CAPITULO 1: REVISION MONOGRAFICA - Mecanismo de ...
CAPITULO 1: REVISION MONOGRAFICA - Mecanismo de ...
CAPITULO 1: REVISION MONOGRAFICA - Mecanismo de ...

You also want an ePaper? Increase the reach of your titles
YUMPU automatically turns print PDFs into web optimized ePapers that Google loves.
ANALISIS MORFOMETRICO Y DEMOGRAFICO DEEspeletia pycnophylla Cuatrecasas, EN UN GRADIENTEALTITUDINALPROVINCIA DE CARCHI-ECUADORADRIANA SÁNCHEZ ANDRADETrabajo <strong>de</strong> grado presentado para optar al título <strong>de</strong> BiólogoDirectorSANTIAGO MADRIÑANProfesor AsociadoLaboratorio <strong>de</strong> Botánica & SistemáticaUniversidad <strong>de</strong> los An<strong>de</strong>sCo-directorJASON T. RAUSHERProfesor VisitanteLaboratorio <strong>de</strong> Botánica & SistemáticaUniversidad <strong>de</strong> los An<strong>de</strong>sUNIVERSIDAD DE LOS ANDESFACULTAD DE CIENCIASDEPARTAMENTO DE CIENCIAS BIOLÓGICASBOGOTÁ D.C.ENERO, 2004
TABLA DE CONTENIDOAGRADECIMIENTOSivRESUMENvLISTA DE TABLASviLISTA DE FIGURASvii1. REVISIÓN MONOGRÁFICA 11.1. Ecosistema páramo 11.1.1 Descripción 11.1.2 Disturbio antrópico 41.2. El género Espeletia 71.3. Efecto <strong>de</strong>l gradiente altitudinal en la estructura poblacional 111.4. Demografía y crecimiento poblacional: Tablas <strong>de</strong> vida 142. OBJETIVOS 163. MATERIALES Y MÉTODOS 173.1. Espeletia pycnophylla Cuatrec 173.2. Area <strong>de</strong> estudio 173.3. Muestreo 213.4. Análisis <strong>de</strong> datos 223.4.1 Morfometría 223.4.2 Estructura poblacional 233.4.2.1 Densidad <strong>de</strong> individuos 233.4.2.2 Tablas <strong>de</strong> vida 233.4.3 Efecto <strong>de</strong> la comunidad vegetal en las poblaciones <strong>de</strong> Espeletiapycnophylla 243.4.3.1 Cobertura vegetal 243.4.3.2 Efecto <strong>de</strong> otras especies vegetales 243.4.4 Análisis estadístico 254. RESULTADOS 264.1 Morfometría 26ii
4.1.1 Altura <strong>de</strong> la roseta 264.1.2 Diámetro <strong>de</strong> la roseta 274.1.3 Capítulos por inflorescencia 284.2 Estructura poblacional 284.2.1 Densidad <strong>de</strong> individuos 284.2.2 Tablas <strong>de</strong> vida 294.3 Efecto <strong>de</strong> la comunidad 304.3.1 Porcentaje <strong>de</strong> Cobertura vegetal 304.3.2 Efecto <strong>de</strong> otras especies vegetales 314.3.2.1 Relación con altura y diámetro <strong>de</strong> la roseta 314.3.2.2 Relación con <strong>de</strong>nsidad <strong>de</strong> individuos 314.3.2.3 Relación con la altitud 32Tablas 33Figuras 375. DISCUSIÓN 516. CONCLUSIONES 667. RECOMENDACIONES 68LITERATURA CITADA 69ANEXOS 78ANEXO 1 78ANEXO 2 80ANEXO 3 82ANEXO 4 89ANEXO 5 93iii
AGRADECIMIENTOSQuisiera agra<strong>de</strong>cer, primero que todo a Santiago Madriñán, Director <strong>de</strong> este trabajo, porsu apoyo, sus opiniones y por la oportunidad <strong>de</strong> trabajar con él en el Laboratorio <strong>de</strong>Botánica y Sistemática. Al profesor Jason Rauscher, Codirector <strong>de</strong>l trabajo, por habermeaconsejado, por su paciencia y sus críticas.A la gente <strong>de</strong> Ecuador por su apoyo y colaboración. A Robert Hofste<strong>de</strong> y María VictoriaArbelaez por haberme acogido en su casa y por hacer que mi estadía en el país esté llena<strong>de</strong> buenos recuerdos. A la gente <strong>de</strong>l grupo Randi Randi, en especial al Proyecto ManrecurII, por facilitarme el trabajo <strong>de</strong> campo e información, y por su amistad. A la estaciónbiológica Guan<strong>de</strong>ra y a la reserva ecológica El Angel, por darme la oportunidad <strong>de</strong>trabajar allá y a los voluntarios que me ayudaron durante el muestreo. A la gente <strong>de</strong>Ecopar, por darme un