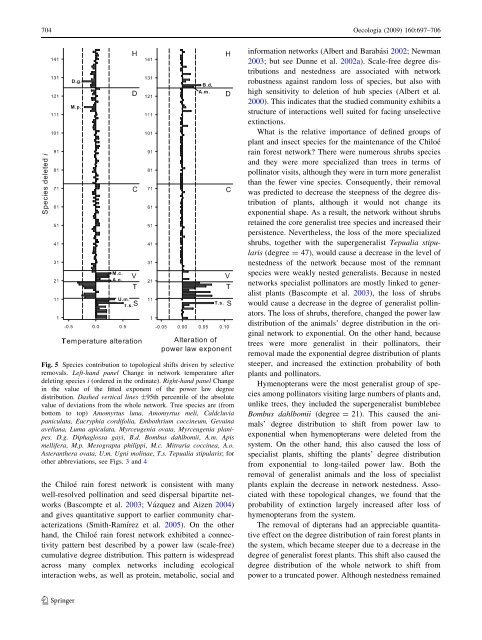
704 Oecologia (2009) 160:697–706Fig. 5 Species contribution to topological shifts driven by selectiveremovals. Left-hand panel Change <strong>in</strong> <strong>network</strong> temperature afterdelet<strong>in</strong>g species i (ordered <strong>in</strong> the ord<strong>in</strong>ate). Right-hand panel Change<strong>in</strong> the value <strong>of</strong> the fitted exponent <strong>of</strong> the power law degreedistribution. Dashed vertical l<strong>in</strong>es ±95th percentile <strong>of</strong> the absolutevalue <strong>of</strong> deviations from the whole <strong>network</strong>. Tree species are (frombottom to top) Amomyrtus luna, Amomyrtus meli, Caldcluviapaniculata, Eucryphia cordifolia, Embothrium cocc<strong>in</strong>eum, Gevu<strong>in</strong>aavellana, Luma apiculata, Myrceugenia ovata, Myrceugenia planipes.D.g. Diphaglossa gayi, B.d. Bombus dahlbomii, A.m. Apismellifera, M.p. Mesograpta philippi, M.c. Mitraria cocc<strong>in</strong>ea, A.o.Asteranthera ovata, U.m. Ugni mol<strong>in</strong>ae, T.s. Tepualia stipularis; forother abbreviations, see Figs. 3 and 4the Chiloé ra<strong>in</strong> forest <strong>network</strong> is consistent with manywell-resolved poll<strong>in</strong>ation and seed dispersal bipartite <strong>network</strong>s(Bascompte et al. 2003; Vázquez and Aizen 2004)and gives quantitative support to earlier community characterizations(Smith-Ramírez et al. 2005). On the otherhand, the Chiloé ra<strong>in</strong> forest <strong>network</strong> exhibited a connectivitypattern best described by a power law (scale-free)cumulative degree distribution. This pattern is widespreadacross many complex <strong>network</strong>s <strong>in</strong>clud<strong>in</strong>g ecological<strong>in</strong>teraction webs, as well as prote<strong>in</strong>, metabolic, social and<strong>in</strong>formation <strong>network</strong>s (Albert and Barabási 2002; Newman2003; but see Dunne et al. 2002a). Scale-free degree distributionsand nestedness are associated with <strong>network</strong>robustness aga<strong>in</strong>st random loss <strong>of</strong> species, but also withhigh sensitivity to deletion <strong>of</strong> hub species (Albert et al.2000). This <strong>in</strong>dicates that the studied community exhibits astructure <strong>of</strong> <strong><strong>in</strong>teractions</strong> well suited for fac<strong>in</strong>g unselectiveext<strong>in</strong>ctions.What is the relative importance <strong>of</strong> def<strong>in</strong>ed groups <strong>of</strong>plant and <strong>in</strong>sect species for the ma<strong>in</strong>tenance <strong>of</strong> the Chiloéra<strong>in</strong> forest <strong>network</strong>? There were numerous shrubs speciesand they were more specialized than trees <strong>in</strong> terms <strong>of</strong>poll<strong>in</strong>ator visits, although they were <strong>in</strong> turn more generalistthan the fewer v<strong>in</strong>e species. Consequently, their removalwas predicted to decrease the steepness <strong>of</strong> the degree distribution<strong>of</strong> plants, although it would not change itsexponential shape. As a result, the <strong>network</strong> without shrubsreta<strong>in</strong>ed the core generalist tree species and <strong>in</strong>creased theirpersistence. Nevertheless, the loss <strong>of</strong> the more specializedshrubs, together with the supergeneralist Tepualia stipularis(degree = 47), would cause a decrease <strong>in</strong> the level <strong>of</strong>nestedness <strong>of</strong> the <strong>network</strong> because most <strong>of</strong> the remnantspecies were weakly nested generalists. Because <strong>in</strong> nested<strong>network</strong>s specialist poll<strong>in</strong>ators are mostly l<strong>in</strong>ked to generalistplants (Bascompte et al. 2003), the loss <strong>of</strong> shrubswould cause a decrease <strong>in</strong> the degree <strong>of</strong> generalist poll<strong>in</strong>ators.The loss <strong>of</strong> shrubs, therefore, changed the power lawdistribution <strong>of</strong> the animals’ degree distribution <strong>in</strong> the orig<strong>in</strong>al<strong>network</strong> to exponential. On the other hand, becausetrees were more generalist <strong>in</strong> their poll<strong>in</strong>ators, theirremoval made the exponential degree distribution <strong>of</strong> plantssteeper, and <strong>in</strong>creased the ext<strong>in</strong>ction probability <strong>of</strong> bothplants and poll<strong>in</strong>ators.Hymenopterans were the most generalist group <strong>of</strong> speciesamong poll<strong>in</strong>ators visit<strong>in</strong>g