genes, indicating<br />
that <strong>the</strong> muscle phenotype is dominant (Fig. 13) (Pomerantz et al. 2009). Similarly, <strong>the</strong> fusion <strong>of</strong><br />
B cells and myotubes causes B cell nuclei to become physically structured like muscle nuclei and<br />
to express muscle-specific genes in a sequence that resembles muscle development. The<br />
establishment <strong>of</strong> muscle nuclear gene expression is associated with <strong>the</strong> silencing <strong>of</strong> lymphocyte<br />
genes, suggesting loss <strong>of</strong> B cell epigenetic memory (Terranova et al. 2006). In both embryonicsomatic<br />
and somatic-somatic fusions, it will be necessary to separate chromatin from each<br />
partner cell in order to understand whe<strong>the</strong>r transient <strong>transcript</strong>ional domination or true<br />
reprogramming has occurred.<br />
VII. Direct Reprogramming<br />
Direct reprogramming is achieved by ectopic expression <strong>of</strong> specific genes that lead to a<br />
switch in cell identity. Although <strong>the</strong> exact mechanisms <strong>of</strong> reprogramming are still being<br />
elucidated, this switch is associated with epigenetic changes, stepwise silencing and activation <strong>of</strong><br />
endogenous genes, and sustained reprogramming after ectopically expressed genes are silenced<br />
(Jaenisch & Young 2008). Direct reprogramming encompasses pluripotent reprogramming, or<br />
<strong>the</strong> generation <strong>of</strong> induced pluripotent stem (iPS) cells from more specialized cells, and lineage<br />
reprogramming, or <strong>the</strong> conversion <strong>of</strong> cells from one lineage to ano<strong>the</strong>r.<br />
The first direct reprogramming experiments showed that forced expression <strong>of</strong> <strong>the</strong> muscle<br />
<strong>transcript</strong>ion factor MyoD in a number <strong>of</strong> cell types, including differentiated hepatocytes and<br />
adipocytes, could lead to <strong>the</strong> activation <strong>of</strong> muscle-specific genes (Fig. 14) (Weintraub et al.<br />
1989). The most famous breakthrough in <strong>the</strong> field came when Takahashi and Yamanaka showed<br />
that retroviral transfection <strong>of</strong> Oct-3/4, Sox2, Klf4, and c-Myc can reprogram murine fibroblasts<br />
into iPS cells (2006). The same factors (Takahashi et al. 2007), as well as a different quartet <strong>of</strong><br />
Oct-3/4, Sox2, Nanog, and Lin28 (Yu et al. 2007), can reprogram human fibroblasts (Fig. 15). In<br />
both cases, endogenous ES-cell-specific genes are reactivated and remain active even after<br />
viruses have been silenced (Fig. 16). Follow-up work has yielded iPS cells that are more ES-like<br />
(Maherali et al. 2007; Okita et al. 2007; Wernig et al. 2007) and more clinically applicable due to<br />
<strong>the</strong> use <strong>of</strong> nonintegrating adenoviruses (Okita et al. 2008; Stadtfeld et al. 2008b). iPS cells can be<br />
generated from a wide range <strong>of</strong> cell types, including terminally differentiated B cells (Hanna et<br />
al. 2007) and pancreatic β cells (Stadtfeld et al. 2008a), and <strong>the</strong>n induced to differentiate into<br />
cells from all three germ layers. The broad applications <strong>of</strong> pluripotent reprogramming support its<br />
5
<strong>the</strong>rapeutic potential. However, it is important to note that genome-wide expression pr<strong>of</strong>iles<br />
suggest that iPS cells are not identical to wild-type ES cells (Mikkelsen et al. 2008).<br />
Recent lineage reprogramming experiments have shown that differentiated cells and<br />
specialized progenitors can take on <strong>the</strong> nuclear gene expression patterns <strong>of</strong> o<strong>the</strong>r lineages,<br />
suggesting that intermediate pluripotent cells might not be required for some switches in cell<br />
identity. Adenoviral infection with Ngn3, Pdx1, and Mafa can lead to in vivo reprogramming <strong>of</strong><br />
pancreatic exocrine cells into insulin-producing β-cells (Zhou et al. 2008). In addition, in vitro<br />
studies have shown that forced expression <strong>of</strong> C/EBPα or C/EBPβ can cause B cells to convert<br />
into phagocytic macrophages with 100% efficiency (Bussmann et al. 2009). The use <strong>of</strong> similar<br />
strategies to successfully convert more distantly related cell types will require a detailed<br />
understanding <strong>of</strong> cell lineages and <strong>the</strong> <strong>transcript</strong>ion factors that establish and maintain specific<br />
cell identities.<br />
VIII. Mechanisms behind Nuclear Reprogramming<br />
The specific mechanisms behind somatic cell nuclear transfer, cell fusion, and direct<br />
reprogramming seem to be different in each scenario. Both SCNT and cell fusion involve a<br />
competition between two <strong>transcript</strong>ional programs, but <strong>the</strong> inherent reprogramming capabilities<br />
and large size <strong>of</strong> eggs increase <strong>the</strong> efficiency <strong>of</strong> SCNT. Unlike SCNT and cell fusion, which<br />
introduce nuclei to a complex cocktail <strong>of</strong> <strong>transcript</strong>ion factors from ano<strong>the</strong>r cell, direct<br />
reprogramming is driven by <strong>the</strong> forced expression <strong>of</strong> select <strong>transcript</strong>ion factors. An<br />
investigation <strong>of</strong> <strong>the</strong> timing <strong>of</strong> expression <strong>of</strong> iPS cell factors Nanog, Klf4, Myc, Lin28, and Oct4,<br />
