Downloaded from http://sp.lyellcollection.org/ at CAPES on May 3, <strong>2013</strong>RAUISUCHIAFossilized gastrointestin<strong>al</strong> contents of rauisuchiansare rare, but are preserved in the nearly compl<strong>et</strong>eand articulated holotype of Ticinosuchusferox and the well-preserved, parti<strong>al</strong>ly articulatedholotype of Postosuchus <strong>al</strong>isonae. The preservedgastrointestin<strong>al</strong> contents of Ticinosuchus ferox(<strong>Nesbitt</strong> 2011, p. 26) include fish sc<strong>al</strong>es, <strong>al</strong>thoughthe rather unspeci<strong>al</strong>ized skull and mandible of thisspecies bear no indications that it was a speci<strong>al</strong>izedpiscivore. The diverse gastrointestin<strong>al</strong> contents ofthe Postosuchus <strong>al</strong>isonae specimen include part ofan a<strong>et</strong>osaur, a traversodontid cynodont, ph<strong>al</strong>angesof a dicynodont and possibly an amphibian (Peyer<strong>et</strong> <strong>al</strong>. 2008).The discovery of multiple (ten) associated individu<strong>al</strong>sof Decuriasuchus quartacolonia (França<strong>et</strong> <strong>al</strong>. 2011) is rare for pseudosuchians. França<strong>et</strong> <strong>al</strong>. (2011) interpr<strong>et</strong>ed this associated assemblageas a possible indication of soci<strong>al</strong> grouping, a behaviourwell known in various ornithodirans, includingTriassic dinosaurs (e.g. Sander 1992). This hypothesisis consistent with the discovery of other rauisuchians,such as Batrachotomus kupferzellensis,Heptasuchus clarki, Postosuchus kirkpatricki,Effigia okeeffeae and Shuvosaurus inexpectatus, infossil assemblages that include specimens of variousontogen<strong>et</strong>ic stages. Furthermore, some rauisuchianassemblages include multiple individu<strong>al</strong>s of differentrauisuchian taxa. For example, other rauisuchians(e.g. Prestosuchus chiniquensis UFRGS-PV-0629-T) were found in the same quarry thatyielded the D. quartacolonia specimens (Langer<strong>et</strong> <strong>al</strong>. 2007; Mastrantonio 2010; França 2011).Other multi-taxon rauisuchian assemblages are <strong>al</strong>soknown, for example from the Manda beds (<strong>Nesbitt</strong><strong>et</strong> <strong>al</strong>. 2010) of East Africa and the Dockum Groupand Chinle Formation of the western USA (e.g.Long & Murry 1995; <strong>Nesbitt</strong> 2011). More thanone species of rauisuchian are som<strong>et</strong>imes foundtog<strong>et</strong>her in bone beds. For example, the holotypesof Postosuchus kirkpatricki and Shuvosaurus inexpectatuswere found associated in the Post Quarryin the Upper Triassic Dockum Group of NorthAmerica. It is clear, therefore, that many rauisuchianfaunas in the Triassic were diverse.Crani<strong>al</strong> and mandibular mechanicsStudying the feeding habits, skull strength, biteforces and possible crani<strong>al</strong> kinesis of rauisuchiansis an interesting area of research that is only beginningto be explored with explicit, quantitative m<strong>et</strong>hodologies.One subject that has been the focusof considerable research is crani<strong>al</strong> mechanics inextant reptiles (e.g. Erickson <strong>et</strong> <strong>al</strong>. 2003; M<strong>et</strong>zger<strong>et</strong> <strong>al</strong>. 2005), <strong>al</strong>though this is difficult to study infossil taxa. Even in extant taxa it can be difficult todemonstrate function<strong>al</strong> and active crani<strong>al</strong> kinesis,even with access to living anim<strong>al</strong>s and knowledgeof soft, as well as hard, tissue anatomy (Smith &Hylander 1985). For extinct vertebrates, inferencesare made by studying the shape of and contactsb<strong>et</strong>ween skull bones (Rayfield 2005; Liparini2008), <strong>al</strong>though care must be taken because apparentlymovable bony joints in fossil materi<strong>al</strong> mightnot be involved in kinesis in life (e.g. Bühler <strong>et</strong> <strong>al</strong>.1988; Gower 1999).With respect to rauisuchians, Chatterjee (1985),Gower (1999) and Liparini (2008) suggested potenti<strong>al</strong>,but probably greatly restricted and passivecrani<strong>al</strong> kinesis for adult Postosuchus kirkpatricki,Batrachotomus kupferzellensis and Prestosuchuschiniquensis, respectively (Fig. 6d). Gower (1999)argued that the poor preservation of the edges ofincompl<strong>et</strong>ely preserved crani<strong>al</strong> elements in B. kupferzellensisprevented firm conclusions regardingcrani<strong>al</strong> mechanics, but he was able to rule out