Create successful ePaper yourself
Turn your PDF publications into a flip-book with our unique Google optimized e-Paper software.
Página1<br />
<strong>TEJIDO</strong> <strong>NERVIOSO</strong><br />
El tejido nervioso es un tejido cuya principal función es la integración anatomofisiológica de nuestro<br />
organismo a través de unas células especializadas que proceden del ectodermo y que progresivamente se<br />
diferencian en una familia de poblaciones celulares que reparten sus funciones desde el principio de modo que<br />
hay células con capacidad excitatoria llamadas neuronas y células de apoyo llamadas células gliales.<br />
Historia<br />
El descubrimiento de estas células arranca en el siglo XIX cuando a partir de la década de 1850 los<br />
microscopios incrementan su resolución para conocer el SN, por su arquitectura celular especial formada por<br />
una especie de coto cerrado en la que fisiológicamente la circulación sanguínea es muy densa, no hay<br />
circulación linfática ni de linfocitos y donde las células parenquimales adoptan una morfología reticular.<br />
El aspecto reticular fue lo que más llamó la atención en el siglo XIX cuando los investigadores descubrieron<br />
que el tejido que acoge los procesos superiores mentales y afectivos está formado por una <strong>com</strong>plicada red. A<br />
partir de este punto aparecen dos corrientes para conocer el SN:<br />
Células con principio y final que interactúan y se organizan en red<br />
Retículo formado por células sin diferenciación entre ellas<br />
Fue Ramón y Cajal quien consiguió, mediante técnicas muy <strong>com</strong>plejas, demostrar el inicio y fin de cada<br />
neurona para explicar la histología del SN.<br />
Durante la primera mitad del siglo XX ha habido un gran desarrollo en el conocimiento de este tejido,<br />
gracias también a la ME. La incorporación de los Ac monoclonales ha permitido conocer todas las sustancias de<br />
señalización y adhesión del tejido nervioso.<br />
En el siglo XXI se plantea el estudio de biomarcadores que permitan conocer la patología del SN y de la<br />
conducta.<br />
NEURONA<br />
La neurona se puede dividir en un cuerpo o soma neuronal:<br />
Cuerpo o Soma Neuronal<br />
Se localiza el citoplasma principal que contiene un núcleo especial de gran tamaño, nucléolo marcado y<br />
central que denota la gran diferenciación de la célula. La célula está detenida en la fase G 0, por lo que no puede<br />
dividirse, excepto neuronas bipolares de la mucosa olfatoria y otras neuronas en diversos puntos del organismo<br />
que se pueden dividir.<br />
En el soma se almacenan pigmentos citoplasmáticos de lipofuscina, melanina y de gránulos<br />
neurosecretores. Esto da una coloración natural del citoplasma que puede teñirse para diferenciarlas. Dentro de<br />
los gránulos destacan los cuerpos de Niss, que contienen una sustancia cromatófila; son vesículas aplanadas de<br />
REL mezcladas con polirribosomas y que están presentes en el cuerpo celular y las dendritas; son basófilas.<br />
Destaca también el aparato de Golgi, que organiza a nivel subcelular la secreción de moléculas que se<br />
liberarán por sus prolongaciones. En el aparato de síntesis molecular también intervienen el RER y el REL; el<br />
conjunto de estos tres <strong>com</strong>ponentes está muy desarrollado en las neuronas.
Página2<br />
Los lisosomas primarios contienen enzimas proteolíticas de acción intra o extracelular. Las mitocondrias se<br />
encuentran en un número alto, ya que permiten el aporte energético de la neurona, su excitabilidad y el flujo de<br />
moléculas; este gran gasto supone un gran estrés oxidativo a la célula.<br />
El citoesqueleto ya fue teñido en el siglo XIX y está formado por haces de neurofibrillas que pueden<br />
alcanzar hasta 3μm de grosor, lo que los hace muy visibles. Estas neurofibrillas dan soporte a la célula. Hay<br />
varios <strong>com</strong>ponentes:<br />
Neurofilamentos que son muy largas y permiten mantener la estructura de la neurona.<br />
Neurotúbulos, de 25nm, mantienen la estructura del axón y su flujo molecular<br />
Prolongaciones Celulares<br />
Las dendritas son procesos celulares muy ramificados que tienen un carácter dinámico, en cuyo interior<br />
hay microtúbulos y microfilamentos y cuya misión es extender la célula hacia territorios adyacentes para tener<br />
una mayor capacidad receptiva. Están regulados por el citoesqueleto y contienen numerosos receptores<br />
funcionales.<br />
Los axones son prolongaciones de longitud y número variable y diámetro constante, que oscila entre 1-<br />
20μm. La longitud depende del tamaño de la neurona. El axón permite transmitir los impulsos desde el soma<br />
neuronal. Se pueden distinguir varios segmentos:<br />
- Cono de salida: punto más proximal del axón<br />
- Segmento inicial<br />
- Tronco del axón: es el más largo<br />
- Ramificaciones colaterales del axón<br />
- Telodendria: ramificaciones más distantes dentro del axón<br />
- Botones terminales: órganos subcelulares en los que se acumulan los neurotransmisores<br />
El axón contiene un citoesqueleto de neurotúbulos y neurofilamentos, aunque también se encuentran<br />
mitocondrias y nunca cuerpos de Niss. En cambio, sí hay REL que tiene capacidad sintética. La membrana<br />
celular axónica o axolema está rodeada por un envoltorio de mielina gracias a las células gliales.<br />
Los axones permiten el transporte molecular de sustancias desde el soma celular hasta territorios alejados;<br />
por esto, el axón tiene un transporte molecular que se conoce <strong>com</strong>o flujo axoplasmático, que es bidireccional:<br />
retrógrado o cerulípedo si va desde la parte distal del axón al soma, pero anterógrado o cerulífugo si va desde el<br />
soma al botón terminal.<br />
El sentido retrógrado permite a la neurona mover moléculas captadas cerca del axón para que los<br />
incorpore la neurona. Permite el transporte de proteínas captadas por el terminal axónico.<br />
El sentido anterógrado permite llevar moléculas neuronales al exterior. Hay dos velocidades:<br />
- Lenta de 1-3mm/día flujo natural de moléculas de forma pasiva desde el soma celular hasta la<br />
periferia y está facilitada por el propio peristaltismo mecánico de la vaina axonal.<br />
- Rápido de 100-500mm/día es independiente de la peristalsis de la vaina, si no que depende del<br />
propio citoesqueleto. Es un transporte muy eficiente que puede darse con varias moléculas desde el<br />
soma al extremo distal del axón.<br />
Por todo esto, el axón permite canalizar moléculas propias para expulsarlas al exterior permitiendo el<br />
trofismo de tejidos externos, pero a la vez puede recaptar moléculas.<br />
Regulación de la Acción Axonal<br />
Existen factores que estimulan el flujo axonal, <strong>com</strong>o los factores neurotróficos desde la época embrionaria.<br />
Los describió en primer lugar Ramón y Cajal en 1909, pero en 1960 se pudieron conocer las primeras<br />
neurotrofinas por Levi-Montalcini. Las neurotrofinas permitían aumentar la motilidad axonal por mecanismos
Página3<br />
quimiotácticos. Esta regulación de la dirección está controlada por quimiocinas, que orientan a los axones, y<br />
moléculas de adhesión, que permite la unión a tejidos periféricos.<br />
Las moléculas neurotróficas regulan el desarrollo y flujo axonal, mientras que la dirección de estos axones<br />
a tejidos periféricos está regulado por las quimiocinas y las moléculas de adhesión.<br />
Tipos de Neurona<br />
La población neuronal es muy heterogénea y en algunas partes del encéfalo existen progenitores<br />
neurogénicos que podrían producir neuronas, <strong>com</strong>o en el hipocampo o zonas subventriculares; en terapia<br />
celular podrían ser un foco para el tratamiento. Los progenitores neurogénicos podrían producir neuroblastos,<br />
aunque el paso a neurona es muy <strong>com</strong>plicado y limitado al desarrollo del encéfalo.<br />
En las etapas del desarrollo neural se pueden establecer 3 estadios:<br />
1. Génesis de neuronas: análogo morfogenéticamente a cualquier otro tejido<br />
2. Crecimiento de axones y dendritas: se dan otros principios diferentes y está regulada por la<br />
morfogénesis específica del encéfalo. Estas permiten formar las conexiones entre diferentes neuronas.<br />
En esta fase son muy importantes los factores que guían a las ramificaciones neuronales en las<br />
diferentes regiones encefálicas.<br />
3. Por señales eléctricas que reciben y transmiten las prolongaciones formadas permiten formar sinapsis<br />
entre las células<br />
Clasificación Morfológica<br />
Debido a la diferenciación hay una clasificación neuronal se pueden clasificar las neuronas según su número de<br />
prolongaciones:<br />
NEURONAS MULTIPOLARES<br />
Son las más frecuentes. Tienen múltiples prolongaciones en todas las direcciones y un axón dirigido hacia la<br />
región efectora. Suelen ser motoneuronas, aunque también hay interneuronas de este tipo. Son:<br />
<br />
<br />
<br />
<br />
<br />
Neuronas en cesto: las dendritas abrazan a otra célula<br />
Neuronas estrelladas: con dendritas en todas las direcciones<br />
Neuronas granulares: sus dendritas se encuentran en todas las direcciones. Se localizan en las capas de<br />
células granulosas del encéfalo (<strong>com</strong>o cerebelo)<br />
Neuronas piramidales<br />
