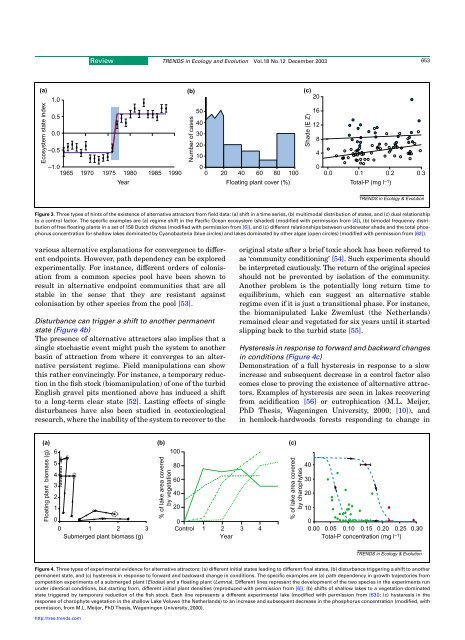
Review TRENDS <strong>in</strong> Ecology and Evolution Vol.18 No.12 December 2003 653(a) (b) (c)201.050160.540120.030208–0.5–1.0100401965 1970 1975 1980 1985 1990 0 20 40 60 80 100Ecosystem state <strong>in</strong>dexNumber of casesYearFloat<strong>in</strong>g plant cover (%)Shade (E Z)0.0 0.1 0.2Total-P (mg l –1 )0.3TRENDS <strong>in</strong> Ecology & EvolutionFigure 3. Three types of h<strong>in</strong>ts of the existence of alternative attrac<strong>to</strong>rs from field data: (a) shift <strong>in</strong> a time series, (b) multimodal distribution of states, and (c) dual relationship<strong>to</strong> a control fac<strong>to</strong>r. The specific examples are (a) <strong>regime</strong> shift <strong>in</strong> the Pacific Ocean ecosystem (shaded) (modified with permission from [4]), (b) bimodal frequency distributionof free float<strong>in</strong>g plants <strong>in</strong> a set of 158 Dutch ditches (modified with permission from [6]), and (c) different relationships between underwater shade and the <strong>to</strong>tal phosphorusconcentration for shallow lakes dom<strong>in</strong>ated by Cyanobacteria (blue circles) and lakes dom<strong>in</strong>ated by other algae (open circles) (modified with permission from [62]).various alternative explanations for convergence <strong>to</strong> differentendpo<strong>in</strong>ts. However, path dependency can be exploredexperimentally. For <strong>in</strong>stance, different orders of colonisationfrom a common species pool have been shown <strong>to</strong>result <strong>in</strong> alternative endpo<strong>in</strong>t communities that are allstable <strong>in</strong> the sense that they are resistant aga<strong>in</strong>stcolonisation by other species from the pool [53].Disturbance can trigger a shift <strong>to</strong> another permanentstate (Figure 4b)The presence of alternative attrac<strong>to</strong>rs also implies that as<strong>in</strong>gle s<strong>to</strong>chastic event might push the system <strong>to</strong> anotherbas<strong>in</strong> of attraction from where it converges <strong>to</strong> an alternativepersistent <strong>regime</strong>. Field manipulations can showthis rather conv<strong>in</strong>c<strong>in</strong>gly. For <strong>in</strong>stance, a temporary reduction<strong>in</strong> the fish s<strong>to</strong>ck (biomanipulation) of one of the turbidEnglish gravel pits mentioned above has <strong>in</strong>duced a shift<strong>to</strong> a long-term clear state [52]. Last<strong>in</strong>g effects of s<strong>in</strong>gledisturbances have also been studied <strong>in</strong> eco<strong>to</strong>xicologicalresearch, where the <strong>in</strong>ability of the system <strong>to</strong> recover <strong>to</strong> theorig<strong>in</strong>al state after a brief <strong>to</strong>xic shock has been referred <strong>to</strong>as ‘community condition<strong>in</strong>g’ [54]. Such experiments shouldbe <strong>in</strong>terpreted cautiously. The return of the orig<strong>in</strong>al speciesshould not be prevented by isolation of the community.Another problem is the potentially long return time <strong>to</strong>equilibrium, which can suggest an alternative stable<strong>regime</strong> even if it is just a transitional phase. For <strong>in</strong>stance,the biomanipulated Lake Zwemlust (the Netherlands)rema<strong>in</strong>ed clear and vegetated for six years until it startedslipp<strong>in</strong>g back <strong>to</strong> the turbid state [55].Hysteresis <strong>in</strong> response <strong>to</strong> forward and backward changes<strong>in</strong> conditions (Figure 4c)Demonstration of a full hysteresis <strong>in</strong> response <strong>to</strong> a slow<strong>in</strong>crease and subsequent decrease <strong>in</strong> a control fac<strong>to</strong>r alsocomes close <strong>to</strong> prov<strong>in</strong>g the existence of alternative attrac<strong>to</strong>rs.Examples of hysteresis are seen <strong>in</strong> lakes recover<strong>in</strong>gfrom acidification [56] or eutrophication (M.L. Meijer,PhD Thesis, Wagen<strong>in</strong>gen University, 2000; [10]), and<strong>in</strong> hemlock-hardwoods forests respond<strong>in</strong>g <strong>to</strong> change <strong>in</strong>(a) (b) (c)6100Float<strong>in</strong>g plant biomass (g)54321% of lake area coveredby vegetation8060402000 00 1 2 3 Control 1 2 3 40.00 0.05 0.10 0.15 0.20 0.25 0.30Submerged plant biomass (g) YearTotal-P concentration (mg l –1 )% of lake area coveredby charophytes40302010TRENDS <strong>in</strong> Ecology & EvolutionFigure 4. Three types of experimental evidence for alternative attrac<strong>to</strong>rs: (a) different <strong>in</strong>itial states lead<strong>in</strong>g <strong>to</strong> different f<strong>in</strong>al states, (b) disturbance trigger<strong>in</strong>g a shift <strong>to</strong> anotherpermanent state, and (c) hysteresis <strong>in</strong> response <strong>to</strong> forward and backward change <strong>in</strong> conditions. The specific examples are (a) path dependency <strong>in</strong> growth trajec<strong>to</strong>ries fromcompetition experiments of a submerged plant (Elodea) and a float<strong>in</strong>g plant (Lemna). Different l<strong>in</strong>es represent the development of the two species <strong>in</strong> the experiments rununder identical conditions, but start<strong>in</strong>g from, different <strong>in</strong>itial plant densities (reproduced with permission from [6]); (b) <strong>shifts</strong> of shallow lakes <strong>to</strong> a vegetation-dom<strong>in</strong>atedstate triggered by temporary reduction of the fish s<strong>to</strong>ck. Each l<strong>in</strong>e represents a different experimental lake (modified with permission from [63]); (c) hysteresis <strong>in</strong> theresponse of charophyte vegetation <strong>in</strong> the shallow Lake Veluwe (the Netherlands) <strong>to</strong> an <strong>in</strong>crease and subsequent decrease <strong>in</strong> the phosphorus concentration (modified, withpermission, from M.L. Meijer, PhD Thesis, Wagen<strong>in</strong>gen University, 2000).http://tree.trends.com
654Review TRENDS <strong>in</strong> Ecology and Evolution Vol.18 No.12 December 2003disturbance <strong>in</strong>tensity [28]. However, a hysteretic patternmight not <strong>in</strong>dicate alternative attrac<strong>to</strong>rs if the response ofthe system is not fast enough relative <strong>to</strong> the rate of change<strong>in</strong> the control fac<strong>to</strong>r. Indeed, one will always see somehysteresis-like pattern unless the system response is muchfaster than the change <strong>in</strong> the control variable.In conclusion, experiments can be a powerful way <strong>to</strong>show that a system has alternative attrac<strong>to</strong>rs. There areobvious limitations <strong>to</strong> explor<strong>in</strong>g large spatial scales andlong time spans. However, small and fast systems, whichare more tractable experimentally, can help us <strong>to</strong> <strong>in</strong>ferwhat might happen <strong>in</strong> larger and slower ones.The contribution of modelsIn addition <strong>to</strong> field data and experiments, models rema<strong>in</strong>a central <strong>in</strong>gredient of most of the work on alternativeattrac<strong>to</strong>rs. Discussion of the long his<strong>to</strong>ry of confusion