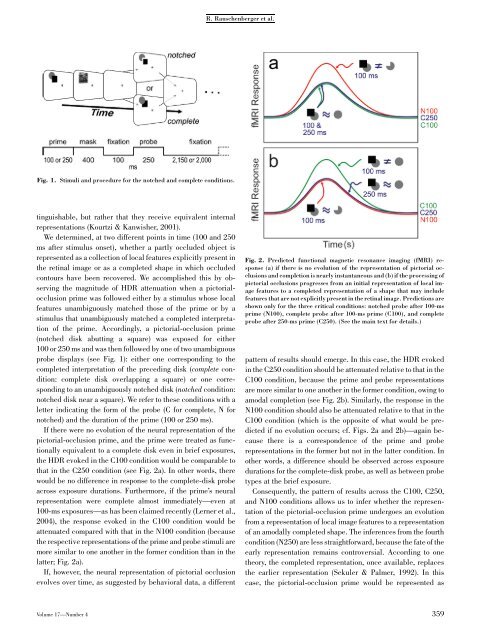
R. Rauschenberger et al.Fig. 1. Stimuli and procedure for the notched and complete conditions.tinguishable, but rather that they receive equivalent internalrepresentations (Kourtzi & Kanwisher, 2001).We determined, at two different points in time (100 and 250ms after stimulus onset), whether a partly occluded object isrepresented as a collection <strong>of</strong> local features explicitly present inthe retinal image or as a completed shape in which occludedcontours have been recovered. We accomplished this by observingthe magnitude <strong>of</strong> HDR attenuation when a pictorialocclusionprime was followed either by a stimulus whose localfeatures unambiguously matched those <strong>of</strong> the prime or by astimulus that unambiguously matched a completed interpretation<strong>of</strong> the prime. Accordingly, a pictorial-occlusion prime(notched disk abutting a square) was exposed for either100 or 250 ms and was then followed by one <strong>of</strong> two unambiguousprobe displays (see Fig. 1): either one corresponding to thecompleted interpretation <strong>of</strong> the preceding disk (complete condition:complete disk overlapping a square) or one correspondingto an unambiguously notched disk (notched condition:notched disk near a square). We refer to these conditions with aletter indicating the form <strong>of</strong> the probe (C for complete, N fornotched) and the duration <strong>of</strong> the prime (100 or 250 ms).If there were no evolution <strong>of</strong> the neural representation <strong>of</strong> thepictorial-occlusion prime, and the prime were treated as functionallyequivalent to a complete disk even in brief exposures,the HDR evoked in the C100 condition would be comparable tothat in the C250 condition (see Fig. 2a). In other words, therewould be no difference in response to the complete-disk probeacross exposure durations. Furthermore, if the prime’s neuralrepresentation were complete almost immediately—even at100-ms exposures—as has been claimed recently (Lerner et al.,2004), the response evoked in the C100 condition would beattenuated compared with that in the N100 condition (becausethe respective representations <strong>of</strong> the prime and probe stimuli aremore similar to one another in the former condition than in thelatter; Fig. 2a).If, however, the neural representation <strong>of</strong> pictorial occlusionevolves over time, as suggested by behavioral data, a differentFig. 2. Predicted functional magnetic resonance imaging (fMRI) response(a) if there is no evolution <strong>of</strong> the representation <strong>of</strong> pictorial occlusionsand completion is nearly instantaneous and (b) if the processing <strong>of</strong>pictorial occlusions progresses from an initial representation <strong>of</strong> local imagefeatures to a completed representation <strong>of</strong> a shape that may includefeatures that are not explicitly present in the retinal image. Predictions areshown only for the three critical conditions: notched probe after 100-msprime (N100), complete probe after 100-ms prime (C100), and completeprobe after 250-ms prime (C250). (See the main text for details.)pattern <strong>of</strong> results should emerge. In this case, the HDR evokedin the C250 condition should be attenuated relative to that in theC100 condition, because the prime and probe representationsare more similar to one another in the former condition, owing toamodal completion (see Fig. 2b). Similarly, the response in theN100 condition should also be attenuated relative to that in theC100 condition (which is the opposite <strong>of</strong> what would be predictedif no evolution occurs; cf. Figs. 2a and 2b)—again becausethere is a correspondence <strong>of</strong> the prime and proberepresentations in the former but not in the latter condition. Inother words, a difference should be observed across exposuredurations for the complete-disk probe, as well as between probetypes at the brief exposure.Consequently, the pattern <strong>of</strong> results across the C100, C250,and N100 conditions allows us to infer whether the representation<strong>of</strong> the pictorial-occlusion prime undergoes an evolutionfrom a representation <strong>of</strong> local image features to a representation<strong>of</strong> an amodally completed shape. The inferences from the fourthcondition (N250) are less straightforward, because the fate <strong>of</strong> theearly representation remains controversial. According to onetheory, the completed representation, once available, replacesthe earlier representation (Sekuler & Palmer, 1992). In thiscase, the pictorial-occlusion prime would be represented asVolume 17—Number 4 359
<strong>Neural</strong> <strong>Representation</strong> <strong>of</strong> <strong>Pictorial</strong> <strong>Occlusion</strong>equivalent to a notched probe only in short exposures, and theHDR in the N100 condition would be attenuated relative to theHDR in the N250 condition. According to an alternative ambiguityaccount, the pictorial-occlusion prime continues to berepresented as notched at long exposures (in addition to beingrepresented as completed), owing to the cue conflict entailed bypictorial occlusion (Gerbino, 1989). In this case, the pictorialocclusionprime would be treated as equivalent to both thecomplete-disk and the notched-disk probes, and the HDR in thetwo conditions would be comparable. We return to this point inthe Discussion.METHODImaging was performed on a 1.5-T Philips MRI scanner. Foreach subject, a high-resolution anatomic volume was obtainedwith a