Downloaded from http://sp.lyellcollection.org/ at CAPES on May 3, <strong>2013</strong>RAUISUCHIAamong rauisuchids and poposauroids, and therelationships of these two clades will be expandedon below. The relationships of the remainingrauisuchians included in Brusatte <strong>et</strong> <strong>al</strong>. (2010)and Butler <strong>et</strong> <strong>al</strong>. (2011) and some included in<strong>Nesbitt</strong> (2011) – Arganasuchus dutuiti, Fasolasuchustenax, Stagonosuchus nyassicus, Ticinosuchusferox, Saurosuchus g<strong>al</strong>ilei, Batrachotomus kupferzellensisand Prestosuchus chiniquensis – will notbe discussed further as the relationships of thes<strong>et</strong>axa are far from understood at this point in time.PoposauroideaThe phylogen<strong>et</strong>ic an<strong>al</strong>yses of <strong>Nesbitt</strong> (2003),<strong>Nesbitt</strong> & Norell (2006), Weinbaum & Hungerbühler(2007), Brusatte <strong>et</strong> <strong>al</strong>. (2010) and <strong>Nesbitt</strong> (2011)have examined the relationships of a diverse arrayof rauisuchians (Figs 3 & 4) and have found agroup of rauisuchians with affinities to Poposaurusgracilis. This clade, Poposauroidea (see definitionabove), is one of the best-supported subgroups ofrauisuchians. Poposauroidea (equiv<strong>al</strong>ent to ‘groupX’ of <strong>Nesbitt</strong> 2007) includes an eclectic array ofarchosaurs ranging in body size, locomotor strategiesand seemingly di<strong>et</strong>ary ecologies, with a stratigraphicrange from the end of the Early Triassic(Ctenosauriscus koeneni) to the end of the Triassic(Effigia okeeffeae). Taxonomic<strong>al</strong>ly, Poposauroideacomprises Qianosuchus mixtus, Lotosaurusadentus, Poposaurus gracilis, Poposaurus langstoni,Ctenosauriscidae and Shuvosauridae.Both Brusatte <strong>et</strong> <strong>al</strong>. (2010) and <strong>Nesbitt</strong> (2011)recovered a similar s<strong>et</strong> of taxa within Poposauroidea,but there are a few important differencesb<strong>et</strong>ween the studies. Brusatte <strong>et</strong> <strong>al</strong>. (2010) foundYarasuchus deccanensis as the sister to <strong>al</strong>l othermembers of their Poposauroidea, whereas <strong>Nesbitt</strong>(2011) did not include the taxon in his phylogen<strong>et</strong>ican<strong>al</strong>ysis. A recent examination by one of us(JBD) suggests that the type series and referredspecimens of Yarasuchus deccanensis describedby Sen (2005) may include a minimum of twodifferent archosauromorph taxa, and it is unclearwh<strong>et</strong>her any of this materi<strong>al</strong> even pertains to a pseudosuchian.Nevertheless, the position of Yarasuchusdeccanensis is only weakly resolved in Brusatte<strong>et</strong> <strong>al</strong>.’s (2010) an<strong>al</strong>ysis. The sister taxon to <strong>al</strong>lother (non-Yarasuchus) poposauroids in Brusatte<strong>et</strong> <strong>al</strong>.’s (2010) an<strong>al</strong>ysis, and the sister taxon to <strong>al</strong>lother poposauroids in <strong>Nesbitt</strong>’s (2011) an<strong>al</strong>ysis, isQianosuchus mixtus. The monophyly of Qianosuchusmixtus + other poposauroids is supported bya number of features rare among pseudosuchiansincluding, but not limited to, a maxilla that bordersthe extern<strong>al</strong> naris, the entrance of the cerebr<strong>al</strong>branches of the intern<strong>al</strong> carotid artery positionedon the ventr<strong>al</strong> surface of the braincase, and at leastthree sacr<strong>al</strong> vertebrae (only two are present in Qianosuchusmixtus) (Fig. 1). Qianosuchus mixtus is theonly poposauroid to bear osteoderms, a characterstate that is apomorphic<strong>al</strong>ly absent in <strong>al</strong>l otherpoposauroids.The recently phylogen<strong>et</strong>ic<strong>al</strong>ly defined Ctenosauriscidae(Butler <strong>et</strong> <strong>al</strong>. 