Neuronas de Purkinje: son de un tamaño > a 100µm y se localizan en la capa de células ganglionares
Página4<br />
Dentro de este grupo también se encuentran las células de Golgi I (motoneuronas con muchas dendritas y un<br />
axón muy largo que forma parte de los nervios) y las células de Golgi II (neuronas granulares de corteza<br />
cerebral con pocas dendritas y un corto axón que no sale de la sustancia gris).<br />
NEURONAS UNIPOLARES<br />
Como en el caso de los fotorreceptores, que tienen un único<br />
axón (muy corto, en este caso).<br />
PSEUDOUNIPOLARES<br />
Al nacer tienen dos polos, es decir, su origen es unipolar<br />
pero al diferenciarse trasladan sus dos prolongaciones y se<br />
unen en el extremo proximal y se separan en distal<br />
mediante una bifurcación. Se encuentran en los ganglios<br />
cocleares y espinales sensitivos.<br />
BIPOLARES<br />
Las neuronas bipolares presentan dos axones, <strong>com</strong>o en el caso de las células olfatorias, de la retina y del oído.<br />
Clasificación Funcional<br />
Motoras<br />
Sensoriales<br />
Interneuronas<br />
Clasificación Neuroquímica<br />
Según el neurotransmisor y hormonas. Ha permitido crear la quimioarquitectura cerebral, una cartografía que<br />
indica la distribución de las neuronas en el encéfalo según sus productos de secreción. Tiene una gran<br />
aplicación farmacológica según la estructura cerebral. Sobre esta base se han creado varios tipos de neuronas:<br />
Neuronas colinérgicas liberadoras de Ach. Se marcan con Ac que marcan la acetilcolintransferasa, enzima<br />
necesaria para el metabolismo de la Ach.<br />
Neuronas noradrenérgidas y adrenérgicas, que producen adrenalina y NA. Se les distingue con Ac contra la<br />
feniletanolamilmetiltransferasa<br />
Neuronas serotoninérgicas que liberan 5-HT<br />
Neuronas dopaminérgicas que usan dopamina<br />
Neuronas que usan GABA o neuronas GABA+<br />
Neuronas peptidérgicos que liberan polipéptidos, <strong>com</strong>o la encefalina, neurotensina, CCK,…<br />
Neuronas de receptores opiáceos, que usan péptidos opiáceos <strong>com</strong>o señalización
Página5<br />
Sinapsis<br />
Las sinapsis son contactos intercelulares entre neuronas o con otras células para transmitir la excitación<br />
entre 2 o más células. Se puede clasificar la sinapsis en dos grupos: eléctricas y químicas.<br />
Las sinapsis eléctricas son muy elementales y la estructura intercelular que permite su desarrollo son las<br />
uniones intercelulares de <strong>com</strong>unicación directa o GAP junctions (GAP-J). A través de las GAP-J se da el paso de<br />
moléculas de una neurona a otra de modo que las neuronas se acoplan o sincronizan funcionalmente porque las<br />
GAP-J permiten el flujo intercelular de iones y pequeñas moléculas entre citoplasmas adyacentes. Las GAP-J<br />
están formadas por hexámeros de conexinas que se acoplan entre células vecinas y cuya permeabilidad está<br />
regulada por el Ca y por factores hormonales que regulan la conformación abierta o cerrada del conexón.<br />
Las sinapsis químicas son las que utilizan neurotransmisores para la propagación del mensaje eléctrico<br />
entre las neuronas con otras neuronas u otros tipos celulares <strong>com</strong>o la célula muscular o glandular. Cuando es<br />
entre neuronas los neurotransmisores permiten la despolarización eléctrica entre células contiguas, por lo que<br />
la sinapsis química transforma una señal eléctrica en una señal química que produce una nueva despolarización<br />
en la células post-sináptica, por lo que se vuelve una señal eléctrica.<br />
El proceso consiste en la llegada de la despolarización de membrana hasta el lugar de sinapsis lo que<br />
aumenta el Ca intracelular, lo que conduce a la liberación del neurotransmisor que interactúa con receptores de<br />
la neurona para despolarizar esa célula.<br />
Estructura de la Sinapsis<br />
1. Zona presináptica: botón axónico (0,5-4µm), en el que se distingue:<br />
a. Axolema: estructura <strong>com</strong>pleja de forma hexagonal asociada al retículo endoplasmático para<br />
crear pasillos de transporte de las vesículas. Los espacios <strong>com</strong>prendidos entre las proyecciones<br />
son activos y se conocen <strong>com</strong>o sinaptoporos.<br />
b. Vesículas sinápticas de 40-60nm que contienen a los nucleótidos. Pueden ser esféricas (S), que<br />
son más densas y contienen ACh, o aplanadas (F), menos densas y que contienen GABA y glicina.<br />
c. Hendidura sináptica<br />
d. Neurotransmisor<br />
2. Zona post-sináptica<br />
Tipos de Sinapsis<br />
Axosomática; encontramos dentro de ella varios subtipos:<br />
I. Botones terminales de Held: típico de sinapsis multipolares, <strong>com</strong>o en el asta A de la ME<br />
II. Terminaciones en cáliz: los axones son cortos y abrazan las neuronas aportando múltiples conexiones<br />
III. Entre neuronas granulares y células de Purkinje: típica del cerebelo.<br />
Axodendrítica: si contacta con las espinas neuronales es una conexión axoespínica<br />
Axoaxónica<br />
Dendrodendríticas, dendrosomáticas y somatosomáticas.<br />
También pueden clasificarse en sinapsis tipo I o excitatorias si se despolariza la neurona post-sináptica o tipo II<br />
o inhibitorias si se hiperpolariza la neurona post-sináptica (todos los <strong>com</strong>ponentes de estas sinapsis son de<br />
menor tamaño).
Página6<br />
CÉLULAS DE LA GLÍA<br />
Es la población celular no neuronal del SNC y SNP; en conjunto, representan más del 50% del peso cerebral<br />
y superan a las neuronas en una proporción 10:1 hasta 50:1. En definitiva, las células gliales predominan en el<br />
tejido nervioso.<br />
Son células de pequeño tamaño, de morfología ramificada (<strong>com</strong>o las neuronas), con gran densidad<br />
numérica en algunos tejidos, es decir, pueden acumularse focalmente. Cuando están asociadas a neuronas, se<br />
asocian a sus prolongaciones. Son células que mantienen su capacidad proliferativa. Funcionalmente están<br />
implicadas en el sostén mecánico del SN, regulación de la nutrición neuronal, control del microambiente<br />
químico de la neurona, forman las vainas de mielina y tienen la capacidad defensiva del SN regulando el acceso<br />
del SI al encéfalo a la vez que eliminan los desechos celulares y moleculares que se puedan acumular en el tejido<br />
nervioso. Por último, son las responsables de la regeneración de las fibras nerviosas en SNP y SNC.<br />
Las gliofibrillas forman el citoesqueleto de las células gliales. Se pueden encontrar tanto en el cuerpo<br />
celular <strong>com</strong>o en sus prolongaciones y tienen su máxima cantidad en los astrocitos fibrosos (menor en los<br />
protoplasmáticos). Son neurofilamentos que mantienen la estructura del cuerpo celular y sus prolongaciones.<br />
Químicamente presentan la proteína fibrilar glial acídica o PFGA que es exclusiva de las células gliales (PFGA+);<br />
junto con astrocitos, también hay células estrelladas hepáticas y pancreáticas que también expresan esta<br />
proteína. La pérdida de las células PFGA+ en hígado significa el paso de célula estrellada a célula que sintetiza<br />
matriz extracelular, por lo que en la clínica se usan 2 Ac para observar si no se expresa la PFGA y, en consecuencia,<br />
se está desarrollando una cirrosis hepática.<br />
Las células gliales se diferencian según su localización:<br />
Células Gliales del SNC<br />
Astrocitos<br />
Son las células gliales grandes que tienen numerosas prolongaciones que se pueden clasificar en función a<br />
dónde las dirija, <strong>com</strong>o las prolongaciones perivasculares que rodean a los capilares del encéfalo formando la<br />
membrana glial limitante perivascular.<br />
La membrana glial limitante perivascular es un envoltorio resultante de la acumulación de las<br />
prolongaciones celulares del astrocito en el entorno capilar que impide el paso de metabolito o sustrato<br />
molecular directo desde el capilar a las neuronas, ya que antes debe pasar por los astrocitos. Así, se regula la<br />
disponibilidad de nutrientes según qué deje pasar esta membrana, por lo que se estabiliza el ambiente<br />
adyacente a la neurona y se evita que la neurona esté afectada por las fluctuaciones del plasma.<br />
El astrocito también envía prolongaciones a la superficie encefálica creando la membrana glial limitante<br />
subpial, que se encuentra bajo al piamadre. Es muy importante porque representa un envoltorio citoplasmático<br />
para todo el ambiente neuronal, sobre todo en la corteza celular.<br />
Las prolongaciones perineuronales envuelven a las propias neuronas para proteger a las neuronas<br />
directamente desde el punto de vista físico, químico y metabólico.<br />
Hay varios tipos de astrocitos:<br />
- Astrocitos protoplasmáticos = 15-20%. Se localizan en la materia gris y tienen procesos<br />
citoplasmáticos amplios que envuelven a neuronas y capilares. Algunos de ellos también forman<br />
parte de la membrana glial limitante subpial.<br />
Protoplasmático = similar a sábana<br />
- Astrocitos fibrosos = 80-85%; son la mayoría. Están en la sustancia blanca, son más pequeños (10-<br />
15μm de diámetro), con prolongaciones finas pero con muchas gliofibrillas. Las prolongaciones<br />
envuelven principalmente a los capilares, aunque también a la piamadre y dan soporte metabólico a<br />
las fibras nerviosas.