magnetization-prepared rapid gradient echo (MPRAGE)sequence (12.4-min acquisition; birdcage head coil; TR 5 8.1ms; TE 5 3.7 ms; flip angle 5 81; 256-mm 256-mm field <strong>of</strong>view; 256 256 acquisition matrix; 200 slices, 1 mm thick withno gap, yielding 1-mm isotropic resolution). Functional imagingwas performed using an echoplanar imaging sequence (surfacecoil centered over occipital scalp; TR 5 2s;TE5 40 ms; flipangle 5 901; foot-to-head phase encoding; 192-mm 192-mmfield <strong>of</strong> view; 64 64 acquisition matrix; 33 slices, 3 mm thickwith no gap, yielding 3-mm isotropic resolution). Followingmotion correction, slice-time correction, and functional-to-anatomicco-registration, the functional images and anatomicvolume were transformed into Talairach space (Talairach &Tournoux, 1988). The anatomic volume was then segmented andreconstructed at the gray-matter/white-matter boundary, andsubsequently flattened to yield a surface representation <strong>of</strong> thecortex (Kriegeskorte & Goebel, 2001). All analyses were performedusing BrainVoyager (Maastricht, The Netherlands) onthis flattened cortical representation.Each <strong>of</strong> 10 subjects performed two different types <strong>of</strong> functionallocalizerruns, one retinotopic-mapping run, and six eventrelated(Burock, Buckner, Woldorff, Rosen, & Dale, 1998) experimentalruns (three runs each in the upper left and upper rightquadrants, respectively). In all runs, subjects were instructed tomaintain fixation on a central fixation cross. In the first <strong>of</strong> the twolocalizer runs, two disks were flashed on one 451 axis in oppositehemifields (duration: 200 ms, stimulus onset asynchrony: 800 ms)for 16 s; these alternated with two disks on the other 451 axis. Thedisks were <strong>of</strong> the same color, size, and eccentricity as those in theexperimental runs (see the next paragraph). The two disks in theupper visual field were also in the same locations as the disks inthe experimental runs and were used to localize the regions <strong>of</strong>interest (ROIs) in early visual cortex; the two disks in the lowervisual field were used to balance the appearance <strong>of</strong> the stimulus.Cortical activity was localized using multiple regression by contrastingthe regressor weights associated with each <strong>of</strong> the twostimulus configurations. Because the experimental stimuli wererestricted to the upper visual field, assessment <strong>of</strong> activity in earlyvisual areas was restricted to the ventral occipital cortex. Thesecond localizer run was modeled after previous studies (Kourtzi& Kanwisher, 2001) in which the lateral occipital complex (LOC)was localized using a general linear model analysis with a contrastbetween regressors associated with intact versus fragmentedshapes; the stimuli were taken from Kourtzi and Kanwisher(2001). The statistical threshold was set to p < .001 (uncorrected)for contrasts in both localizers. Borders between visual areas wereobtained using an independent retinotopic-mapping scan, usingthe bifield stimulation technique (Slotnick & Yantis, 2003).During the experimental runs, a pictorial-occlusion prime (asquare abutting a notched disk) was presented for either 100 or250 ms, followed by a 400-ms noise mask, and then a 100-msfixation display. The probe (250 ms in duration) was either acomplete disk overlapping a square or a notched disk standingseparate from the square (Fig. 1). Both the diameter <strong>of</strong> the diskand one side <strong>of</strong> the square subtended 1.21 <strong>of</strong> visual angle.Squares were red and disks green, presented on a black background.The probe and prime were both presented in the upperleft or upper right quadrant, with the center <strong>of</strong> the disk 21 fromfixation. There were four different stimulus orientations (disk attop left, top right, bottom left, or bottom right <strong>of</strong> square), thusproviding variability in the stimulus. A trailing fixation displaywas presented for either 2,150 or 2,000 ms, depending on theexposure duration <strong>of</strong> the prime (constraining the duration <strong>of</strong>each trial to be 3,000 ms).Null trials were included in addition to the four experimentalconditions to create between-trial temporal jitter (Kourtzi &Kanwisher, 2001). The five trial types were pseudorandomlyintermixed such that each trial type was preceded and followedequally <strong>of</strong>ten by every other trial type. Approximately 48 trialswere presented for each condition in each quadrant. The subject’stask, designed to enhance attention to the stimuli, wasto indicate whether the second stimulus (the probe) contained acomplete or notched disk, by pressing one <strong>of</strong> two buttons <strong>of</strong> aresponse pad.For each subject, event-related time courses (expressed aspercentage signal change) for each experimental condition werecalculated in independently defined ROIs, using the activity atevent onset (time 0) as baseline (Slotnick & Schacter, 2004).Event-related activity was interpolated, yielding 1-s temporalresolution. ROIs in the left and right hemispheres were used toobtain event-related averages corresponding to stimuli in theright and left hemifields, respectively. There was no qualitativedifference in data from the two hemispheres; therefore, wecollapsed data from the two hemispheres <strong>of</strong> each subject andcomputed group mean time courses.RESULTSEarly visual areas (V1, V2, VP, V4v, and LOC) were localizedseparately for each subject and served as ROIs for the analysis <strong>of</strong>360 Volume 17—Number 4