2011) comprises a cladeof sail-backed poposauroids (Fig. 1) glob<strong>al</strong>ly widespreadfrom the end of the Early Triassic to the endof the Anisian (<strong>Nesbitt</strong> 2003; Butler <strong>et</strong> <strong>al</strong>. 2009,2011; <strong>Nesbitt</strong> 2011). Presacr<strong>al</strong>, sacr<strong>al</strong> and anteriorcaud<strong>al</strong> vertebrae with extremely elongated neur<strong>al</strong>spines (which would have supported a sail) havebeen known for more than a century from theEarly–Middle Triassic deposits of Germany, Africaand China, but the affinities of these anim<strong>al</strong>s werepoorly understood. After the discovery of a wellpreservedspecimen of Arizonasaurus babbitti,<strong>Nesbitt</strong> (2003, 2005a) hypothesized that sail-backedarchosaurs formed a clade among Poposaurus-lik<strong>et</strong>axa. Later work corroborated this hypothesis(Butler <strong>et</strong> <strong>al</strong>. 2011; <strong>Nesbitt</strong> 2011), but hypothesized<strong>al</strong>so that the sail-backed form Lotosaurus adentuswas not part of Ctenosauriscidae but instead wasmore closely related to Shuvosauridae, and thusmay have evolved its sail convergently. Butler<strong>et</strong> <strong>al</strong>. (2011) presented revised an<strong>al</strong>yses of the Brusatte<strong>et</strong> <strong>al</strong>. (2010) and <strong>Nesbitt</strong> (2011) data s<strong>et</strong>s thatincluded greater taxon sampling, and found that Ctenosauriscus,Hypselorhachis mirabilis, a GermanAnisian group of specimen referred to a singl<strong>et</strong>axon (¼‘W<strong>al</strong>dhaus taxon’), Xilousuchus sapingensis,Arizonasaurus babbitti and Bromsgroveiaw<strong>al</strong>keri comprise the Ctenosauriscidae. This cladeis diagnosed by sever<strong>al</strong> character states relating tothe neur<strong>al</strong> spines, including dors<strong>al</strong> neur<strong>al</strong> spinesthat are more than seven times t<strong>al</strong>ler than centrumheight and strongly curved dors<strong>al</strong> neur<strong>al</strong> spines.Fig. 4. Rauisuchian skull morphology: (a) two skulls of the newly named Decuriasuchus quartacolonia (MCNPV10105c, d) from França <strong>et</strong> <strong>al</strong>. (2011); (b) likely skull of Prestosuchus chiniquensis (UFRGS 0156-T) (reversed);(c) holotype skull of Qianosuchus mixtus (IVPP V14300); (d) skull of the poposauroid Lotosaurus adentus (IVPP V48013); (e) holotype maxilla of Postosuchus kirkpatricki (TTUP 9000); (f ) holotype maxilla of Teratosaurus suevicus(NHMUK 38646) (reversed); (g) holotype maxilla of Polonosuchus silesiacus (ZPAL Ab III/563); (h) referred maxillaof Fasolasuchus tenax (PVL 3851); (i) holotype maxilla of Batrachotomus kuperferzellensis (SMNS 52970);(j) holotype maxilla of Xilousuchus sapingensis (IVPP V6024); (k) holotype skel<strong>et</strong>on of Ticinosuchus ferox (PIZT2817). Sc<strong>al</strong>e bars: 5 cm (a–j); 10 cm (k). See appendix for institution<strong>al</strong> abbreviations.