showed that only Lin28 and Oct4 are expressed in <strong>the</strong> early embryo. Nanog, Klf4, and Myc,<br />
which have been implicated in iPS generation, are expressed at later stages, suggesting that <strong>the</strong>y<br />
do not mediate reprogramming in SCNT (Esikov et al. 2004; Esikov et al. 2006). Fur<strong>the</strong>rmore,<br />
<strong>the</strong> addition <strong>of</strong> Nanog to <strong>the</strong> Oct-3/4/Sox2/Klf4/c-Myc quartet <strong>of</strong> iPS cell factors has not been<br />
shown to increase <strong>the</strong> efficiency <strong>of</strong> pluripotent reprogramming (Silva et al. 2008). In contrast, <strong>the</strong><br />
fusion <strong>of</strong> Nanog-overexpressing ES cells with fibroblasts or thymocytes can significantly<br />
increase <strong>the</strong> number <strong>of</strong> ES-like hybrid cells (Silva et al. 2006).<br />
Despite <strong>the</strong>se differences, <strong>the</strong>re are commonalities between <strong>the</strong> reprogramming processes<br />
observed for each method. Egg cytoplasm, whole cells, and select <strong>transcript</strong>ion factors can bring<br />
about epigenetic changes when introduced to nuclei from unrelated cell types. In SCNT, <strong>the</strong><br />
treatment <strong>of</strong> oocytes with a histone deacetylase inhibitor leads to more faithful reprogramming<br />
and a 100% increase in <strong>the</strong> number <strong>of</strong> normal blastocysts (Rybouchkin et al. 2006). Compared to<br />
non-treated oocytes, treated oocytes display more ES-like levels <strong>of</strong> chromatin structure- and<br />
DNA methylation-related genes (Li et al. 2008). Cell fusion has shown that histone deacetylases<br />
have a cell-type-specific effect on reprogramming. While HDACs are required for erasure <strong>of</strong><br />
lymphocyte identity in B cell-myotube heterokaryons (Terranova et al. 2006), <strong>the</strong>y promote<br />
muscle gene expression in hepatocyte-myotube heterokaryons (Pomerantz et al. 2009). In direct<br />
reprogramming, histone deacetylase inhibitors (Huangfu et al. 2008a; Huangfu et al. 2008b), and<br />
demethylation-promoting agents (Mikkelsen et al. 2008) can increase <strong>the</strong> efficiency <strong>of</strong> iPS cell<br />
generation. Although epigenetic changes are a common feature <strong>of</strong> reprogramming, <strong>the</strong>y vary<br />
according to <strong>the</strong> differentiation states <strong>of</strong> <strong>the</strong> nucleus and incoming <strong>transcript</strong>ional program.<br />
In addition to epigenetic changes, SCNT, cell fusion, and direct reprogramming involve a<br />
competition between <strong>transcript</strong>ional programs. The introduction <strong>of</strong> a cellular cocktail or select<br />
<strong>transcript</strong>ion factors challenges a cell’s ability to self-reprogram with its own set <strong>of</strong> factors<br />
6
(Gurdon & Melton 2008). During <strong>the</strong> process <strong>of</strong> reprogramming to a pluripotent state, egg<br />
cytoplasm, ES cells, and defined ES-cell-associated <strong>transcript</strong>ion factors override a somatic<br />
<strong>transcript</strong>ional program by activating <strong>the</strong> Oct-3/4/Nanog/Sox2 autoregulatory loop. Oct-3/4,<br />
Nanog, and Sox2 co-occupy a significant number <strong>of</strong> <strong>the</strong>ir target genes, including <strong>the</strong>ir own genes<br />
(Boyer et al. 2005). Although special reprogramming powers have been attributed to eggs, recent<br />
research suggests that DNA methylation patterns in sperm and ES cells are surprisingly similar,<br />
except for methylation <strong>of</strong> Oct-3/4 and Nanog in sperm (Farthing et al. 2008; Imamura et al.<br />
2006). This indicates that post-fertilization reprogramming events might not be as extensive as<br />
previously thought and are instead concerned with demethylation <strong>of</strong> key pluripotency regulators<br />
(Albert & Peters 2009). Similarly, Oct-3/4 is required for reactivation <strong>of</strong> Oct-3/4, Nanog, and<br />
Sox2 in lymphocyte nuclei when ES cells are fused with B cells. Heterokaryon-based<br />
reprogramming occurs after nuclear translocation <strong>of</strong> ES-derived Oct-3/4, suggesting that Oct-3/4<br />
directly activates ES core <strong>transcript</strong>ional circuitry in lymphocyte nuclei (Pereira et al. 2008).<br />
Although factors o<strong>the</strong>r than Oct-3/4, Nanog, and Sox2 have been implicated in iPS cell<br />
generation, treatment <strong>of</strong> transfected cells with a histone deacetylase inhibitor results in<br />
pluripotent reprogramming with only Oct-3/4 and Sox2 (Huangfu et al. 2008b). It has been<br />
suggested that Nanog is dispensable for pluripotent reprogramming because it is sufficiently<br />
activated by Oct-3/4 (Jaenisch & Young 2008). Taken toge<strong>the</strong>r, <strong>the</strong>se data indicate that onset <strong>of</strong><br />
ES cell identity in a somatic nucleus is associated with activation <strong>of</strong> <strong>the</strong> Oct-3/4/Nanog/Sox2<br />
autoregulatory loop.<br />
It is important to note that <strong>the</strong> temporal relationship between epigenetic changes and<br />
activation <strong>of</strong> a new <strong>transcript</strong>ional program may be different for each method. Epigenetic<br />
changes can be directed by <strong>the</strong> regulatory activity <strong>of</strong> <strong>transcript</strong>ion factors (Lu et al. 2009).<br />