Página7<br />
- Astrocito plasmofibrilar: se localizan en la frontera entre sustancia blanca y gris. Envían procesos<br />
protoplasmáticos a la sustancia gris y procesos más fibrosos a la sustancia blanca.<br />
- Astrocitos en velo: se localizan en la capa granular del cerebelo. Rodean a ciertas neuronas asociadas<br />
a capilares de esta zona<br />
- Células de Müller: aparecen en la retina<br />
- Célula de Bergman: aparece en el cerebelo<br />
Cumplen muchas funciones:<br />
o Son depredadores iónicos, es decir, captan iones de K y aniones de modo que evitan desestabilizar la<br />
capacidad excitatoria de las neuronas<br />
o Manejo de la glucosa: las células gliales activadas por algunos neurotransmisores estimulan la<br />
glucogenolisis y por tanto se convierten en células liberadoras de glucosa a la neuronaç<br />
o Forman parte de la barrera hemato-encefálica (H-E) porque recubren la superficie de los vasos<br />
o Células muy proliferativas con capacidad cicatricial, cicatrizando lesiones cerebrales; al estudiarlo en<br />
biopsias de tumores cerebrales, proliferan cerca de los tumores formando la respuesta gliar peritumoral<br />
o gliosis peritumoral. El tumor se sobrepone a esta contención glial beneficiándose de las neurotrofinas,<br />
nutrientes y factores de crecimiento gliales para desarrollarse.<br />
Oligodentrocitos<br />
Los oligodentrocitos son células pequeñas de citoplasma oscuro que se localizan en la sustancia gris y<br />
blanca. A pesar de su pequeño diámetro (8μm), tienen un gran núcleo muy marcado y posee muchos procesos<br />
citoplasmáticos cuya función es la formación de cubiertas de mielina durante el proceso de recubrimiento<br />
axonal de la fibras nerviosas del encéfalo.<br />
El oligodentrocito suele tener 8-10 prolongaciones que se dirigen a diferentes puntos, mientras que la<br />
célula de Schwann sólo emite una. Esto lo hace muy versátil para proteger los axones celulares con la mielina.<br />
Cada oligodentrocito puede mielinizar diferentes segmentos de una misma fibra o enviar cada<br />
prolongación a una neurona diferente. Así, un oligodentrocito mediante diferentes procesos citoplasmáticos<br />
puede recubrir la misma fibra nerviosa o recubrir diferentes neuronas.<br />
Hay dos tipos de oligodentrocitos:<br />
- Interfasciculares: sustancia blanca<br />
- Oligodentrocitos perineuronales: en la sustancia gris son células satélite para las neuronas (<strong>com</strong>o los<br />
astrocitos protoplasmáticos); no se conoce bien su función<br />
Hay enfermedades desmilinizantes del encéfalo y su tratamiento es escaso. Se produce progresivamente<br />
con una pérdida paralela de las funciones mentales. Hoy en día, la terapia está orientada en inocular<br />
oligodentrocitos no afectados y sin diferenciar para poder remilinizar las fibras nerviosas; es preciso conocer<br />
primero dónde se localizan los oligodentrocitos precursores.<br />
Los precursores de los oligodentrocitos poseen el factor de transcripción Olig2, que regula genes que<br />
intervienen en la función normal del oligodentrocito. También se ha descubierto una proteoglucano de la<br />
superficie del oligodentrocito maduro que se ha denominado <strong>com</strong>o NG2, que puede usarse mediante Ac para<br />
detectar los oligodentrocitos en biopsias (son células NG2+).<br />
Las células de la macroglía son los astrocitos y los oligodentrocitos.
Página8<br />
Microglía<br />
Representa el 20% de todas las células gliales en el encéfalo y el 10-15% de todas las células del SN. Es un<br />
apoblación celular muy abundante. Son células de pequeño tamaño que se organizan cerca de los capilares del<br />
SNC adoptando una morfología estrellada y que están asociadas a la microcirculación encefálica. Se las<br />
descubrió en 1880 porque llamaba la atención sus fagosomas, por lo que parecían células facogíticas del SN; fue<br />
Del Río Ortega 30 años después quien las englobó en el SN, aunque hasta 1980 no se pudieron descubrir sus<br />
precursores hematopoyético. De todos modos, aunque forme parte de la estirpe hematopoyética y en la línea<br />
celular fagocítica, también tienen origen en el neuroectodermo. Por ello, tienen un doble origen: cresta neural y<br />
médula ósea.<br />
En el SN es la primera célula que se activa si hay un trastorno <strong>com</strong>o una trombosis cerebral, si hay un lesión<br />
traumática, encefalitis por infección viral o si se padece una enfermedad neurodegenerativa <strong>com</strong>o Parkinson o<br />
Alzheimer. Para esta detección poseen receptores para antígenos extraños, moléculas de desecho, proteínas<br />
inflamatorias y otros muy sensibles al estrés oxidativo e hipoxia. Cuando se activan estas células, van a<br />
convertirse en la población que defienda el encéfalo: proliferan, aumentan de tamaño y activan otras células. Si<br />
aparece una alteración <strong>com</strong>o un tumor o una infección, la microglía se activa rápidamente y se acumula en esa<br />
zona.<br />
Las células de la microglía tienen un citoplasma muy denso, fagosomas, muchos lisosomas, cuerpos<br />
residuales y son APC. De todos modos, también pueden secretar algunas neurotrofinas que activan a los<br />
astrocitos y a las neuronas, la proliferación de los oligodentrocitos y van a regular el paso de leucocitos al<br />
encéfalo. Los leucocitos no aparecen en encéfalo porque de su actividad resulta una gran cantidad de agentes<br />
oxidativos que pueden dañar a las neuronas, por lo que en condiciones normales las células de la microglía<br />
pueden liberar factores antimigratorios de los leucocitos, aunque en situación de daño grave se inhibe la<br />
secreción de estos factores.<br />
Las células de la microglía permiten eliminar las neuronas muertas con colaboración de los macrófagos.<br />
Células Ependimarias<br />
Las células ependimarias son células gliales con origen en la cresta neural que se organiza a modo de<br />
epitelio para revestir la superficie interna de los ventrículos cerebrales y el canal central de la ME (canal<br />
ependimario). Se sitúan en la superficie interna de cavidades que contienen Liquido Encefalo Raquídeo (LER)<br />
separándose del parénquima cerebral, por lo que son barrea entre el LER y el parénquima cerebral regulando el<br />
tránsito de moléculas entre ambos.<br />
Pertenecen a la población de los ependimocitos: células cúbicas que forman una estructura epitelial sin<br />
apoyarse en membrana basal, aunque sí aparecen uniones entre ellas. Acumulan glucógeno, tienen un gran<br />
aparato de Golgi, tienen lisosomas, microfilamentos y en su superficie apical presenta cilios, que se encuentra<br />
embebidas en el LER.<br />
Por debajo de ellas existen prolongaciones subependimarias de astrocitos que forman un lecho para la<br />
estructura.<br />
Hay otra población de células ependimarias que se conocen <strong>com</strong>o tanicitos: células cúbicas que poseen un<br />
proceso citoplasmático que se dirige hacia el interior del parénquima cerebral y que suelen rodear a los<br />
capilares. Los tanicitos permiten mediante su prolongación el paso de moléculas desde el capilar al LER y<br />
viceversa. Estas células carecen de cilios, aunque en ocasiones aparecen microvellosidades.<br />
Células Gliales del SNP<br />
Células de Schwann<br />
Células satélite o capsulares de los ganglios periféricos
Página9<br />
FIBRA NERVIOSA<br />
Una de las principales diferencias del SN con respecto al resto de los tejidos es la formación de circuitos<br />
que permite la integración del cuerpo. Los circuitos, observados a nivel microscópico, están formados por fibras<br />
nerviosas muy parecidas en el SNC y SNP.<br />
Las fibras nerviosas son axones neuronales rodeados por células gliales especializadas, que en el caso del<br />
SNC son los oligodentrocitos y en el SNP son las células de Schwann.<br />
Los oligodentrocitos son capaces de enviar 20- 50 prolongaciones citoplasmáticas que recubren pequeñas<br />