large numbers <strong>of</strong> plants and,unlike trees, they <strong>in</strong>cluded the supergeneralist bumblebeeBombus dahlbomii (degree = 21). This caused the animals’degree distribution to shift from power law toexponential when hymenopterans were deleted from thesystem. On the other hand, this also caused the loss <strong>of</strong>specialist plants, shift<strong>in</strong>g the plants’ degree distributionfrom exponential to long-tailed power law. Both theremoval <strong>of</strong> generalist animals and the loss <strong>of</strong> specialistplants expla<strong>in</strong> the decrease <strong>in</strong> <strong>network</strong> nestedness. Associatedwith these topological changes, we found that theprobability <strong>of</strong> ext<strong>in</strong>ction largely <strong>in</strong>creased after loss <strong>of</strong>hymenopterans from the system.The removal <strong>of</strong> dipterans had an appreciable quantitativeeffect on the degree distribution <strong>of</strong> ra<strong>in</strong> forest plants <strong>in</strong>the system, which became steeper due to a decrease <strong>in</strong> thedegree <strong>of</strong> generalist forest plants. This shift also caused thedegree distribution <strong>of</strong> the whole <strong>network</strong> to shift frompower to a truncated power. Although nestedness rema<strong>in</strong>ed123
Oecologia (2009) 160:697–706 705unchanged after delet<strong>in</strong>g flies, the dynamic output didchange. At high ext<strong>in</strong>ction/colonization rates, remov<strong>in</strong>gdipterans did not differ from random poll<strong>in</strong>ator speciesdeletion. At low and moderate k, species ext<strong>in</strong>ctions werelower after remov<strong>in</strong>g dipterans compared to randomremoval, which is expla<strong>in</strong>ed by the strong effect <strong>of</strong>hymenopterans. Hence, random hymenopteran removalwould constitute a more severe perturbation than dipteranremoval. The exception was found for plants at k = 0.25,where deletion <strong>of</strong> dipterans predicted the ext<strong>in</strong>ction <strong>of</strong> twoplant species (Gaultheria mucronata and Myrteola nummularia)that depend exclusively on them.Remov<strong>in</strong>g coleopterans from the web exerted only amarg<strong>in</strong>al effect on <strong>network</strong> persistence, because ext<strong>in</strong>ction<strong>of</strong> poll<strong>in</strong>ators was entirely expla<strong>in</strong>ed by decreased animalspecies richness. Plant ext<strong>in</strong>ction probability was lowerwhen remov<strong>in</strong>g beetles relative to random species loss, andnot different from the dynamics <strong>of</strong> the unmanipulated<strong>network</strong> (except at k = 0.75). Removal <strong>of</strong> coleopterans didnot change the shape <strong>of</strong> the degree distributions, either forthe whole <strong>network</strong> or for plants and animals separately.The same occurred when v<strong>in</strong>es, mostly related to specializedvisitors, were removed from the mutualistic web.The effect <strong>of</strong> v<strong>in</strong>e removal on ext<strong>in</strong>ction probability canbe considered negligible at low and moderate k because <strong>of</strong>the low number <strong>of</strong> ext<strong>in</strong>ct species (mean = 0.99 plantspecies, range 0–3). At k = 0.75, random removal exerteda more severe effect on species persistence than removal <strong>of</strong>v<strong>in</strong>es due to the greater relative weight <strong>of</strong> trees.Overall, this work highlights the importance <strong>of</strong> flower<strong>in</strong>gcanopy trees and hymenopterans as the core groups thatma<strong>in</strong>ta<strong>in</strong> the species richness, structure and dynamics <strong>of</strong>the Chiloé ra<strong>in</strong> forest poll<strong>in</strong>ation <strong>network</strong>. This wasrevealed by the changes detected <strong>in</strong> topological anddynamic features <strong>of</strong> the ra<strong>in</strong> forest <strong>network</strong> after model<strong>in</strong>gthe removal <strong>of</strong> each group <strong>of</strong> species. Furthermore, discern<strong>in</strong>gwhich species are mostly responsible for the effectsobserved after removal <strong>of</strong> multi-species groups can be <strong>of</strong>great value from a conservation perspective, becauseresearch, conservation and management efforts can bedirected to few pivotal species <strong>in</strong> cases where protect<strong>in</strong>gthe entire community is not possible. By assess<strong>in</strong>g thesensitivity <strong>of</strong> degree distribution and nestedness to theremoval <strong>of</strong> <strong>in</strong>dividual nodes we obta<strong>in</strong>ed a measure <strong>of</strong>the relative contribution <strong>of</strong> each species to the ma<strong>in</strong>tenance<strong>of</strong> community structure and diversity. In this way, weidentified a number <strong>of</strong> species that deserve special attentionfor the conservation <strong>of</strong> the entire species assemblage. Thehymenopterans Bombus dahlbomii and Diphaglossa gayi,the shrubs Tepualia stipularis and Ugni mol<strong>in</strong>ae, the v<strong>in</strong>esMitraria cocc<strong>in</strong>ea and Asteranthera ovata, and the wholeset <strong>of</strong> trees were predicted to exert a disproportionatelylarge <strong>in</strong>fluence on the preservation <strong>of</strong> the structure <strong>of</strong> the<strong>network</strong> under study. Their ext<strong>in</strong>ction would seriouslyharm the reproductive success <strong>of</strong> plants <strong>in</strong> this <strong>temperate</strong>ra<strong>in</strong> forest. Some tree species with the richest sets <strong>of</strong>poll<strong>in</strong>ators, such as the emergent Eucryphia cordilfolia, arecurrently under severe and rapid decl<strong>in</strong>e due to selectivelogg<strong>in</strong>g and land clear<strong>in</strong>g, and their loss may promotecascade ext<strong>in</strong>ctions <strong>of</strong> <strong>network</strong> participants (Armesto et al.1996). In particular, the woody species Tepualia stipularisand Eucryphia cordifolia were shown <strong>in</strong> this study to playan important structural role with<strong>in</strong> the Chiloé <strong>network</strong> andthey are also relevant for habitat provision (Díaz et al.2005). Unfortunately, they both are highly threatenedspecies <strong>in</strong> Chiloé Island, represent<strong>in</strong>g respectively 55 and11% <strong>of</strong> the total commercialized firewood (unpublisheddata).It is accepted that the stability and robustness <strong>of</strong> poll<strong>in</strong>ation<strong>network</strong>s to species loss are sensitive to structuralattributes <strong>of</strong> the <strong>network</strong>. More specifically, nestedness andlong-tailed degree distribution have been postulated as keytopological properties that prevent species loss (Memmottet al. 2004; Fortuna and Bascompte 2006; Bascompte andJordano 2007). Here we explored the possible <strong>in</strong>terplaybetween structure and dynamics <strong>of</strong> a poll<strong>in</strong>ation systemthat supports a highly valued and remarkably endangeredforest ecosystem. A better understand<strong>in</strong>g <strong>of</strong> the function<strong>in</strong>gand evolution <strong>of</strong> target ecological systems would be ga<strong>in</strong>edif further efforts were made to ensure that most <strong>of</strong> theparticipant species were recorded. In addition, it is necessaryto identify them at the best level <strong>of</strong> taxonomic resolution,record<strong>in</strong>g their temporal and spatial variation, and todevelop better analytical tools to manage, organize andtake advantage <strong>of</strong> the data.Acknowledgements The authors thank J. D. Flores for his valuablehelp <strong>in</strong> scientific comput<strong>in</strong>g, M. A. Fortuna for his help with thedynamic model, M. A. Rodríguez-Gironés for specially implement<strong>in</strong>ga batch version <strong>of</strong> his s<strong>of</strong>tware BINMATNEST, and M. Franco forimprov<strong>in</strong>g the readability <strong>of</strong> this paper. All the experiments andsampl<strong>in</strong>g comply with the current laws <strong>of</strong> the country (Chile) <strong>in</strong>which they were performed. Work started by an Endowed PresidentialChair <strong>in</strong> Sciences to J. J. A. We acknowledge support from FON-DAP–FONDECYT 1501-0001, P05-002 ICM and PFB-23 CONI-CYT, Chile. This is a contribution to the research program <strong>of</strong> SendaDarw<strong>in</strong> Biological Station, Chiloé, Chile. We appreciate the companyand assistance <strong>in</strong> the field <strong>of</strong> M. Nuñez-Ávila, L. Suárez, P. Martínezand el Negro.ReferencesAizen MA, Ezcurra C (1998) High <strong>in</strong>cidence <strong>of</strong> plant-animalmutualisms <strong>in</strong> the woody flora <strong>of</strong> the <strong>temperate</strong> forest <strong>of</strong>southern South America: biogeographical orig<strong>in</strong> and presentecological significance. Ecol Aust (Argent<strong>in</strong>a) 8:217–236Aizen MA, Vasquez DP, Smith-Ramírez C (2002) Historia natural yconservación de los mutualismos planta-animal del bosquetemplado de Sudamérica austral. Rev Chil Hist Nat 75:79–97123