However, in reprogramming, chromatin reconfiguration may be necessary for syn<strong>the</strong>sis <strong>of</strong><br />
<strong>transcript</strong>ion factors that mediate additional epigenetic changes and activate cell identity-defining<br />
genes. Reprogramming by SCNT, including demethylation <strong>of</strong> <strong>the</strong> Oct-3/4 promoter, can occur in<br />
<strong>the</strong> absence <strong>of</strong> DNA replication and RNA/protein syn<strong>the</strong>sis, suggesting that existing maternal<br />
factors induce epigenetic changes (Simonsson & Gurdon 2004). Cell fusion-mediated<br />
reprogramming, which can take place during <strong>the</strong> heterokaryon stage, has been associated with<br />
diffusible trans-acting regulators that initiate expression <strong>of</strong> regulatory genes in <strong>the</strong> reprogrammed<br />
nucleus (Blau & Blakely 1999). Although lineage reprogramming can be mediated by one<br />
lineage-specific <strong>transcript</strong>ion factor, pluripotent reprogramming requires ES-cell-associated<br />
<strong>transcript</strong>ion factors and genes or chemicals implicated in chromatin modification. This suggests<br />
that conversions between more distantly related cell types involve a dynamic interaction between<br />
epigenetic changes and gene expression in <strong>the</strong> case <strong>of</strong> direct reprogramming. These differences<br />
might account for some <strong>of</strong> <strong>the</strong> variance in duration and efficiency that has been observed for<br />
different reprogramming methods.<br />
IX. Nuclear Reprogramming Exploits Inherent Cell Plasticity<br />
Although cell differentiation is normally a unidirectional, irreversible process, organisms<br />
rely on some level <strong>of</strong> cell plasticity for environmental responses. While <strong>the</strong> controversial<br />
transdifferentiation events reported by a number <strong>of</strong> studies are most likely <strong>the</strong> result <strong>of</strong> cell<br />
fusion or stress-induced cellular look-alikes that express only limited markers <strong>of</strong> ano<strong>the</strong>r cell<br />
type (Jaenisch & Young 2008), certain evidence indicates that some cell types might not follow<br />
traditional lineage paths (Slack 2008). A number <strong>of</strong> mysteries remain in <strong>the</strong> hematopoietic<br />
7
system, in which tissue-specific stem cells that are not present in <strong>the</strong> early embryo are found later<br />
in development (Orkin & Zon 2008). In addition, recent work has shown that a small percentage<br />
<strong>of</strong> ES-derived endo<strong>the</strong>lial cells loosen <strong>the</strong>ir tight junctions, ga<strong>the</strong>r toge<strong>the</strong>r, and express erythroid<br />
and monocytic hematopoietic antigens in culture (Eilken et al. 2009). In this context, <strong>the</strong> efficient<br />
switch from B cell to macrophage identity in vitro could imply a mechanism for generating<br />
macrophages in vivo. Specific pathogens could induce this reprogramming event by triggering<br />
expression <strong>of</strong> C/EBPα or C/EBPβ when <strong>the</strong>y infect B cells (Xie et al. 2004). In order to identify<br />
true transdifferentiation events in vivo, it will be necessary to mark homogenous cell populations<br />
with a label that rigorously defines cells lineages.<br />
Epigenetic status and <strong>transcript</strong>ion factor activity also reveal natural cell plasticity.<br />
Consistent with <strong>the</strong>ir pluripotency, embryonic stem cells have epigenetic marks that allow <strong>the</strong>m<br />
to efficiently commit to cell fates. In ES cells, developmentally significant <strong>transcript</strong>ion factors<br />
are associated with bivalent domains that contain repressive and activating histone modifications<br />
(Bernstein et al. 2006). These domains, which are mostly monovalent in differentiated cells, may<br />
demonstrate readiness to take on a specific cell fate. On a <strong>transcript</strong>ional level, <strong>the</strong> activity <strong>of</strong><br />
<strong>transcript</strong>ion-associated proteins is paused during cell division. Displacement <strong>of</strong> <strong>transcript</strong>ion<br />
factors, chromatin remodeling complexes, and RNA polymerases from mitotic chromosomes has<br />
been observed for a range <strong>of</strong> cell types (Delcuve et al. 2008). Asymmetric segregation <strong>of</strong> <strong>the</strong>se<br />
displaced factors can allow daughter cells to inherit different <strong>transcript</strong>ional programs from a<br />
parent cell. Thus, mitosis represents a weak link in a cell’s ability to preserve epigenetic<br />
memory, temporarily causing vulnerability to <strong>transcript</strong>ional changes. In interphase, ES cells are<br />
also amenable to large-scale <strong>transcript</strong>ional changes because <strong>of</strong> lower protein binding constants<br />
than in differentiated cells. Recent work has shown that ES cell proteins have a looser, more<br />
transient association with DNA, while proteins in differentiated cell types can be associated with<br />
DNA for several minutes (Meshorer et al. 2006; Phair et al. 2004).<br />
Taken toge<strong>the</strong>r, <strong>the</strong>se data suggest that reprogramming may be dependent on a<br />
combination <strong>of</strong> epigenetic and <strong>transcript</strong>ional plasticity. An understanding <strong>of</strong> <strong>the</strong> characteristics<br />