zonas del axón. En cambio, en el SNP los axones son más gruesos y largos, por lo que la célula de Schwann sólo<br />
recubre pequeñas micras de longitud de una fibra axónica. Para poder recubrir los axones, las células de<br />
Schwann dan hasta 50 vueltas muy <strong>com</strong>pactadas alrededor del axón; el gran empaquetamiento produce una<br />
eliminación del citoplasma en la zona enrollada concentrándose todo en el núcleo. La zona que se enrolla en el<br />
axón es fundamentalmente membrana, muy rico en esfingomielina, por lo que a estos axones se les conoce<br />
<strong>com</strong>o mielínicos. La vaina de mielina estabiliza la corriente eléctrica que se distribuye por el axón.<br />
Entre dos células de Schwann adyacentes siempre queda un espacio no recubierto por mielina que se<br />
conoce <strong>com</strong>o nodo de Ranvier, mientras que los recubiertos por mielina son los segmentos internodales. Las<br />
hojas se asientan en forma de lengüeta sobre la zona paranodal; los nodos de Ranvier son abultados y tiene una<br />
gran cantidad de canales de Ca voltaje-dependientes, por lo que la transmisión nerviosa es saltatoria de nodo a<br />
nodo.<br />
Por encima del nodo de Ranvier se forman uniones estrechas heterotípicas entre las hojas mielínicas de<br />
dos células de Schwann diferentes. También se forman uniones estrechas homotípicas entre las diferentes hojas<br />
de la vaina de mielina de la misma célula. Es decir, la célula de Schwann tiene dos tipos de uniones estrechas:<br />
heterotípicas y homotípicas que son atacados por autoanticuerpos, generando patologías autoinmunes<br />
desmielinizantes.<br />
Génesis de la Vaina de Mielina<br />
La vaina de mielina se forma por enrollamiento de la prolongación citoplasmática de la célula de Schwann,<br />
con la consecuente pérdida de citoplasma hacia el núcleo; en ocasiones quedan restos de citoplasma que<br />
forman las incisuras de Schmidt-Lantenan y aparecen de forma clara.<br />
La mielina al MET (Microscopio Electrónico de Transmisión) aparece una línea densa mayor formada por<br />
<strong>com</strong>pactación del ínfimo citoplasma remanente con las capas internas o citoplasmáticas de la bicapa lipídica y<br />
una línea densa menor formada por dos capas externas de la bicapa de dos células diferentes con el resto de<br />
espacio extracelular. Por ello, si hay 50 vueltas habrá un total de 100 líneas densas en total.<br />
Existen <strong>com</strong>unicaciones homotípicas a diferentes niveles de la vaina de mielina entre los puntos donde<br />
aparecen incisuras de Schmidt-Lantenan mediante canales de conexina 32 para el paso de iones desde el<br />
exterior celular y de la vaina al axón; si se forman autoanticuerpos contra esta conexina, se forman<br />
enfermedades desmielinizantes, <strong>com</strong>o la esclerosis múltiple.<br />
Las células de Schwann no sólo protegen los axones, si no que además regeneran las fibras nerviosas por su<br />
capacidad de regeneración. Reparan el axón y lo revitalizan funcionalmente. Sin las células de Scwhann no es<br />
posible regenerar las fibras nerviosas. Desde el punto de vista clínico tienen mucha importancia, porque<br />
individuos jóvenes que pierden inervación somática o visceral pueden recuperar fibras nerviosas. Actualmente<br />
se conoce que las células de Schwann sufren procesos retrodiferenciativos con liberación de factores de<br />
crecimiento y neurotrofinas; una de las neurotrofinas más conocidas es el factor de crecimiento neural. Gracias<br />
a este polipéptido, los axones pueden orientarse cuando se regeneran en la dirección del órgano que tienen que<br />
inervar. Por ello, el factor de crecimiento neural es una molécula de gran importancia en la regeneración axonal<br />
y en dirigirlo en una dirección determinada.
Página10<br />
Lesión Proximal<br />
Si hay un traumatismo en la salida del axón a partir del<br />
cuerpo neuronal conduce a una degeneración proximal de la<br />
fibra nerviosa. En este caso el cuerpo neuronal sufre cambios<br />
funcionales llamados la degeneración axonal??. A partir de aquí<br />
se produce la regeneración del axón. Pero, debido a la longitud<br />
que debe recorrer el nuevo axón hay proliferación de células de<br />
Schwann que guían y tutelan el recorrido que debe realizar el<br />
nuevo axón. El proceso dura algunas semanas hasta que se<br />
reinerva el órgano denervado.<br />
Lesión Distal<br />
Si hay una lesión distal (región axonal alejada del soma<br />
neuronal) se produce la degeneración del extremo distal o<br />
Walleriana. En estos casos lo que queda de axón emite una serie<br />
de prolongaciones que pretenden de nuevo encontrar al órgano<br />
objeto de inervación. Llama la atención la proliferación de las<br />
células de Schwann que van creando correderas que son el<br />
nuevo axón en desarrollo. Forman una especie de banda<br />
llamada banda de Bunger que de alguna manera guían el<br />
desarrollo axonal.<br />
Para que finalice el proceso es necesaria la activación de<br />
macrófagos en la zona para eliminar restos axonales y de<br />
mielínica (se puede evidenciar con gránulos de mielina en<br />
macrófagos de la zona). Por ello, se cree que son el sistema<br />
monocito-macrófago y el proceso inflamatorio los responsables<br />
de la activación de las células de Schwann. Esto ocurre en los<br />
dos tipos de lesión.<br />
Endoneuro<br />
Vaina de tejido conjuntivo reticular que rodea individualmente cada fibra nerviosa. Tiene una gran<br />
importancia en las fibras nerviosas porque aporta la matriz extracelular que rodea a las células de Schwann y<br />
porque sirve para que cada fibra nerviosa se pueda pegar a sus vecinas en la formación de fascículos nerviosos.<br />
Así, los fascículos nerviosos no se deshilachan en los nervios que le <strong>com</strong>ponen. Se denomina cemento porque<br />
absorbe los factores de crecimiento que necesite la célula nerviosa en caso necesario. Así, si una fibra nerviosa<br />
se lesiona, también se rompe el cemento adyacente, de modo que se liberan factores de crecimiento que activan<br />
a las células de Schwann para la regeneración axonal.<br />
El endoneuro también tiene capacidad proangiogénica, es decir, es capaz de atraer capilares a las fibras<br />
nerviosas lesionadas gracias a factores de crecimiento liberados en la ruptura del endoneuro de la zona<br />
lesionada. La irrigación facilita la regeneración axonal aportando nutrientes y recogiendo los desechos de las<br />
células de Schwann, que tienen una gran actividad metabólica. La correcta irrigación permite una correcta<br />
mielinización del axón; cuando se perturba la irrigación de las fibras nerviosas se dan problemas de conducción<br />
debido a los defectos de mielinización. Además, la irrigación permite la llegada de neurotrofinas (incluso<br />
encefálicas) y de leucocitos a la zona interactuando previamente con el endotelio.<br />
Perineuro y Epineuro<br />
Es un tejido conjuntivo más estructural que aglutina los fascículos nerviosos formando nervios. Es un tejido<br />
conectivo denso y fibroso, pero de alta vitalidad porque es el lecho por donde discurren los vasos sanguíneos<br />
que nutren a los fascículos y sus fibras nerviosas. El perineuro es el que más se inflama en las neuropatías<br />
porque debido a su alta vascularización existe una gran llegada de leucocitos.
Página11<br />
Rodeando al perineuro se encuentra el epineuro, la cobertura más externa de tejido conjuntivo de un<br />
nervio. Es un tejido conjuntivo denso, fibroso y apenas vascularizado que principalmente permite proteger el<br />
nervio e independizarlo de su entorno.<br />
Inmunidad de los Nervios<br />
La inflamación en los nervios suelen estar inhibidas porque la inflamación cursa con disfunción, lo que no<br />
es <strong>com</strong>patible. Por ello, los tejidos liberan factores antimigratorios que intentan evitar la aparición de<br />
neuropatías inflamatorias con demasiada frecuencia. Por esta razón, la presencia de vasos linfáticos es mínima<br />
y despreciable en los nervios.