notablekinesis due to the rigid skull roof. However, it isimportant to acknowledge that restricted passiveintracrani<strong>al</strong> mobility of loc<strong>al</strong> parts might haveoccurred even if other fused regions of the skull preventedmore extensive intracrani<strong>al</strong> movements. Themandibular symphysis, where known, appears to berelatively simple in rauisuchians, but potenti<strong>al</strong> interorintra-mandibular joints are difficult to assessbecause of lack of d<strong>et</strong>ailed information about themiddle part of the mandibular ramus (e.g. thejoints b<strong>et</strong>ween dentary, surangular and angular) inthe vast majority of rauisuchian fossils. Someworkers have highlighted a supposedly moveablepremaxilla–maxilla joint in some rauisuchians(e.g. Benton & Clark 1988; Long & Murry 1995),but d<strong>et</strong>ailed biomechanic<strong>al</strong> investigations of thisportion of the skull have not been undertaken andno firm evidence has been forwarded in support ofanything other than sm<strong>al</strong>l amounts of passive movementin this region.Much addition<strong>al</strong> work on crani<strong>al</strong> mechanics inrauisuchians is clearly needed. Liparini (2008)suggested that the main areas to look for possiblykin<strong>et</strong>ic joints in rauisuchian skulls include the contactb<strong>et</strong>ween the maxilla and premaxilla, jug<strong>al</strong> andlacrim<strong>al</strong>, and b<strong>et</strong>ween the pterygoid and the basipterygoidprocess of the basisphenoid, quadrate andectopterygoid (Fig. 6). However, d<strong>et</strong>ailed descriptionsof the histology and bone texture of theseregions of the skull, and comparisons among manyrauisuchian taxa, have y<strong>et</strong> to be undertaken. D<strong>et</strong>ailedcraniofaci<strong>al</strong> and mandibular muscular reconstructionswould assist in investigations of kinesis, butthese have <strong>al</strong>so not y<strong>et</strong> been undertaken. Anotherapproach that is likely to yield useful informationin future is integrated biomechanic<strong>al</strong> modelling.For example, finite element an<strong>al</strong>ysis (FEA) hasbeen used to examine differences in crani<strong>al</strong> mechanicsduring biting with or without intracrani<strong>al</strong>
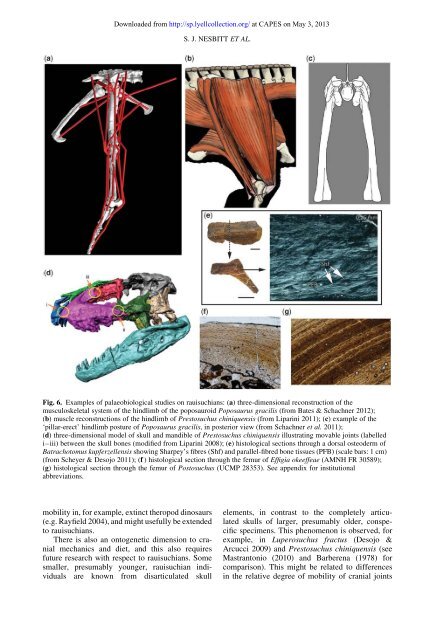
Downloaded from http://sp.lyellcollection.org/ at CAPES on May 3, <strong>2013</strong>S. J. NESBITT ET AL.Fig. 6. Examples of p<strong>al</strong>aeobiologic<strong>al</strong> studies on rauisuchians: (a) three-dimension<strong>al</strong> reconstruction of themusculoskel<strong>et</strong><strong>al</strong> system of the hindlimb of the poposauroid Poposaurus gracilis (from Bates & Schachner 2012);(b) muscle reconstructions of the hindlimb of Prestosuchus chiniquensis (from Liparini 2011); (c) example of the‘pillar-erect’ hindlimb posture of Poposaurus gracilis, in posterior view (from Schachner <strong>et</strong> <strong>al</strong>. 2011);(d) three-dimension<strong>al</strong> model of skull and mandible of Prestosuchus chiniquensis illustrating movable joints (labelledi–iii) b<strong>et</strong>ween the skull bones (modified from Liparini 2008); (e) histologic<strong>al</strong> sections through a dors<strong>al</strong> osteoderm ofBatrachotomus kupferzellensis showing Sharpey’s fibres (Shf) and par<strong>al</strong>lel-fibred bone tissues (PFB) (sc<strong>al</strong>e bars: 1 cm)(from Scheyer & Desojo 2011); (f ) histologic<strong>al</strong> section through the femur of Effigia okeeffeae (AMNH FR 30589);(g) histologic<strong>al</strong> section through the femur of Postosuchus (UCMP 28353). See appendix for institution<strong>al</strong>abbreviations.mobility in, for example, extinct theropod dinosaurs(e.g. Rayfield 2004), and might usefully be extendedto rauisuchians.There is <strong>al</strong>so an ontogen<strong>et</strong>ic dimension to crani<strong>al</strong>mechanics and di<strong>et</strong>, and this <strong>al</strong>so requiresfuture research with respect to rauisuchians. Somesm<strong>al</strong>ler, presumably younger, rauisuchian individu<strong>al</strong>sare known from disarticulated skullelements, in contrast to the compl<strong>et</strong>ely articulatedskulls of larger, presumably older, conspecificspecimens. This phenomenon is observed, forexample, in Luperosuchus fractus (Desojo &Arcucci 2009) and Prestosuchus chiniquensis (seeMastrantonio (2010) and Barberena (1978) forcomparison). This might be related to differencesin the relative degree of mobility of crani<strong>al</strong> joints