that support cell plasticity may elucidate new strategies for efficient reprogramming <strong>of</strong> highly<br />
differentiated cells.<br />
X. Clinical Progress in <strong>the</strong> Field <strong>of</strong> Reprogramming<br />
Nuclear reprogramming has yielded immune-compatible cells for transplantation<br />
<strong>the</strong>rapies in <strong>the</strong> rodent model, as well as human iPS cell lines from individuals with a variety <strong>of</strong><br />
diseases. The isolation <strong>of</strong> ntES cells, or embryonic stem cells that are derived from nuclear<br />
transfer blastocysts, has enabled fur<strong>the</strong>r investigation <strong>of</strong> <strong>the</strong>rapeutic cloning (Wakayama et al.<br />
2001). One study showed that ntES cells with <strong>the</strong> genotype <strong>of</strong> immune-deficient mice could be<br />
repaired using homologous recombination and <strong>the</strong>n induced to differentiate into hematopoietic<br />
stem cells for transplantation. This treatment led to partial rescue <strong>of</strong> immune function in<br />
immune-deficient mice (Rideout et al. 2002). In ano<strong>the</strong>r study, ntES-cell-derived dopaminergic<br />
neurons were transplanted into mice with Parkinson’s disease, resulting in alleviation <strong>of</strong><br />
behaviors associated with <strong>the</strong> disease (Barberi et al. 2003).<br />
Pluripotent reprogramming has also been used to generate immune-compatible cells for<br />
transplantation. Recent work in <strong>the</strong> rat and mouse models has shown that iPS-based cell<br />
transplantation <strong>the</strong>rapies can improve <strong>the</strong> symptoms <strong>of</strong> Parkinson’s disease (Wernig et al. 2008)<br />
and sickle cell anemia (Hanna et al. 2007). However, it is important to note that some <strong>of</strong> <strong>the</strong><br />
8
Parkinson’s diseased animals developed tumors after treatment, possibly due to <strong>the</strong> presence <strong>of</strong><br />
undifferentiated cells in <strong>the</strong> graft. In addition, patient-specific iPS cell lines have been generated<br />
from humans with a variety <strong>of</strong> diseases, including diabetes, Parkinson’s disease, and<br />
Amyotrophic Lateral Sclerosis (ALS) (Dimos et al. 2008; Park et al. 2008). In <strong>the</strong> future, <strong>the</strong>se<br />
cell lines could facilitate disease modeling and drug screening.<br />
Currently, lineage reprogramming is <strong>the</strong> only method <strong>of</strong> reprogramming that has potential<br />
for in vivo cell conversion, since pluripotent stem cells do not have direct <strong>the</strong>rapeutic<br />
applications and could not be reliably differentiated in vivo without risk <strong>of</strong> tumorigenesis.<br />
Lineage reprogramming can be used to convert an existing specialized cell into a needed cell<br />
type in <strong>the</strong> body. The recent study in which pancreatic exocrine cells were reprogrammed into<br />
insulin-producing β cells in vivo showed that <strong>the</strong>se cells could improve hyperglycemia by<br />
secreting insulin (Zhou et al. 2008).<br />
To date, immune-incompatibility and high DNA ploidy <strong>of</strong> hybrid cells limit <strong>the</strong> clinical<br />
applications <strong>of</strong> cell fusion. Tetraploidy is thought to promote genetic instability that can lead to<br />
aneuploidy and tumorigenesis in vivo (Ganem et al. 2007). Although ES cell chromosomes have<br />
been successfully targeted and eliminated from ES-somatic hybrids using a chromosome<br />
elimination cassette (Matsumura et al. 2007), this technique could lead to global chromosomal<br />
instability in hybrids (Jaenisch & Young 2008). In order to generate clinically applicable patientspecific<br />
cells using cell fusion, it will be necessary to efficiently and completely remove ES cell<br />
chromatin before or after fusion.<br />
XI. Conclusion<br />
Nuclear reprogramming takes advantage <strong>of</strong> inherent epigenetic and <strong>transcript</strong>ional<br />
plasticity in a range <strong>of</strong> cell types. An understanding <strong>of</strong> <strong>the</strong> molecular mechanisms that support<br />
this plasticity can support <strong>the</strong> development <strong>of</strong> reprogramming strategies that exploit natural cell<br />
behaviors. One crucial question that remains is whe<strong>the</strong>r <strong>the</strong> introduction <strong>of</strong> a <strong>transcript</strong>ional<br />
program to permissive chromatin can trigger appropriate epigenetic changes. A future direction<br />
could be to elicit epigenetic changes by exposure to small molecules before introducing a new<br />
<strong>transcript</strong>ional program to a nucleus. In <strong>the</strong> case <strong>of</strong> cell fusion, it would be ideal to transfer<br />
mitotic chromatin from a patient cell to a mitotic cytoplast derived from a needed cell type,<br />
although this remains technically challenging. This strategy would minimize competition<br />
between <strong>transcript</strong>ional programs by exploiting mitotic detachment from <strong>transcript</strong>ional identity.<br />
However, <strong>the</strong> dramatic nature <strong>of</strong> certain cell conversions might make it necessary for a cell to<br />
return to <strong>the</strong> branch point between two lineages before being induced to differentiate into a<br />