Página12
Página13<br />
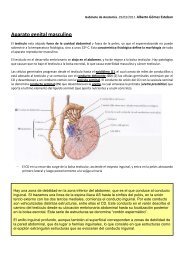
MÉDULA ESPINAL<br />
Configuración Externa de la ME<br />
La ME es una estructura alargada por la que el cerebro se <strong>com</strong>unica con el resto del cuerpo. Se extiende<br />
desde el foramen magno hasta L1-2. Tiene una longitud de 43-45cm, sin llegar a ocupar la totalidad del<br />
conducto vertebral; en este espacio donde no hay ME es donde realizan las punciones porque así no se lesiona<br />
la ME.<br />
La ME es segmentaria porque filogenéticamente es la estructura más antigua del SNC para permitir la<br />
integración del cuerpo. Desde el punto de vista morfo-funcional es mucho más sencilla. No sólo es el lugar de<br />
paso de los axones del encéfalo, ya que también es capaz de organizar respuestas por sí mismas, <strong>com</strong>o por<br />
ejemplo los reflejos simples; se usan en la clínica para evaluar diferentes segmentos medulares¸ <strong>com</strong>o el reflejo<br />
patelar para L1-L2 o el bicipital para C5-C6.<br />
El límite superior de la SE se une con el encéfalo a nivel del bulbo y el límite inferior termina en el cono<br />
medular cuyo vértice se ubica a la altura de L1-L2. De aquí parte el ligamento coccígeo o filum terminale<br />
(duramadre) que lo fija inferiormente, aunque esta fijación está también aumentada por la salida de los nervios<br />
espinales y por la unión superior con el encéfalo.<br />
La ME está segmentada en 31 segmentos, que representan el propio origen embriológico de la ME. De cada<br />
segmento parten los nervios espinales por los agujeros intervertebrales, por lo que salen 31 pares de nervios<br />
espinales.<br />
La ME es desde fuera de color blanco porque aparecen axones muy mielinizados. Presenta dos<br />
intumescencias o engrosamientos: cervical y lumbar.<br />
Configuración Interna de la ME<br />
Sustancia Blanca<br />
Los cordones están formados por sustancia blanca. Existe un cordón A (entre fisura medial anterior y surco<br />
ventrolateral), un cordón lateral (entre surcos ventrolateral y dorsolateral) y un cordón P (entre surco<br />
dorsolateral y fisura medial dorsal).<br />
Sustancia Gris<br />
La sustancia gris cambia desde la zona más superior de la ME a la más inferior. En el segmento C la<br />
sustancia gris es muy abundante, disminuye en T y vuelve a aumentar en L; los aumentos de sustancia gris se<br />
corresponden a la inervación de miembros. En el asta anterior hay dos segmentos (cabeza y base), mientras<br />
que el asta posterior tiene 3 segmentos (cabeza, cuello y base). La <strong>com</strong>isura gris une las dos astas; la sustancia<br />
gris alrededor del conducto ependimario es la sustancia gris intermedia (es parte de la <strong>com</strong>isura gris).<br />
ASTA ANTERIOR<br />
El asta anterior contiene las motoneuronas:<br />
α-motoneuronas. Son neuronas piramidales muy grandes<br />
Golgi tipo I: células piramidales o estrelladas con axón largo. Hay de 3 tipos:<br />
o Neuronas Radiculares: sus axones forman parte de la raíz A<br />
o Neuronas Cordonales: sus axones se introducen en los cordones cercanos de sustancia blanca<br />
para poder asociar diferentes segmentos medulares.<br />
o Gamma-motoneuronas: son pequeñas y radiculares<br />
Motoneuronas de axón corto: Renshaw son inhibidoras de las anteriores
Página14<br />
ASTA LATERAL<br />
Neuronas de asociación: son las neuronas de asociación, intercalares o internunciales. Son Golgi tipo II y<br />
asocian las astas del asta posterior con las del asta A del mismo lado. También se las llama neuronas de ajuste<br />
porque tienen memoria para modular informaciones del asta A y P.<br />
Neuronas <strong>com</strong>isurales: conectan las neuronas de astas de lados opuestos<br />
ASTA POSTERIOR<br />
Hay neuronas de axón corto que son receptoras. Reciben impulsos desde las raíces P que han sinaptado en<br />
el ganglio raquídeo que contiene neuronas seudomonopolares o en T. Es la zona sensorial y se da un filtro de<br />
estímulos.<br />
Se distinguen en el asta posterior 3 zonas:<br />
Cabeza es la parte más distal y recibe estímulos propioceptivos<br />
Cuello que recibe estímulos del aparato osteoarticular<br />
Base, que contiene neuronas que reciben estímulos interoceptivos de órganos endodérmicos.
Página15<br />
GLOBO OCULAR<br />
Embriología Básica<br />
Embriológicamente, el globo ocular deriva de unas proyecciones del encéfalo anterior que forman las<br />
vesículas ópticas recubiertas de ectodermo, que formará las cubiertas externas. Además, el globo ocular recibe<br />
mesénquima que forma la capa media muscular.<br />
Se pueden distinguir 5 capas en el ojo del embrión:<br />
1. Epitelio<br />
2. Membrana de Bowman<br />
3. Estroma<br />
4. Membrana Descendente<br />
5. Endotelio<br />
Las capas 2 y 4 son capas acelulares que sirven <strong>com</strong>o membranas basales de las capas 1 y 5.<br />
Espacios o Zonas del Globo Ocular<br />
Se distinguen varios espacios:<br />
<br />
<br />
<br />
Parte externa entre esclerótica y córnea, es de naturaleza epitelial y fibrosa cuya función es proteger.<br />
Embriológicamente tiene origen endodérmico.<br />
Capa media del globo ocular tiene naturaleza mesenquimal y está <strong>com</strong>prendido en anterior por el<br />
iris, los cuerpos ciliares en lateral y el coroides en posterior.<br />
Los cuerpos ciliares excretan humor acuoso, el iris regula la entrada de luz y la coroides es una capa<br />
<strong>com</strong>prendida entre la esclerótica y la retina cuya función es vascular.<br />
Parte interna es fotosensible porque sus capas celulares incluyen a fotorreceptores y células gliales;<br />
es, a fin de cuentas, la parte nerviosa del órgano.
Página16<br />
Córnea<br />
Es la estructura más anterior, transparente y convexa y forma parte del sistema dióptrico (n = 1,36). Tiene<br />
un grosor de casi 1 mm y su diámetro de es 1cm. Su grosor es mayor en la zona periférica que en el centro<br />
(1,1mm en el limbo esclerocorneal a 0,8-0,9mm del centro).<br />
Epitelio Corneal Anterior<br />
El epitelio corneal anterior es un epitelio plano estratificado no queratinizado formado por 5-8 capas de<br />
células unidas entre sí por desmosomas y mediante hemidesmosomas con la membrana basal (membrana de<br />
Bowman); este epitelio tiene una altura de 50µm .<br />
El epitelio está expuesto al medio, por lo que recibe el impacto de partículas del aire, aunque su gran<br />
capacidad proliferativa permite que esté en condiciones óptimas a pesar de la gran descamación y daños que se<br />
producen en la córnea.<br />
El epitelio corneal anterior se apoya sobre la membrana de Bowman, una banda PAS+ de matriz<br />
extracelular con un diámetro de 5-7µm, descubierta por Bowman. Es un tejido conjuntivo laxo que contiene<br />
fibras de colágeno IV y V dispuestas paralelamente para permitir el paso de luz. Es un tejido avascular y no<br />
entran leucocitos (es un tejido inmunoprivilegiado) gracias a factores antimigratorios y antiangiogénicos que<br />
secretan las células del estroma.<br />
Obviamente, existen condrocitos en el tejido (fibroblastos especializados).<br />
La trasparencia de la córnea se encuentra asegurada por la disposición del colágeno y la presencia en la<br />
sustancia fundamental de condroitín-sulfato y queratán sulfato.<br />
Tanto el epitelio <strong>com</strong>o la membrana de Bowman están inervados por terminaciones nerviosas sensoriales.<br />
Estroma de la Córnea<br />
Membrana de Descemet<br />
Es una lámina basal que tiene una parte fibrosa anterior y una lámina basal típica posterior. La lámina<br />
basal verdadera sirve de apoyo para una membrana córnea posterior y evita la formación de edemas por humor<br />
acuoso, ya que es impermeable.<br />
El epitelio es monoestratificado y aparecen bombas iónicas que regulan el gradiente.<br />
Endotelio<br />
Este epitelio evita la formación de edemas.<br />
Esclerótica<br />
Es una capa blanquecina que recubre el globo ocular; el color blanco proviene de su estructura fibrosa.<br />
Presenta:<br />
<br />
<br />
<br />
<br />
<br />
Capa fibrosa externa: haz colágeno entrecruzado<br />
Capa tensional: con fibras de colágeno que evitan el agrandamiento y sus consecuencias, <strong>com</strong>o el<br />
aumento de la presión intraocular. Protege el globo ocular. Tiene pocos vasos sanguíneos.<br />
Capa fina de melanocitos: dan opacidad<br />
Capa cribosa: pasan vasos y nervios a través de ella hacia el nervio óptico; se localiza en el polo P del<br />
globo.<br />
Capa supracoroidea: es la más profunda
Página17<br />
ÚVEA<br />
La úvea es la parte media del globo ocular y se puede dividir en varias partes:<br />
Coroides<br />
Se extiende desde la porción posterior del globo hasta la ora serrata. Es una capa vascular y melanogénica que<br />
se <strong>com</strong>pone de varias capas:<br />
<br />
<br />
<br />
<br />
<br />
Membrana de Bruch: está en contacto con el epitelio pigmentario de la retina. En realidad, es <strong>com</strong>o una<br />
membrana basal que acumula Ca y lípidos<br />
Capa Coriocapilar: tiene capilares fenestrados permeables para nutrir a la retina<br />
Capa Vascular: tiene arterias y venas que dan lugar a capilares; se asocia a tejido conjuntivo fibroso<br />
Capa Supracoroidea: asociada a melanocitos que forman el tejido conjuntivo pigmentario<br />
Espacio pericoroideo: mirando a la esclerótida<br />
Cuerpos Ciliares<br />
Son anillos fibromusculares que sostienen el cristalino y regulan su ensanchamiento o alargamiento. Este<br />
anillo está en continuidad con la coroides.<br />
Hay una zona A plegada y contiene de 70 a 80 invaginaciones que forman los procesos ciliares cuya función<br />
es sujetar el cristalino. El proceso ciliar está tapizado por el epitelio del proceso ciliar que secreta el humor<br />
acuoso, que rellena las cámaras A y P del globo ocular.<br />
Hay también una parte posterior no plegada.<br />
Epitelio Ciliar<br />
Es un epitelio biestratificado que recubre los procesos ciliares.<br />
EPITELIO NO PIGMENTARIO O EXTERNO<br />
La célula que sostiene al cristalino es<br />
una célula externa y cúbica que tiene<br />
microvellosidades de las que parten fibras<br />
zonulares, que se enganchan en el<br />
cristalino. Además, estas células secretan<br />
el humor acuoso. En su citoplasma se<br />
encuentran muchas mitocondrias y<br />
fagosomas, por lo que se determina que<br />
son fagocíticas.<br />
Las células están unidas por uniones<br />
estrechas que lo hacen impermeable.<br />
Las fibras zonulares son<br />
microfilamentos de 10nm de diámetro<br />
que, en conjunto, forman el aparato<br />
suspensorio del cristalino. La primera<br />
asociación de las fibras zonulares es de<br />
100-140 fibras formando los fardos, que<br />
pueden tener 2 orientaciones: las fibras de<br />
la porción plegada van hacia P y las de la<br />
porción lisa van a A, por lo que se cruzan y<br />
forman un espacio triangular llamado<br />
triángulo o espacio de Petit.