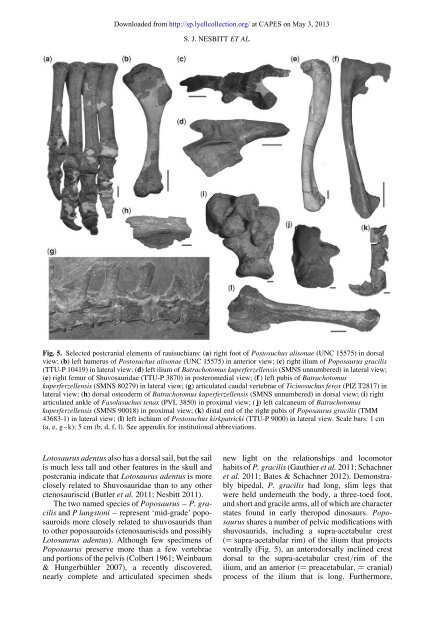
Downloaded from http://sp.lyellcollection.org/ at CAPES on May 3, <strong>2013</strong>S. J. NESBITT ET AL.Fig. 5. Selected postcrani<strong>al</strong> elements of rauisuchians: (a) right foot of Postosuchus <strong>al</strong>isonae (UNC 15575) in dors<strong>al</strong>view; (b) left humerus of Postosuchus <strong>al</strong>isonae (UNC 15575) in anterior view; (c) right ilium of Poposaurus gracilis(TTU-P 10419) in later<strong>al</strong> view; (d) left ilium of Batrachotomus kuperferzellensis (SMNS unnumbered) in later<strong>al</strong> view;(e) right femur of Shuvosauridae (TTU-P 3870) in posteromedi<strong>al</strong> view; (f ) left pubis of Batrachotomuskuperferzellensis (SMNS 80279) in later<strong>al</strong> view; (g) articulated caud<strong>al</strong> vertebrae of Ticinosuchus ferox (PIZ T2817) inlater<strong>al</strong> view; (h) dors<strong>al</strong> osteoderm of Batrachotomus kuperferzellensis (SMNS unnumbered) in dors<strong>al</strong> view; (i) rightarticulated ankle of Fasolasuchus tenax (PVL 3850) in proxim<strong>al</strong> view; ( j) left c<strong>al</strong>caneum of Batrachotomuskuperferzellensis (SMNS 90018) in proxim<strong>al</strong> view; (k) dist<strong>al</strong> end of the right pubis of Poposaurus gracilis (TMM43683-1) in later<strong>al</strong> view; (l) left ischium of Postosuchus kirkpatricki (TTU-P 9000) in later<strong>al</strong> view. Sc<strong>al</strong>e bars: 1 cm(a, e, g–k); 5 cm (b, d, f, l). See appendix for institution<strong>al</strong> abbreviations.Lotosaurus adentus <strong>al</strong>so has a dors<strong>al</strong> sail, but the sailis much less t<strong>al</strong>l and other features in the skull andpostcrania indicate that Lotosaurus adentus is moreclosely related to Shuvosauridae than to any otherctenosauriscid (Butler <strong>et</strong> <strong>al</strong>. 2011; <strong>Nesbitt</strong> 2011).The two named species of Poposaurus – P. gracilisand P langstoni – represent ‘mid-grade’ poposauroidsmore closely related to shuvosaurids thanto other poposauroids (ctenosauriscids and possiblyLotosaurus adentus). Although few specimens ofPoposaurus preserve more than a few vertebraeand portions of the pelvis (Colbert 1961; Weinbaum& Hungerbühler 2007), a recently discovered,nearly compl<strong>et</strong>e and articulated specimen shedsnew light on the relationships and locomotorhabits of P. gracilis (Gauthier <strong>et</strong> <strong>al</strong>. 2011; Schachner<strong>et</strong> <strong>al</strong>. 2011; Bates & Schachner 2012). Demonstrablybiped<strong>al</strong>, P. gracilis had long, slim legs thatwere held underneath the body, a three-toed foot,and short and gracile arms, <strong>al</strong>l of which are characterstates found in early theropod dinosaurs. Poposaurusshares a number of pelvic modifications withshuvosaurids, including a supra-ac<strong>et</strong>abular crest(¼ supra-ac<strong>et</strong>abular rim) of the ilium that projectsventr<strong>al</strong>ly (Fig. 5), an anterodors<strong>al</strong>ly inclined crestdors<strong>al</strong> to the supra-ac<strong>et</strong>abular crest/rim of theilium, and an anterior (¼ preac<strong>et</strong>abular, ¼ crani<strong>al</strong>)process of the ilium that is long. Furthermore,