needed cell type. To date, experimental methods can recapitulate cell differentiation for a range<br />
<strong>of</strong> pathways, including hematopoietic, neural, and mesenchymal.<br />
Even though nuclear reprogramming has demonstrated clinical value in <strong>the</strong> mouse model,<br />
it is important to recognize that reprogrammed cells are not exactly <strong>the</strong> same as <strong>the</strong>ir wild-type<br />
counterparts. Reprogrammed cells seem to stably take on <strong>the</strong> nuclear gene expression pattern <strong>of</strong><br />
a specific cell type. However, it remains to be seen whe<strong>the</strong>r we can generate reprogrammed cells<br />
with <strong>the</strong> epigenetic signature, global gene expression, and cell architecture <strong>of</strong> corresponding cells<br />
in <strong>the</strong> body. Ultimately, it may be that reprogramming will be most successful when different<br />
reprogramming methods are applied to specific cell type conversions. By understanding <strong>the</strong><br />
molecular mechanisms behind reprogramming, we can develop more efficient, direct strategies<br />
for generating patient-specific cells for transplantation (Fig. 22).<br />
9
References:<br />
1. Abeyta, M.J. et al. Unique gene expression signatures <strong>of</strong> independently-derived human<br />
embryonic stem cell lines. Hum. Mol. Genet. 13, 601-608 (2004).<br />
2. Albert, M. & Peters, A. Genetic and epigenetic control <strong>of</strong> early mouse development.<br />
Curr. Opin. Genet. Dev. 19, 113-121 (2009).<br />
3. Ambrosi, D.J. et al. Genome-wide reprogramming in hybrids <strong>of</strong> somatic cells and<br />
embryonic stem cells. Stem Cells 25, 1104-1113 (2007).<br />
4. Aoto, T., Saitoh, N., Ichimura, T., Niwa, H. & Nakao, M. Nuclear and chromatin<br />
reorganization in <strong>the</strong> MHC-Oct3/4 locus at developmental phases <strong>of</strong> embryonic stem cell<br />
differentiation. Dev. Biol. 298, 354-367 (2006).<br />
5. Attwood, J.T., Yung, R.L. & Richardson, B.C. DNA methylation and <strong>the</strong> regulation <strong>of</strong><br />
<strong>transcript</strong>ion. Cell Mol. Life Sci. 59, 241-257 (2002).<br />
6. Barberi, T. et al. Neural subtype specification <strong>of</strong> fertilization and nuclear transfer<br />
embryonic stem cells and application in parkinsonian mice. Nat. Biotechnol. 21, 1200-<br />
1207 (2003).<br />
7. Bernstein, B.E. et al. A Bivalent Chromatin Structure Marks Key Developmental Genes<br />
in Embryonic Stem Cells. Cell 125, 315-326 (2006).<br />
8. Blau, H.M. & Blakely, B.T. Plasticity <strong>of</strong> cell fate: Insights from heterokaryons. Cell Dev.<br />
Biol. 10, 267-272 (1999).<br />
9. Boyer, L.A. et al. Core <strong>transcript</strong>ional regulatory circuitry in human embryonic stem<br />
cells. Cell 122, 947-956 (2005).<br />
10. Briggs, R. & King, T.J. Changes in <strong>the</strong> nuclei <strong>of</strong> differentiating endoderm cells as<br />
revealed by nuclear transplantation. J. Morphol. 100, 269-311 (1957).<br />
11. Bussmann, L.H. et al. A robust and highly efficient immune cell reprogramming system.<br />
Cell Stem Cell 5, 554-566 (2009).<br />
12. Cowan, C.A., Atienza, J., Melton, D.A. & Eggan, K. Nuclear reprogramming <strong>of</strong> somatic<br />
cells after fusion with human embryonic stem cells. Science 309, 1369-1373 (2005).<br />
13. Delcuve, G., He, S. & Davie, J.R. Mitotic partitioning <strong>of</strong> <strong>transcript</strong>ion factors. J. Cell.<br />
Biochem. 105, 1-8 (2008).<br />
14. Delcuve, G.P., Rastegar, M. & Davie, J. Epigenetic Control. J. Cell. Physiol. 219, 243-<br />
250 (2009).<br />
10
15. Dimos, J.T. et al. Induced pluripotent stem cells generated from patients with ALS can be<br />
differentiated into motor neurons. Science 321, 1218-1221 (2008).<br />
16. Do, J.T. & Schöler, H.R. Nuclei <strong>of</strong> embryonic stem cells reprogram somatic cells. Stem<br />
Cells 22, 941-949 (2004).<br />
17. Eggan, K. et al. Mice cloned from olfactory sensory neurons. Nature 428, 44-49 (2004).<br />
18. Eilken, H.M., Nishikawa, S. & Schroeder, T. Continuous single-cell imaging <strong>of</strong> blood<br />
generation from haemogenic endo<strong>the</strong>lium. Nature 457, 896-900 (2009).<br />
19. Esikov, A.V. et al. Systems biology <strong>of</strong> <strong>the</strong> 2-cell mouse embryo. Cytogenet. Gen. Res.<br />
105, 240-250 (2004).<br />
20. Esikov, A.V. et al. Cracking <strong>the</strong> egg: Molecular dynamics and evolutionary aspects <strong>of</strong> <strong>the</strong><br />
transition from <strong>the</strong> fully grown oocyte to embryo. Genes Dev. 20, 2713-2727 (2006).<br />
21. Farthing, C.R. et al. Global mapping <strong>of</strong> DNA methylation in mouse promoters reveals<br />
epigenetic reprogramming <strong>of</strong> pluripotency genes. PLoS Genet. 4, e1000116 (2008).<br />
22. Ganem, N.J., Storchova, Z. & Pellman, D. Tetraploidy, aneuploidy and cancer. Curr.<br />
Opin. Genet. Dev. 17, 157-162 (2007).<br />
23. Gasser, S.M. Visualizing chromatin dynamics in interphase nuclei. Science 296, 1412-<br />
1416 (2002).<br />
24. Gurdon, J.B. The developmental capacity <strong>of</strong> nuclei taken from intestinal epi<strong>the</strong>lium cells<br />
<strong>of</strong> feeding tadpoles. J. Embryol. Exp. Morphol. 10, 622-640 (1962).<br />
25. Gurdon, J.B. & Melton, D.A. Nuclear reprogramming in cells. Science 322, 1811-1815<br />
(2008).<br />
26. Hanna, J. et al. Treatment <strong>of</strong> sickle cell anemia mouse model with iPS cells generated<br />