Página18<br />
EPITELIO PIGMENTARIO O INTERNO<br />
Las células de la capa interna del epitelio ciliar tienen melanina. La melanina aparece por la gran cantidad<br />
de melanocitos en la capa estromal del cuerpo ciliar; pueden darse melanomas<br />
Ambas capas están unidas funcionalmente porque tienen que trasladar el humor acuoso que extraen de los<br />
capilares fenestrados y altamente permeables del estroma del cuerpo ciliar hasta el interior de las cámaras A y<br />
P del globo ocular.<br />
El humor acuoso permite que la córnea pueda ser nutrida desde su cara P en un transporte mediado por la<br />
capa de Descemet.<br />
Músculos del Cuerpo Ciliar<br />
Se encuentran en el estroma del cuerpo ciliar y pueden ser de 2 tipos:<br />
<br />
<br />
Músculo de Müller: se dispone de forma circular y, cuando se contrae, se relaja la sujeción del cristalino,<br />
por lo que pierde la tensión y se ensancha (disminuye el diámetro). Está regulado por el SNPS.<br />
Músculo de Brucke: son perpendiculares a las fibras anteriores, por lo que forman una estructura radial<br />
que al contraerse adelgazan el cristalino (aumentan el diámetro).<br />
Iris<br />
Es una placa anular delgada de 12mm de diámetro que se localiza la parte A de la úvea y que tiene un<br />
orificio llamado pupila en su interior.<br />
El epitelio A de la placa es rugoso y tiene fibras de colágeno plegadas mezcladas con fibroblastos y melanocitos.<br />
En la parte P está el estroma del iris, un tejido conectivo melánico donde se concentran una cantidad de<br />
melanocitos: mayor cantidad de melanocitos supone un color más oscuro y una menor cantidad los hace más<br />
claros. En el estroma hay también estructuras vasculares que forman rugosidades, observables<br />
macroscópicamente. La gran cantidad de melanocitos puede producir melanomas.<br />
El epitelio posterior tiene células pigmentarias que se apoyan sobre el cristalino.<br />
El iris tiene además un músculo que rodea la pupila cuya contracción empequeñece el diámetro de la pupila (su<br />
relajación lo amplía) y un músculo dilatador de la pupila, que se dispone radialmente a la pupila y cumple<br />
acciones contrarias al músculo anterior.
Página19<br />
Cristalino<br />
Es un órgano transparente biconvexo cuya misión es enfocar la imagen sobre la retina. El cristalino está<br />
sujeto por las fibras zonulares y está en contacto con el epitelio posterior del iris.<br />
Histológicamente se distingue una capa envolvente impermeable, incluso a las células migratorias <strong>com</strong>o<br />
leucocitos, que podría degradar al cristalino. Además, es un tejido sin vascularización ni inervación.<br />
Por debajo de la cápsula envolvente se distingue en su cara A el epitelio A del cristalino. Es un epitelio<br />
cúbico en cuya capa más central se dan mitosis que permiten renovar las estructuras del propio cristalino,<br />
aunque las divisiones se dan lentamente. Las células tienen un tamaño máximo cúbico en esta zona central de la<br />
capa A (tras la mitosis), pero según van migrando hacia lateral adoptan una estructura más fibrilar, por lo que<br />
adoptan el nombre de fibras lenticulares que se disponen paralelamente.<br />
La transparencia y el n se mantiene por la disposición geométrica de sus células y por su <strong>com</strong>pactación.<br />
Entre las células aparecen GAP-junctions que permiten la <strong>com</strong>unicación entre células; estas células son las<br />
más ricas en conexina del organismo. Así, las células más externas reciben nutrientes y factores solubles que<br />
difunden a su citoplasma y de ahí al resto del cristalino.<br />
Hay 3 zonas en el cristalino:<br />
Zona central o nuclear: células muertas sin núcleo que poseen un<br />
citoplasma pobre<br />
Zona de la corteza: es amplia y perinuclear y en ella las células<br />
subsisten en quiescencia por difusión de nutrientes<br />
Zona periférica: formada por células cúbicas.<br />
Retina<br />
Es la parte fotosensible del ojo. La parte A no es funcional y se relaciona con la ora serrata, cuerpos ciliares<br />
y parte P del iris.<br />
La parte P se extiende desde la ora serrata hasta la parte P del globo ocular y es la parte fotosensible.<br />
En la retina hay 3 tipos celulares:<br />
Fotorreceptores: conos y bastones<br />
Glía: células de Müller<br />
Neuronas intrínsecas de la retina: amacrinas, ganglionares, bipolares y horizontales<br />
Las capas de la retina son, de exterior a interior:<br />
1. Epitelio pigmentario de la retina: está formada por células de aspecto cúbico que se relacionan con la<br />
coroides en A.<br />
2. Capa de conos y bastones: es la capa fotosensible<br />
3. Capa limitante externa: tiene <strong>com</strong>plejos de unión entre los fotorreceptores y las células de Müller, unas<br />
células gliales del tipo astrocitos especializados en el globo ocular.<br />
4. Capa nuclear externa: es donde se alinean los núcleos de los fotorreceptores<br />
5. Capa plexiforme externa: es un plexo en el que hay sinapsis de los fotorreceptores con las neuronas de<br />
la retina, <strong>com</strong>o las neuronas bipolares y las neuronas horizontales.<br />
6. Capa nuclear interna: están los núcleos de las principales neuronas de la retina; también están los<br />
núcleos de las células de Müller. También están los núcleos de las neuronas amacrinas<br />
