

Function of the chemokine receptor CXCR4 in haematopoiesis and ...
Function of the chemokine receptor CXCR4 in haematopoiesis and ...
Function of the chemokine receptor CXCR4 in haematopoiesis and ...

You also want an ePaper? Increase the reach of your titles
YUMPU automatically turns print PDFs into web optimized ePapers that Google loves.
letters to nature<br />
result from deficiency <strong>in</strong> commitment <strong>and</strong>/or proliferation <strong>of</strong> Bprogenitor<br />
cells.<br />
<strong>CXCR4</strong> is highly expressed <strong>in</strong> <strong>the</strong> thymus, particularly <strong>in</strong> immature<br />
CD4 + CD8 + cells 4 . However, T lymphopoiesis occurred normally<br />
<strong>in</strong> <strong>the</strong> mutant embryos at E17.5 (not shown). At this stage,<br />
most thymocytes are immature CD4 + CD8 + cells; few CD4 + CD8 − or<br />
CD4 − CD8 + mature T lymphocytes are present, <strong>and</strong> <strong>the</strong>re is little or<br />
no emigration <strong>of</strong> thymocytes to <strong>the</strong> peripheral lymphoid organs.<br />
Emigration commences only after birth through a process <strong>in</strong> which<br />
<strong>the</strong> Gi prote<strong>in</strong>s are important 14,15 . As <strong>CXCR4</strong> expressed <strong>in</strong> rest<strong>in</strong>g T<br />
cells is coupled to G�i2 (refs 4, 10) <strong>and</strong> as mature T cells can migrate<br />
<strong>in</strong> response to SDF-1 stimulation 9 , migration <strong>and</strong> recirculation <strong>of</strong><br />
mature T cells might require signals delivered through <strong>CXCR4</strong>. To<br />
determ<strong>in</strong>e whe<strong>the</strong>r maturation <strong>and</strong> emigration <strong>of</strong> thymocytes<br />
requires <strong>CXCR4</strong>, we engrafted <strong>the</strong> thymuses from E17.5 <strong>CXCR4</strong> −/−<br />
embryos under <strong>the</strong> kidney capsule <strong>of</strong> mice lack<strong>in</strong>g T-cell antigen<br />
<strong>receptor</strong> (TCR)-�. As <strong>the</strong> recipient mice are deficient <strong>in</strong> ��-T-cell<br />
development, any TCR-�� T cells detected <strong>in</strong> <strong>the</strong> periphery are<br />
derived from <strong>the</strong> donor thymus. Our results <strong>in</strong>dicate that mature<br />
CD4 + CD8 − <strong>and</strong> CD4 − CD8 + T cells developed normally <strong>in</strong> <strong>the</strong><br />
implanted thymuses. Moreover, <strong>the</strong>se T cells efficiently populated<br />
<strong>the</strong> peripheral lymphoid organs, <strong>in</strong>clud<strong>in</strong>g peripheral blood<br />
8<br />
(Fig. 2d), spleen, <strong>and</strong> mesenteric lymph nodes (results not<br />
shown). Thus, <strong>CXCR4</strong> has a negligible effect on thymocyte maturation<br />
<strong>and</strong> subsequent migration to lymphoid organs.<br />
As <strong>CXCR4</strong> messenger RNA is expressed <strong>in</strong> bra<strong>in</strong> 4 , we used <strong>in</strong> situ<br />
hybridization to study <strong>the</strong> embryonic mouse bra<strong>in</strong> at E13, E15 <strong>and</strong><br />
E18. <strong>CXCR4</strong> mRNA transcripts were found <strong>in</strong> many regions <strong>of</strong> <strong>the</strong><br />
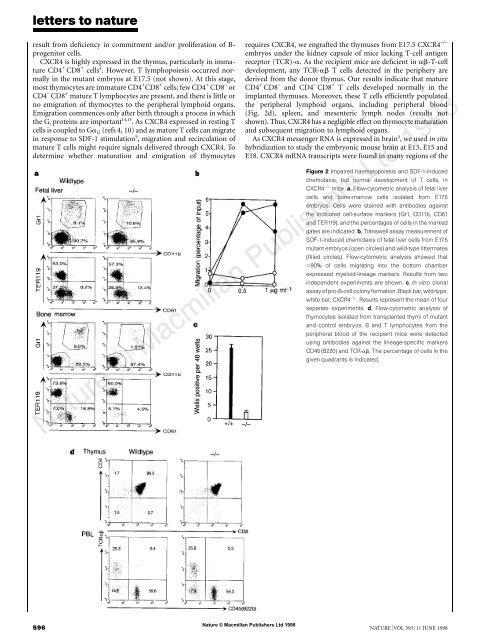
Figure 2 Impaired <strong>haematopoiesis</strong> <strong>and</strong> SDF-1-<strong>in</strong>duced<br />
chemotaxis, but normal development <strong>of</strong> T cells, <strong>in</strong><br />
<strong>CXCR4</strong> −/− mice. a, Flow-cytometric analysis <strong>of</strong> fetal liver<br />
cells <strong>and</strong> bone-marrow cells isolated from E17.5<br />
embryos. Cells were sta<strong>in</strong>ed with antibodies aga<strong>in</strong>st<br />
<strong>the</strong> <strong>in</strong>dicated cell-surface markers (Gr1, CD11b, CD61<br />
<strong>and</strong> TER119), <strong>and</strong> <strong>the</strong> percentages <strong>of</strong> cells <strong>in</strong> <strong>the</strong> marked<br />
gates are <strong>in</strong>dicated. b, Transwell assay measurement <strong>of</strong><br />
SDF-1-<strong>in</strong>duced chemotaxis <strong>of</strong> fetal liver cells from E17.5<br />
mutant embryos (open circles) <strong>and</strong> wild-type littermates<br />
(filled circles). Flow-cytometric analysis showed that<br />
�90% <strong>of</strong> cells migrat<strong>in</strong>g <strong>in</strong>to <strong>the</strong> bottom chamber<br />
expressed myeloid-l<strong>in</strong>eage markers. Results from two<br />
<strong>in</strong>dependent experiments are shown. c, In vitro clonal<br />
assay <strong>of</strong> pro-B-cell colony formation. Black bar, wild-type;<br />
white bar, <strong>CXCR4</strong> −/− . Results represent <strong>the</strong> mean <strong>of</strong> four<br />
separate experiments. d, Flow-cytometric analysis <strong>of</strong><br />
thymocytes isolated from transplanted thymi <strong>of</strong> mutant<br />
<strong>and</strong> control embryos. B <strong>and</strong> T lymphocytes from <strong>the</strong><br />
peripheral blood <strong>of</strong> <strong>the</strong> recipient mice were detected<br />
us<strong>in</strong>g antibodies aga<strong>in</strong>st <strong>the</strong> l<strong>in</strong>eage-specific markers<br />
CD45 (B220) <strong>and</strong> TCR-��. The percentage <strong>of</strong> cells <strong>in</strong> <strong>the</strong><br />
given quadrants is <strong>in</strong>dicated.<br />
Nature © Macmillan Publishers Ltd 1998<br />
596 NATURE | VOL 393 | 11 JUNE 1998