from autologous skin. Science 318, 1920-1923 (2007).<br />
27. Hochedlinger, K. et al. Reprogramming <strong>of</strong> a melanoma genome by nuclear<br />
transplantation. Genes Dev. 18, 1875-1885 (2004).<br />
28. Hochedlinger, K. & Jaenisch, R. Monoclonal mice generated by nuclear transfer from<br />
mature B and T donor cells. Nature 415, 1035-1038 (2002).<br />
29. Hochedlinger, K. & Jaenisch, R. Nuclear reprogramming and pluripotency. Nature 441,<br />
1061-1067 (2006).<br />
30. Huangfu, D. et al. Induction <strong>of</strong> pluripotent stem cells by defined factors is greatly<br />
improved by small-molecule compounds. Nat. Biotechnol. 26, 795-797 (2008a).<br />
11
31. Huangfu, D. et al. Induction <strong>of</strong> pluripotent stem cells from primary human fibroblasts<br />
with only Oct4 and Sox2. Nat. Biotechnol. 26, 1269-1275 (2008b).<br />
32. Imamura, M. et al. Transcriptional repression and DNA hypermethylation <strong>of</strong> a small set<br />
<strong>of</strong> ES cell marker genes in male germline stem cells. BMC Dev. Biol. 6, 34 (2006).<br />
33. Jackson, M. et al. Severe global DNA hypomethylation blocks differentiation and<br />
induces histone hyperacetylation in embryonic stem cells. Mol. Cell Biol. 24, 8862-8871<br />
(2004).<br />
34. Jaenisch, R. Human cloning—<strong>the</strong> science and ethics <strong>of</strong> nuclear transplantation. N. Engl.<br />
J. Med. 351, 2787-2791 (2004).<br />
35. Jaenisch, R. & Young, R. Stem cells, <strong>the</strong> molecular circuitry <strong>of</strong> pluripotency and nuclear<br />
reprogramming. Cell 132, 567-582 (2008).<br />
36. Jenuwein, T. & Allis, C.D. Translating <strong>the</strong> histone code. Science 293, 1074-1080 (2001).<br />
37. Kimura, H., Tada, M., Nakatsuji, N. & Tada, T. Histone code modifications on<br />
pluripotential nuclei <strong>of</strong> reprogrammed somatic cells. Mol. Cell. Biol. 24, 5710–5720<br />
(2004).<br />
38. Kouzarides, T. Chromatin modifications and <strong>the</strong>ir function. Cell 128, 693-705 (2007).<br />
39. Lee, J.H., Hart, S.R., & Skalnik, D.G. Histone deacetylase activity is required for<br />
embryonic stem cell differentiation. Genesis 38, 32-38 (2004).<br />
40. Li, E., Bestor, T.H., & Jaenisch, R. Targeted mutation <strong>of</strong> <strong>the</strong> DNA methyltransferase<br />
gene results in embryonic lethality. Cell 69, 915-926 (1992).<br />
41. Li, J., Ishii, T., Feinstein, P. & Mombaerts, P. Odorant receptor gene choice is reset by<br />
nuclear transfer from mouse olfactory sensory neurons. Nature 428, 393-399 (2004).<br />
42. Li, X., Kato, Y., Tsuji, Y., & Tsunoda, Y. The effects <strong>of</strong> trichostatin A on mRNA<br />
expression <strong>of</strong> chromatin structure-, DNA methylation-, and development-related genes in<br />
cloned mouse blastocysts. Cloning and Stem Cells 10, 133-142 (2008).<br />
43. Lu, R. et al. Systems-level dynamic analyses <strong>of</strong> fate change in murine embryonic stem<br />
cells. Nature 462, 358-362 (2009).<br />
44. Maherali, N. et al. (2007). Directly reprogrammed fibroblasts show global epigenetic<br />
reprogramming and widespread tissue contribution. Cell Stem Cell 1, 55-70.<br />
45. Matsumura, H. et al. Targeted chromosome elimination from ES-somatic hybrid cells.<br />
Nat. Methods 4, 23-25 (2007).<br />
12
46. Meshorer, E., Yellajoshula, D., George, E. et al. Hyperdynamic plasticity <strong>of</strong> chromatin<br />
proteins in pluripotent embryonic stem cells. Dev. Cell 10, 105-116 (2006).<br />
47. Mikkelsen, T.S., Ku, M., Jaffe, D.B., Issac, B., Lieberman, E. et al. Genome-wide maps<br />
<strong>of</strong> chromatin state in pluripotent and lineage-committed cells. Nature 448, 553-560<br />
(2007).<br />
48. Mikkelsen, T.S. et al., Dissecting direct reprogramming through integrative genomic<br />
analysis. Nature 454, 49-55 (2008).<br />
49. Okano, M., Bell, D.W., Haber, D.A. & Li, E. DNA methyltransferases Dnmt3a and<br />
Dnmt3b are essential for de novo methylation and mammalian development. Cell 99,<br />
247-257 (1999).<br />
50. Okita, K., Ichisaka, T., & Yamanaka, S. Generation <strong>of</strong> germline-competent induced<br />
pluripotent stem cells. Nature 448, 313-317 (2007).<br />
51. Okita, K., Nakagawa, M., Hyenjong, H., Ichisaka, T., & Yamanaka, S. Generation <strong>of</strong><br />
mouse induced pluripotent stem cells without viral vectors. Science 322, 949-953 (2008).<br />
52. Orkin, S.H. & Zon, L.I. Hematopoiesis: An evolving paradigm for stem cell biology. Cell<br />
132, 631-644 (2008).<br />
53. Park, I.H. et al. Disease-specific induced pluripotent stem cells. Cell 134, 877-886<br />
(2008).<br />
54. Pereira, C.F. et al. Heterokaryon-based reprogramming <strong>of</strong> human B lymphocytes for<br />
pluripotency requires Oct4 but not Sox2. PLoS Genet. 4, e1000170 (2008).<br />
55. Phair R.D. et al. Global nature <strong>of</strong> dynamic proteinchromatin interactions in vivo: Threedimensional<br />
genome scanning and dynamic interaction networks <strong>of</strong> chromatin proteins.<br />
Mol. Cell. Biol. 24, 6393-6402 (2004).<br />
56. Pomerantz, J.H., Mukherjee, S., Palermo, A.T. & Blau, H.M. Reprogramming to a<br />
muscle fate by fusion recapitulates differentiation. J. Cell Sci. 122, 1045-1053 (2009).<br />
57. Rideout, W.M., III, Hochedlinger K., Kyba, M., Daley, G.Q., & Jaenisch, R. Correction<br />
<strong>of</strong> a genetic defect by nuclear transplantation and combined cell and gene <strong>the</strong>rapy. Cell<br />