7. Capa plexiforme interna: sinapsis entre neuronas bipolares y neuronas ganglionares y amacrinas<br />
8. Capa de células ganglionares<br />
9. Capa de fibras nerviosas<br />
10. Membrana Limitante Interna: es la membrana basal de las células de Müller.
Página20
Página21<br />
OÍDO<br />
OÍDO EXTERNO<br />
Oreja<br />
Tiene una estructura cartilaginosa con piel diferente en ambos lados:<br />
La piel A es fija e inmovilizada por el pericondrio.<br />
La piel P es móvil y tiene más hipodermis. Además, presenta pelos (en la A son muy escasos).<br />
El lóbulo de la oreja tiene un eje de hipodermis sin cartílago y está muy vascularizada; en cardiología el lóbulo<br />
de la oreja hinchado suele indicar congestión cardiovascular.<br />
El cartílago de la oreja es elástico por un pericondrio rico en fibras elásticas.<br />
Conducto Auditivo Externo<br />
Canal de 25mm de longitud y 8mm de diámetro que pone en contacto el exterior con la membrana timpánica.<br />
La estructura de este conducto tiene 2 partes:<br />
<br />
<br />
Parte cartilaginosa externa recubierta por una epidermis sin apenas papilas pero muchos pelos, muchas<br />
glándulas sebáceas y glándulas ceruminosas. Las glándulas ceruminosas son de tipo túbulo-alveolar<br />
cuya secreción es muy densa y lipídica, también rica en sustancias pigmentarias y con moléculas muy<br />
bacteriocidas.<br />
Parte ósea interna recubierta por epidermis fina sin glándulas ceruminosas ni pelos que llega a la cara A<br />
de la membrana timpánica.<br />
OÍDO MEDIO<br />
Caja Timpánica<br />
Se encuentra en el seno del hueso temporal y está delimitada lateralmente por la membrana timpánica, y<br />
posteriormente por las células mastoideas.<br />
Está revestida por una mucosa con un epitelio plano simple, a veces cúbico e incluso algo<br />
seudoestratificado, según la zona donde se localice; en los niños, el epitelio es ciliar y pueden encontrarse<br />
células caliciformes.<br />
La lámina propia es de tejido conjuntivo laxo ricamente vascularizada e inervada, lo que conlleva gran<br />
dolor en inflamación del oído medio.<br />
La cavidad timpánica contiene los huesecillos del oído. Estos huesecillos están articulados por<br />
articulaciones diartrósicas y están tensionados por los tendones del músculo tensor del tímpano y del músculo<br />
estapedio; la inervación viene dada por el nervio cuerda del tímpano, rama del facial. En conjunto, el objetivo es<br />
la transmisión de las vibraciones.<br />
Los huesecillos son tres: estribo, yunque y martillo. Son huesos recubiertos por un epitelio cúbico simple<br />
sobre el periostio. Los huesecillos finalmente terminan en la ventana oval.<br />
Los tendones de las musculaturas tensoras del tímpano son musculatura esquelética que llegan al<br />
manubrio del yunque. Tiene un gran endomisio que lo hace muy irrigado.<br />
El músculo estapedio es del estribo. Es también musculatura esquelética y llega hasta la cabeza del estribo.
Página22<br />
Membrana Timpánica<br />
Es una membrana muy fina que separa el CAE de la cavidad timpánica. La membrana se <strong>com</strong>pone de dos partes:<br />
parte flácida y parte tensa.<br />
CARA LATERAL<br />
La membrana timpánica está recubierta por una parte cutánea consistente en una epidermis fina que<br />
protege la membrana timpánica y que se apoya en una dermis fina que se inserta en la estructura de la<br />
membrana timpánica.<br />
La estructura central de la membrana timpánica es fibrosa con fibras de colágenos dispuestas de dos<br />
maneras: unas de forma circular (porción más interna) y otras de forma radial desde el centro hacia los<br />
extremos de la membrana (similar a una tela de araña).<br />
Estas fibras se <strong>com</strong>binan con un cartílago hialino con forma de anillo y dispuesto en los límites externos y<br />
forma el anillo fibrocartilaginoso de la membrana timpánica.<br />
En la membrana timpánica también hay fibras elásticas, vasos y nervios, por lo que es sensible.<br />
CARA MEDIAL<br />
La parte mucosa mira hacia la caja timpánica. Está muy vascularizada por las ramas de la arteria del CAE y<br />
cavidad timpánica. Los nervios derivan del nervio aurículo-temporal, aunque también hay ramificaciones del<br />
nervio vago y de ramas timpánicas del nervio glosofaríngeo.<br />
Trompa de Eustaquio<br />
Tiene unos 4cm de longitud y ponen en <strong>com</strong>unicación la faringe con la cavidad timpánica. Se pueden apreciar<br />
dos partes:<br />
<br />
<br />
Parte ósea que representa el 1/3 lateral: epitelio simple cilíndrico ciliar; se apoya en un tejido<br />
conjuntivo<br />
Parte ósea, que representa los 2/3 mediales: tiene un epitelio seudoestratificado con muchas células<br />
caliciformes, glándulas tubulo-alveolares y la amígdala tubárica (conglomerado de nódulos linfáticos<br />
solitarios que forman parte del anillo de Waldeyer)<br />
OÍDO INTERNO<br />
Es un conjunto de sáculos y túbulos alojados en cavidades labradas en el hueso temporal y que contienen la<br />
perilinfa.<br />
Se distingue un vestíbulo y cóclea. Tiene una cápsula ósea de hueso <strong>com</strong>pacto cuyo interior se sitúan las<br />
estructuras auditivas. La cara interna está revestida por endostio bañado en perilinfa. La endolinfa es muy rica<br />
en K y muy escasa en Na, mientras que la perilinfa es rica en Na y pobre en K.<br />
Zona Vestibular<br />
El vestíbulo representa una cavidad ovoidea en donde terminan los canales semicirculares: 3 túneles dispuestos<br />
en los 3 ejes del espacio (superior, P y lateral).<br />
La zona vestibular contiene al utrículo y sáculo y está rodeada de endostio.<br />
Las crestas ampulares o ampollas de los conductos semicirculares, junto con las máculas de sáculo y utrículo,<br />
forman en conjunto las crestas del equilibrio. Están constituidas por un epitelio neurosensorial muy similar.
Página23<br />
Este epitelio es seudoestratificado formado por células ciliadas o de tipo I, que son redondas y tienen un<br />
cuello estrecho, desde donde se proyectan cilios sensoriales. Son de 40-80 cilios por célula formando un<br />
penacho entre los que sobresale un estereocilio. Esta célula está rodeada por una terminación sináptica en<br />
forma de cáliz del nervio vestibular para sostener y sinaptar así con las células.<br />
Las células sensoriales de tipo II tienen cilios mecanorreceptores y están inervadas basal y lateralmente por<br />
terminaciones nerviosas.<br />
Alrededor, hay células de sostén que están sujetas por las células nerviosas y su función es proteger y nutrir<br />
el tejido, ya que hasta estos epitelios no llegan los vasos.<br />
El fenómeno del equilibrio se da porque el líquido que va por los conductos semicirculares y utrículo y<br />
sáculo, se mueve y contacta con una membrana formada por otolitos (cuerpos cristalinos de 3 a 5 µm)<br />
embebidos en la otoconia, una matriz proteinácea que los embebe, que flota encima de los penachos de las<br />
células neurosensoriales tipo I y II. Al moverse la endolinfa, también se mueve la membrana y conlleva el<br />
movimiento de los cilios.<br />
Esta región recibe el nombre de ampollas y máculas porque forman <strong>com</strong>o manchas en el laberinto coclear.<br />
Zona Coclear o Auditiva<br />
La cóclea tiene 35mm de longitud y de 3-9mm de diámetro. Este tubo termina en el helicotrema y su eje es el<br />
modiolo o columna, en torno al que da 2 vueltas y media, de tejido óseo esponjoso, donde se sitúan el ganglio<br />
espiral, los nervios del VIII par y vasos. En el modiolo se forma la lámina espiral, una lámina de hueso desde<br />
donde arranca la membrana basilar y, superior a esta, la membrana de Reissner.<br />
La luz del canal está dividida por la lámina espiral en un espacio superior e inferior; el espacio superior, a su<br />
vez, está dividido por la membrana basilar en otros dos espacios. Así resultan 3 rampas o espacios:<br />
1. Rampa vestibular<br />
2. Rampa media o coclear<br />
3. Rampa inferior o timpánica<br />
La membrana basilar divide la luz del canal en las rampas vestibular y timpánica; la rampa de Reissner<br />
divide entre ellas la rampa media o coclear.<br />
En la zona más superior de la espiral sólo se ve una estructura central que es donde se unen la rampa<br />
vestibular y la timpánica para permitir que las vibraciones del sonido pasen de una rampa y otra. Estas dos<br />
rampas contienen perilinfa y la rampa media contiene líquido endolinfático.<br />
La lámina espiral tiene las células bipolares del ganglio espiral. Al ser bipolares el polo apical recoge<br />
información y el polo basal emite axones para formar el VIII par.<br />
Estría Vascular<br />
El líquido endolinfático debe tener los niveles<br />
de K en unos niveles fijos para conseguir la<br />
excitabilidad de las células sensoriales. Esto se<br />
consigue gracias la estría vascular, formada por<br />
un epitelio pseudoestratificado vascularizado (el<br />
epitelio es en términos generales avascular).<br />
Entre estas células están las marginales, que<br />
extraen K del plasma de los capilares que lo<br />
rodean. Tienen un laberinto basal similar a las<br />
células de los túbulos renales que les permite<br />
secretar K. También hay céluals basales e<br />
intermedias (en las zonas basales e intermedias<br />
del epitelio, respetivamente).