overthe question of what models can tell us exactly is beyondthe scope of this review, but we highlight some crucialpo<strong>in</strong>ts of <strong>in</strong>terest.Qualitative reason<strong>in</strong>gAlternative attrac<strong>to</strong>rs require positive feedbacks, andchange among attrac<strong>to</strong>rs implies a shift <strong>in</strong> dom<strong>in</strong>antcontroll<strong>in</strong>g feedbacks. For <strong>in</strong>stance, <strong>in</strong> stratified lakes, aclear-water state prevents anoxic phosphorus recycl<strong>in</strong>gfrom the sediments, which, <strong>in</strong> turn, limits phy<strong>to</strong>plank<strong>to</strong>ngrowth. By contrast, <strong>in</strong> a turbid state, high algal biomassboosts anoxic phosphate recycl<strong>in</strong>g, promot<strong>in</strong>g more algalgrowth [10]. Positive feedbacks are sometimes <strong>in</strong>terpretedas be<strong>in</strong>g almost synonymous with the possibility of switchesbetween alternative attrac<strong>to</strong>rs [57]. Although this mightseem reasonable <strong>in</strong>tuitively, positive feedbacks only lead <strong>to</strong>alternative attrac<strong>to</strong>rs if they are sufficiently strong. Thus,qualitative reason<strong>in</strong>g is not sufficient <strong>to</strong> <strong>in</strong>fer alternativeattrac<strong>to</strong>rs from observed feedback loops.Simple mathematical modelsMathematical models provide a much more powerful<strong>to</strong>ol than qualitative reason<strong>in</strong>g for show<strong>in</strong>g that certa<strong>in</strong>mechanisms can lead <strong>to</strong> phenomena of <strong>in</strong>terest such asalternative attrac<strong>to</strong>rs. Most models <strong>in</strong> the literature areso-called m<strong>in</strong>imal models that focus on a m<strong>in</strong>imal set ofmechanisms needed <strong>to</strong> produce a certa<strong>in</strong> behaviour [58].Such models have been useful for explor<strong>in</strong>g mechanismsthat are <strong>to</strong>o <strong>in</strong>tricate <strong>to</strong> grasp from common sense alone,and also serve well for explor<strong>in</strong>g the fundamental implicationsof important aspects such as spatial heterogeneity[30,31,33] and environmental fluctuations [16,17,22].Large simulation modelsA drawback of simple models is that they necessarily leaveout many potentially important aspects. This has <strong>in</strong>vokedthe development of much criticised [58] big simulationmodels <strong>in</strong>tended <strong>to</strong> provide quantitative predictions.Although true validation of such predictive models issimply impossible [59], they rema<strong>in</strong> very important. Onesimply cannot do appropriate experiments with largeimportant systems, such as oceans and the atmosphere.In these situations, our best hope is a comb<strong>in</strong>ation ofmulti-<strong>in</strong>terpretable records of past behaviour with‘plausible models’ of the system.In conclusion, different models have different strengthsand weaknesses, and the best strategy is obviously <strong>to</strong> usedifferent models <strong>in</strong> parallel. Each model is a ‘lie’ <strong>in</strong> thesense that it is an imperfect representation of reality.However, if various <strong>in</strong>dependent models co<strong>in</strong>cide <strong>in</strong> predict<strong>in</strong>galternative attrac<strong>to</strong>rs, then one can adopt thephilosophy that ‘the truth is the <strong>in</strong>tersection of <strong>in</strong>dependentlies’ [60].ProspectsClearly, we have advanced much over the past decade <strong>in</strong>bridg<strong>in</strong>g the gap between abstract <strong>theory</strong> and ecosystemdynamics