109, 17-27 (2002).<br />
58. Rybouchkin, A., Kato, Y. & Tsunoda, Y. Role <strong>of</strong> histone acetylation in reprogramming<br />
<strong>of</strong> somatic nuclei following nuclear transfer. Biol. Reprod. 74, 1083-1089 (2006).<br />
59. Silva, J., Chambers, I., Pollard, S. & Smith, A. Nanog promotes transfer <strong>of</strong> pluripotency<br />
after cell fusion. Nature 441, 997-1001 (2006).<br />
13
60. Silva, J. et al. Promotion <strong>of</strong> reprogramming to ground state pluripotency by signal<br />
inhibition. PLoS Biol. 6, e253 (2008).<br />
61. Simonsson, S. & Gurdon, J. DNA demethylation is necessary for <strong>the</strong> epigenetic<br />
reprogramming <strong>of</strong> somatic cell nuclei. Nat. Cell Biol. 6, 984-990 (2004).<br />
62. Slack, J.M.W. Origin <strong>of</strong> stem cells in organogenesis. Science 322, 1498-1501 (2008).<br />
63. Stadtfeld, M., Brennand, K. & Hochedlinger, K. Reprogramming <strong>of</strong> pancreatic beta cells<br />
into induced pluripotent stem cells. Curr. Biol. 18, 890-894 (2008a).<br />
64. Stadtfeld, M., Nagaya, M., Utakal, J., Weir, G., & Hochedlinger, K. Induced pluripotent<br />
stem cells generated without viral integration. Science 322, 945-949 (2008b).<br />
65. Surani, A.M., Hayashi, K., and Hajkova, P. Genetic and epigenetic regulators <strong>of</strong><br />
pluripotency. Cell 128, 747-762 (2007).<br />
66. Tada, M., Takahama, Y., Abe, K., Nakatsuji, N. & Tada, T. Nuclear reprogramming <strong>of</strong><br />
somatic cells by in vitro hybridization with ES cells. Curr. Biol. 11, 1553-1558 (2001).<br />
67. Tada, M., Morizane, A., Kimura, H., Kawasaki, H., Ainscough, J.F.X. et al. Pluripotency<br />
<strong>of</strong> reprogrammed somatic genomes in embryonic stem cells. Dev. Dyn. 227, 504-510<br />
(2003).<br />
68. Takahashi, K. & Yamanaka, S. Induction <strong>of</strong> pluripotent stem cells from mouse<br />
embryonic and adult fibroblast cultures by defined factors. Cell 126, 663-676 (2006).<br />
69. Takahashi, K. et al. Induction <strong>of</strong> pluripotent stem cells from adult human fibroblasts by<br />
defined factors. Cell 131, 1-12 (2007).<br />
70. Tecirlioglu, T., Guo, J. & Trounson, A. Interspecies somatic cell nuclear transfer and<br />
preliminary data for horse-cow/mouse iSCNT. Stem Cell Rev. 2, 277-287 (2006).<br />
71. Terranova, R., Pereira, C.F., Du Roure, C., Merkenschlager, M. & Fisher, A.G.<br />
Acquisition and extinction <strong>of</strong> gene expression programs are separable events in<br />
heterokaryon reprogramming. J. Cell Sci. 119, 2065-2072 (2006).<br />
72. Vakoc, C.R., Mandat, S.A., Olenchock, B.A., & Blobel, G.A. Histone H3 lysine 9<br />
methylation and HP1γ are associated with <strong>transcript</strong>ion elongation through mammalian<br />
chromatin. Mol. Cell. 19, 381-391 (2005).<br />
73. Vakoc, C.R., Sachdeva, M.M., Wang, H., & Blobel, G.A. Pr<strong>of</strong>ile <strong>of</strong> histone lysine<br />
methylation across transcribed mammalian chromatin. Mol. Cell. Biol. 26, 9185-9195<br />
(2006).<br />
14
74. Wakayama, T. et al. Differentiation <strong>of</strong> embryonic stem cell lines generated from adult<br />
somatic cells by nuclear transfer. Science 292, 740-743 (2001).<br />
75. Weintraub, H. et al. Activation <strong>of</strong> muscle-specific genes in pigment, nerve, fat, liver, and<br />
fibroblast cell lines by forced expression <strong>of</strong> MyoD. Proc. Natl. Acad. Sci. USA 86, 5434-<br />
5438 (1989).<br />
76. Wernig, M. et al. In vitro reprogramming <strong>of</strong> fibroblasts into a pluripotent ES-cell-like<br />
state. Nature 448, 318-324 (2007).<br />
77. Wernig, M. et al. Neurons derived from reprogrammed fibroblasts functionally integrate<br />
into <strong>the</strong> fetal brain and improve <strong>the</strong> symptoms <strong>of</strong> rats with Parkinson’s disease. Proc.<br />
Natl. Acad. Sci. USA 105, 5856-5861 (2008).<br />
78. Wilmut, I., Schnieke, A.E., McWhir, J., Kind, A.J. & Campbell, K.H. Viable <strong>of</strong>fspring<br />
derived from fetal and adult mammalian cells. Nature 385, 810-813 (1997).<br />
79. Wilmut, I. et al. Somatic cell nuclear transfer. Nature 419, 583-586 (2002).<br />
80. Xie, H., Min, Y., Feng, R. & Graf, T. Stepwise reprogramming <strong>of</strong> B cells into<br />
macrophages. Cell 117, 663-676 (2004).<br />
81. Yu, J. et al. Induced pluripotent stem cell lines derived from human somatic cells.<br />
Science 318, 1917-1920 (2007).<br />
82. Zhou, Q., Brown, J., Kanarek, A., Rajagopal, J. & Melton, D.A. In vivo reprogramming<br />
<strong>of</strong> adult pancreatic exocrine cells to β-cells. Nature 455, 627-632 (2008).<br />
83. Zvetkova, I. et al. Global hypomethylation <strong>of</strong> <strong>the</strong> genome in XX embryonic stem cells.<br />
Nat. Genet. 37, 1274-1279 (2005).<br />
15
Figure 1. Cells in <strong>the</strong> inner<br />
cell mass are pluripotent,<br />
meaning that <strong>the</strong>y can give<br />
rise to all three germ layers:<br />
neurons (ectoderm), red<br />
blood cells (mesoderm), and<br />
pancreatic cells (endoderm).<br />
Figure 2. Cell<br />
differentiation is associated<br />
with loss <strong>of</strong> cell potential<br />
and decreased cell division<br />
activity.<br />
Figure 3. DNA methylation,<br />
or <strong>the</strong> addition <strong>of</strong> methyl<br />
groups to specific cytosine<br />
bases, can repress or silence<br />
gene activity by preventing<br />
<strong>transcript</strong>ion factors from<br />
binding to a gene’s<br />
promoter.