Página24<br />
También sirven para drenar aquellas moléculas perjudiciales para el órgano.<br />
Membrana de Reissner<br />
En la Membrana de Reissner hay células aplanadas con microvellosidades que se apoyan sobre una lámina<br />
basal y, bajo esta, hay colágeno que amortigua la fuerza que ejerce la rampa vestibular sobre la media o coclear.<br />
Limbo Espiral<br />
Del modiolo sale la lámina espiral que proyecta el limbo espiral en su ángulo interno. La membrana tectoria<br />
es una sustancia gelatinosa sintetizada por las células interdentales, que secretan mucopolisacáridos y<br />
proteínas, <strong>com</strong>o las glucoproteínas α y β tectorinas; las mutaciones de estas proteínas producen sordera.<br />
Rodete Espiral<br />
El rodete espiral tiene un epitelio cúbico que se continúan con las células de Claudius, unas células de soporte.<br />
Células del Órgano de Corti<br />
Células mecanotransductoras: tienen cilios que se excitan por contacto cuando la membrana tectoria se mueve.<br />
Esta excitación produce un potencial de acción que se transduce a una señal neurológica por los axones.<br />
Células de los pilares externos e internos forman, respectivamente, los pilares externo e interno que delimitan<br />
el túnel de Corti. Son células muy altas (70µm de altura) y se asocian en hileras a lo largo de todo el órgano.<br />
Pilares externos: 85µm de altura y hay 3800 células de pilares externos.<br />
Células de sostén o falángicas: tienen forma de pedestal y acogen a las células sensoriales. Hay 4-5 células<br />
falángicas en cada hilera. Estas células tienen prolongaciones apicales que sostienen la membrana tectoria para<br />
impedir que las prolongaciones de las células sensoriales estén en contacto continuo con la membrana tectoria.<br />
Hay células falángicas externas en el lado externo del túnel de Corti y células falángicas internas en el lado<br />
interno.<br />
Las células de Hensen y las células de Böttcher, células de sostén externas del órgano de Corti.<br />
Membrana Basilar<br />
Se extiende desde el rodete a la espiral. Hay 2 zonas:<br />
Externa: es la mayor (2/3) y contacta con el rodete mediante el ligamento espiral<br />
Interna: 1/3 y contacta con la lámina espiral<br />
La parte que mira hacia el órgano de Corti tiene una capa de células mesoteliales que se asocian con células de<br />
colágeno que forman una especie de ligamento que forman las cuerdas auditivas.
Página25
Página26
Página27<br />
CEREBRO<br />
Brodman, en 1909, estudió la arquitectura del cerebro en diversas capas.<br />
Las prolongaciones axónicas pueden teñirse con la tinción de Weigert que tiñe la mielina y puede observarse<br />
la mieloarquitectura.<br />
Para observar la citoarquitectura se usan tinciones <strong>com</strong>o la tinción de metionina o con el azul de metileno y<br />
era usada por Nissl.<br />
La angioarquitectura permite describir el encéfalo según la distribución de los vasos sanguíneos y a su nivel<br />
de funcionamiento. Para ello se usan tinciones específicas de células vasculares <strong>com</strong>o los Ac contra el CD31 que<br />
corresponde a la PECAM-1, específica del endotelio. Así, no se observan demás células neuronales.<br />
También puede realizarse una tinción para moléculas del encéfalo que permite conocer la<br />
quimioarquitectura del encéfalo según los neurotransmisores y neurotrofinas que contiene.<br />
Patrón de Brodman de la Corteza Cerebral<br />
Capa I o Molecular<br />
En la corteza frontal observó una primera capa que apenas tiene células y sólo suele tener dendritas de<br />
células más profundas. Debido a los <strong>com</strong>ponentes que posee, se le atribuye función asociativa.<br />
Capa II o Granular Externa<br />
Está formada por células pequeñas de tipo piramidal o estrellado pequeñas con un diámetro de unas<br />
10µm. De ellas salen dendritas ascendentes a la capa molecular. A esta capa granular externa se le<br />
otorga función receptora a través de sus dendritas.<br />
Capa III o Piramidal Externa<br />
Tiene neuronas con forma prismática de tamaño pequeño o mediano que pueden llegar a alcanzar hasta<br />
las 40µm de diámetro. Tienen axones que salen de esta capa y forman un tracto que se observa <strong>com</strong>o la<br />
estría de Kaes-Bechterer. Este tracto recoge axones que van a núcleos subcorticales, aunque también<br />
hay axones que van a capas superiores.<br />
La función que se asocia a esta capa es asociativa.<br />
Capa IV o Granular Interna<br />
Tiene una alta concentración de células estrelladas de pequeño tamaño y cuyas ramificaciones<br />
terminales normalmente horizontales van principalmente al tálamo formando la estría externa de<br />
Baillarger, que reúne los axones de estas neuronas formando un tracto.<br />
Es una capa receptora.<br />
Capa V o Piramidal Interna<br />
Tiene neuronas piramidales de gran tamaño que se llaman neuronas giganto-piramidales de Beth, que<br />
suelen estar en las circunvoluciones precentrales (área 4) y sus axones forman la estría interna de<br />
Baillarger. Estas neuronas piramidales otorgan a esta capa una función efectora.<br />
Capa VI o Multiforme<br />
Tiene células de diferente morfología y muy irregulares: fusiformes, estrelladas, piramidales,… Su<br />
función es poco relevante.<br />
Capa Ínfima (VII)<br />
Sería una séptima capa que no se considera en el patrón de Brodman porque es inconstante. Sería una<br />
especie de lámina basal que separa sustancia gris de sustancia blanca.
Página28<br />
Tipos de Corteza<br />
Este patrón de distribución neuronal es muy variable de unas zonas a otras. De esta manera, esta es la<br />
corteza homotípica que aparece principalmente en lóbulos frontal, parietal y temporal.<br />
Por el contrario, hay una corteza agranular de tipo heterotípico que carece de las capas granulares externa e<br />
interna (II y IV) y abundan las células giganto-piramidales en las cortezas límbica y motora.<br />
La corteza hipergranular tiene una gran cantidad de células estrelladas, <strong>com</strong>o en la corteza auditiva,<br />
sensorial, visual, tálamo o hipocampo. Además, hay una pérdida de capas piramidales.<br />
Entre estos dos extremos (tipos de corteza I y V) hay otros tipos de corteza intermedios.<br />
Mieloarquitectura<br />
De superficie del encéfalo al interior hay varias capas:<br />
I. Lámina Tangencial: una especie de lámina tangencial al cerebro de estrías superficiales<br />
II. Capa de Kaes-Bechterer<br />
III. Capa Baillanger externa<br />
IV. Capa Baillanger interna<br />
V. Capa subestriada<br />
VI. Capa de las columnas radiales: envía fibras nerviosas a la sustancia blanca; esta capa tiene los axones en<br />
dirección perpendicular a las anteriores (y a la superficie cerebral).<br />
Columnas Cerebrales<br />
Se observa que el encéfalo tiene asociaciones de neuronas en columnas verticales que forman una especie de<br />
cilindro a modo de área en el que las neuronas que la constituyen están unidas entre ellas. Estas áreas crean<br />
unidades funcionales llamadas columnas cerebrales en el encéfalo de 300µm de diámetro (3columnasx1mm).<br />
Se ha observado que además hay conexiones entre las columnas adyacentes.<br />
Angioarquitectura<br />
El encéfalo tiene restringida la llegada de células y moléculas debido a 3 barreras:<br />
<br />
<br />
<br />
Endotelio: no es fenestrado ni tiene poros, pero sí tiene muchos receptores para la transcitosis. Fuera de<br />
la membrana endotelial está la membrana basal gruesa del endotelio, a la que se unen los podocitos de<br />
los astrocitos.<br />
Membrana glial limitante perivascular: formada por los podocitos de los astrocitos en torno a los<br />
capilares. Los podocitos se unen entre ellos por uniones estrechas que impermeabilizan los espacios<br />
entre los pies perivasculares.<br />
Membrana glial limitante superficial: formada por los podocitos de los astrocitos bajo las meninges.<br />
La neurona está envuelta por los podocitos perineuronales del astrocito que regula el acceso de iones y<br />
metabolitos a su interior.<br />
En un glioma o un metástasis maligno, la quimioterapia no suele llegar al tumor porque no pasa la barrera H-E.<br />
Además, la gliosis es una reacción cicatrizante en torno a las neuronas para protegerlas (aunque sean malignas).<br />
Esto se convierte en un gran limitante del tratamiento.<br />
Los capilares penetran en las columnas y organizan estas según el patrón vascular, que es desde el interior<br />
del encéfalo a la superficie, donde puede contactar con la circulación subpial que llega al capa I del encéfalo.
Página29<br />
CEREBELO<br />
Capa Molecular<br />
Está <strong>com</strong>o vacía porque sólo tiene células estrelladas y células en cesto. También tiene las prolongaciones<br />
dendríticas de las células giganto-piramidales del tipo célula de Purkinje. Es por tanto asociativa y receptora.<br />
Capa de las Células de Purkinje<br />
En el límite entre capa molecular y capa granular están las células de Purkinje con un gran tamaño y un<br />
núcleo con nucléolo marcado forman un núcleo en ojo de búho. Tienen en su zona apical un árbol dendrítico<br />
que puede formar hasta 2000 sinapsis que pueden llegar a la zona molecular. Su axón es descendente y puede<br />
llegar hasta las células de la capa granular.<br />
Asociadas a las células de Purkinje hay 3 tipos de células gliales:<br />
a) Células de Fanjana<br />
b) Células de Begbmann<br />
c) Oligodentrocitos<br />
Capa Granular<br />
Tiene una gran cantidad de células de Golgi tipo II, que son pequeñas con un diámetro 6µm. Forman los<br />
glomérulos cerebelosos, unos espacios sin neuronas rodeados de astrocitos (glía) del tipo células en velo. En<br />
estos espacios convergen axones procedentes de las células granulares.<br />
También hay granos.