observed <strong>in</strong> the field. Field data are exploredwith more powerful techniques, and models are be<strong>in</strong>gused <strong>to</strong> address implications of important aspects suchas environmental fluctuations and spatial complexity.Because neither field patterns nor models alone can beconclusive, it is also crucial that well-designed experimentsare be<strong>in</strong>g used now <strong>to</strong> demonstrate alternativeattrac<strong>to</strong>rs. Still, although strong cases have been made, itrema<strong>in</strong>s remarkably difficult <strong>to</strong> prove the existence ofalternative attrac<strong>to</strong>rs <strong>in</strong> the field. This is perhaps notsurpris<strong>in</strong>g <strong>in</strong> view of the his<strong>to</strong>rical difficulties <strong>in</strong> demonstrat<strong>in</strong>gthe general importance of virtually all importantmechanisms (e.g. competition, chaos or density dependence)<strong>in</strong> ecology. However, <strong>in</strong> view of the formidableconsequences of alternative attrac<strong>to</strong>rs for conservationBox 4. Outstand<strong>in</strong>g challengesObta<strong>in</strong><strong>in</strong>g better clues from field dataThere is a wealth of ecological time-series and cross-systems datathat rema<strong>in</strong> <strong>to</strong> be explored. F<strong>in</strong>d<strong>in</strong>g h<strong>in</strong>ts of alternative attrac<strong>to</strong>rsrequires the use of novel techniques [18]. Perhaps even morechalleng<strong>in</strong>g is the question of how we can obta<strong>in</strong> early warn<strong>in</strong>gsignals of <strong>regime</strong> <strong>shifts</strong> from time-series data [18]. Foresee<strong>in</strong>g <strong>shifts</strong>is probably very difficult [1], but it has been suggested that subtlechanges <strong>in</strong> the fluctuation pattern such as ‘redden<strong>in</strong>g’ can serve as aclue of an upcom<strong>in</strong>g collapse [70].Build<strong>in</strong>g more realism <strong>in</strong><strong>to</strong> simple modelsTheory has so far been restricted largely <strong>to</strong> the easiest case of afew species <strong>in</strong>teract<strong>in</strong>g <strong>in</strong> a spatially homogeneous (‘well mixed’)and constant environment. It seems important <strong>to</strong> pursue crucialaspects that have been largely neglected such as spatial heterogeneity[30,31,33], seasonality [31,36] and environmental fluctuations[16,17,21,22]. Also, empirical relationships can be l<strong>in</strong>ked <strong>in</strong><strong>to</strong> simplemodels <strong>to</strong> give transparent quantitative predictions.Perform<strong>in</strong>g crucial experimentsControlled experiments are obviously the most powerful way <strong>to</strong>demonstrate alternative attrac<strong>to</strong>rs and <strong>to</strong> reveal underly<strong>in</strong>g mechanisms.A real challenge here lies <strong>in</strong> experiments on realistic spatialand temporal scales [71]. Small-scale experiments can be veryhelpful <strong>in</strong> expla<strong>in</strong><strong>in</strong>g large-scale patterns [6,47], but this has beenpoorly explored.Design<strong>in</strong>g smart res<strong>to</strong>ration strategiesInsight <strong>in</strong><strong>to</strong> alternative attrac<strong>to</strong>rs can be used <strong>to</strong> design unconventionalres<strong>to</strong>ration methods [72,73]. In particular, the idea of tak<strong>in</strong>gadvantage of climatic extremes such as El Niño events <strong>to</strong> <strong>in</strong>duce ashift <strong>to</strong> a desired <strong>regime</strong> [27] is an excit<strong>in</strong>g, novel development.http://tree.trends.com