Figure 4. Histone<br />
acetylation can decrease <strong>the</strong><br />
affinity <strong>of</strong> histones for DNA,<br />
allowing <strong>transcript</strong>ion<br />
factors to access and bind to<br />
a gene’s promoter.<br />
Figure 5. Although <strong>the</strong><br />
naturally occurring process<br />
<strong>of</strong> cell differentiation is<br />
unidirectional, experimental<br />
procedures have been<br />
identified that allow<br />
dramatic switches in gene<br />
expression and cell identity.<br />
Somatic cell nuclear transfer<br />
(top), cell fusion (middle),<br />
and direct reprogramming<br />
(bottom) are <strong>the</strong> three main<br />
methods.<br />
d<br />
Figure 6. With <strong>the</strong> exception<br />
<strong>of</strong> antibody-producing cells,<br />
all cells in <strong>the</strong> body are<br />
genetically identical, even<br />
though <strong>the</strong>ir distinct<br />
morphology and behavior is<br />
determined by a specific<br />
pattern <strong>of</strong> gene expression.
Figure 7. Nuclear<br />
reprogramming describes a<br />
change in <strong>the</strong> nuclear gene<br />
expression <strong>of</strong> one cell type<br />
to that <strong>of</strong> an unrelated cell<br />
type or embryo.<br />
Figure 8. Nuclear<br />
reprogramming provides a<br />
model for <strong>the</strong> generation <strong>of</strong><br />
patient-specific cells for<br />
transplantation. Somatic<br />
cells from a patient can be<br />
reprogrammed into immunecompatible<br />
cells that behave<br />
like pluripotent stem cells.<br />
These cells can be expanded<br />
and induced to differentiate<br />
into a needed cell type in<br />
vitro, providing patientspecific<br />
cells for<br />
replacement <strong>of</strong> diseased or<br />
damaged tissues.<br />
Figure 9. A needed cell type<br />
may be directly generated<br />
from patient cells without<br />
<strong>the</strong> need for intermediate<br />
pluripotent cells.
Figure 10. In somatic cell nuclear transfer (SCNT), <strong>the</strong> nucleus <strong>of</strong> a somatic cell is injected into an egg that has had its own<br />
chromatin removed. Although some <strong>of</strong> <strong>the</strong> somatic proteins are transferred with <strong>the</strong> nucleus, <strong>the</strong> large volume <strong>of</strong> egg cytoplasm<br />
dilutes <strong>the</strong> somatic <strong>transcript</strong>ional program, allowing <strong>the</strong> embryonic <strong>transcript</strong>ional program to dominate.<br />
Figure 11. When two cells<br />
are fused, a heterokaryon<br />
with two distinct nuclei is<br />
formed. In some cases,<br />
nuclei from fused partner<br />
cells merge, leading to a<br />
stable hybrid cell.<br />
Figure 12. In a small<br />
percentage <strong>of</strong> hybrid cells,<br />
stem cell-specific genes that<br />
were previously silenced in<br />
<strong>the</strong> somatic cell are<br />
reactivated from somatic<br />
chromosomes.<br />
Reprogrammed cells that<br />
display embryonic stem cell<br />
morphology, growth, and<br />
gene expression have been<br />
isolated by using a genetic<br />
marker that indicates<br />
reactivation <strong>of</strong> stem-cellspecific<br />
genes.
Figure 13. When<br />
hepatocytes are fused with<br />
myotubes, non-muscle<br />
nuclei in <strong>the</strong> resulting<br />
heterokaryons express<br />
muscle genes, indicating that<br />
<strong>the</strong> muscle phenotype is<br />
dominant.<br />
Figure 14. Liver cells, as<br />
well as o<strong>the</strong>r cell types, have<br />
been redirected to a musclelike<br />
<strong>transcript</strong>ional program<br />
through forced expression <strong>of</strong><br />
<strong>the</strong> muscle <strong>transcript</strong>ion<br />
factor MyoD.<br />
Figure 15. Retroviral<br />
transfection <strong>of</strong> Oct-3/4,<br />
Sox2, Klf4, and c-Myc can<br />
reprogram murine and<br />
human fibroblasts into<br />
induced pluripotent stem<br />
(iPS) cells. This process is<br />
termed pluripotent<br />
reprogramming.
Figure 16. In iPS cells,<br />
endogenous ES-cell-specific<br />
genes are reactivated and<br />
remain active even after<br />
viruses have been silenced.<br />
Figure 17. Adenoviral<br />
infection with Ngn3, Pdx1,<br />
and Mafa can lead to in vivo<br />
reprogramming <strong>of</strong> pancreatic<br />
exocrine cells into insulinproducing<br />
!-cells.
Figure 18. By understanding<br />
<strong>the</strong> molecular mechanisms<br />
behind reprogramming, we<br />
can develop more efficient,<br />
direct strategies for<br />
generating patient-specific<br />
cells.