espacio entre ellos, por su ayuda y por todos los buenos momentosque pasé en su compañía. A toda la gente que hizo <strong>de</strong> mi estadía en Ecuador, unatemporada tan agradable.Agra<strong>de</strong>zco a la gente <strong>de</strong>l Laboratorio <strong>de</strong> Botánica y Sistemática: Santiago Madriñán,Alfredo Navas, Juliana Chacón, Rafael Arévalo y Jorge Salgado. A todos los que mecolaboraron con sus opiniones, me asesoraron con bibliografía y programas estadísticos:Santiago Madriñán, Jason Rauscher, Robert Hofste<strong>de</strong>, Adolfo Amézquita, Chris James,Julio Betancur, Oscar Ramos, Rafael Arévalo, Alfredo Navas y Ecopar.Por último a la Universidad <strong>de</strong> Los An<strong>de</strong>s, por la formación académica. A mi familia porapoyarme en el curso <strong>de</strong> mi carrera y en esta última etapa, especialmente a mis papás, mishermanos y a Germán Andra<strong>de</strong>. A mis amigos, por darme una mano cuando más lanecesité y a Rafael Arévalo por su compañía y por haber recorrido este camino conmigo.iv
RESUMENSe estudió la variación morfométrica y <strong>de</strong>mográfica <strong>de</strong> Espeletia pycnophylla en ungradiente altitudinal, en la Provincia <strong>de</strong> Carchi, Ecuador. Dentro <strong>de</strong> la provincia, serealizaron mediciones en tres localida<strong>de</strong>s diferentes: Reserva biológica Guan<strong>de</strong>ra, y<strong>de</strong>ntro <strong>de</strong>l Páramo El Angel, las localida<strong>de</strong>s <strong>de</strong> la Asociación Agrícola y Gana<strong>de</strong>ra 23 <strong>de</strong>Julio y la Zona <strong>de</strong> las Lagunas <strong>de</strong> El Vola<strong>de</strong>ro. Se realizaron mediciones <strong>de</strong> la altura total,el diámetro <strong>de</strong> la roseta y el número <strong>de</strong> capítulos por inflorescencia; para el estudio<strong>de</strong>mográfico, se tuvo en cuenta la variación altitudinal en la <strong>de</strong>nsidad <strong>de</strong> juveniles yadultos, y se construyeron tablas <strong>de</strong> vida estáticas. Se encontró que el único caractermorfológico que se correlacionó con la altitud, fue el número <strong>de</strong> capítulos porinflorescencia; la ten<strong>de</strong>ncia observada fue hacia una disminución en el número <strong>de</strong>capítulos al aumentar la elevación. Las otras dos medidas —altura y diámetro <strong>de</strong> laroseta— se comportan <strong>de</strong> manera diferente en cada una <strong>de</strong> las tres localida<strong>de</strong>s, y por lotanto no se observa una ten<strong>de</strong>ncia altitudinal inequívoca. Con respecto al análisis<strong>de</strong>mográfico, las <strong>de</strong>nsida<strong>de</strong>s <strong>de</strong> adultos y juveniles no mostraron ninguna ten<strong>de</strong>nciaaltitudinal para la especie. Al parecer las poblaciones se están viendo afectadas por otrascondiciones diferentes al gradiente altitudinal, como historia <strong>de</strong> disturbio local, efecto <strong>de</strong>competencia, entre otros. Las tablas <strong>de</strong> vida fueron muy similares para cada altitud y paracada localidad. No se encontraron diferencias en las curvas <strong>de</strong> supervivencia, por lo queparece que la especie tiene un comportamiento <strong>de</strong>mográfico muy estable. Se analizótambién, el efecto <strong>de</strong> comunidad, es <strong>de</strong>cir <strong>de</strong>l porcentaje <strong>de</strong> cobertura vegetal y el efecto<strong>de</strong> otras especies dominantes en la morfometría y <strong>de</strong>mografía <strong>de</strong> las rosetas. Se observóque el porcentaje <strong>de</strong> cobertura no tuvo efecto en la altura <strong>de</strong> las rosetas; la altura <strong>de</strong> lapaja, Calamagrostis sp., se correlacionó con la altura y el diámetro <strong>de</strong> las rosetas, lo quepue<strong>de</strong> indicar una misma historia <strong>de</strong> disturbio, mismas condiciones <strong>de</strong> microclima oedafológicas en una cierta localidad, o competencia. Con respecto al efecto <strong>de</strong> otrasespecies (altura <strong>de</strong> Calamagrostis sp. y <strong>de</strong> Puya sp.) en la <strong>de</strong>nsidad <strong>de</strong> individuos <strong>de</strong>Espeletia, se observó que existen diferentes ten<strong>de</strong>ncias en cada localidad, aunque engeneral, existe un efecto en la reducción <strong>de</strong>l número <strong>de</strong> juveniles.v
LISTA DE TABLASTabla 1.Tabla 2.Tabla 3.Tabla 4.Tabla 5.Tabla 6.Tabla 7.Tabla 8.Descripción <strong>de</strong>l rango altitudinal y el área muestreada <strong>de</strong> cada una <strong>de</strong> laslocalida<strong>de</strong>s estudiadas.Resultados <strong>de</strong> los coeficientes <strong>de</strong> correlación <strong>de</strong> Spearman para los datos <strong>de</strong>Morfometría.Resultados <strong>de</strong> la prueba <strong>de</strong> Kruskal Wallis para los datos <strong>de</strong> Morfometría yCobertura Vegetal.Resultados <strong>de</strong> los coeficientes <strong>de</strong> correlación <strong>de</strong> Spearman para los datos <strong>de</strong>Estructura <strong>de</strong> Población.Resultados <strong>de</strong> la prueba <strong>de</strong> Kruskal Wallis para los datos <strong>de</strong> EstructuraPoblacional.Tabla <strong>de</strong> vida para vida para toda la especie, utilizando los datos obtenidos <strong>de</strong>las tres localida<strong>de</strong>s.Resultados <strong>de</strong> los coeficientes <strong>de</strong> correlación <strong>de</strong> Spearman para los datos <strong>de</strong>lEfecto <strong>de</strong> comunidad.Resultados <strong>de</strong> la prueba <strong>de</strong> Kruskal Wallis para los datos <strong>de</strong>l Efecto <strong>de</strong> lacomunidad.vi
LISTA DE FIGURASFigura 1. Representa la variación en los tipos <strong>de</strong> bosque y sus alturas, al aumentar elgradiente altitudinal. El Bosque Montano Bajo, como su nombre lo indica,ocurre a bajas elevaciones, mientras que el Páramo se encuentra en laselevaciones mayores.Figura 2. Mapa base <strong>de</strong> la Reserva <strong>de</strong> Guan<strong>de</strong>raFigura 3. Localida<strong>de</strong>s <strong>de</strong> la Asociación Agrícola y Gana<strong>de</strong>ra 23 <strong>de</strong> Julio, y la zona <strong>de</strong>las lagunas <strong>de</strong>l Vola<strong>de</strong>ro.Figura 4. Descripción <strong>de</strong> las partes básicas <strong>de</strong> un Espeletia, y medidas morfométricasque se tomaron en el estudio.Figura 5. Relación entre las alturas <strong>de</strong> la roseta y la altitud en la zona <strong>de</strong> la 23.Figura 6. Relación entre las alturas <strong>de</strong> las plantas <strong>de</strong> la localidad Guan<strong>de</strong>ra y la altitud.Figura 7. Relación entre las alturas <strong>de</strong> las plantas <strong>de</strong> El Vola<strong>de</strong>ro y la altitud.Figura 8. Variación <strong>de</strong> la diez alturas mayores por altitud al aumentar la elevación, parala Zona <strong>de</strong> la 23.Figura 9. Variación <strong>de</strong> la diez alturas mayores por altitud al aumentar la elevación, en elsitio <strong>de</strong> estudio Guan<strong>de</strong>ra.Figura 10. Variación <strong>de</strong> la diez alturas mayores por altitud al aumentar la elevación, enEl Vola<strong>de</strong>ro.Figura 11. Valores <strong>de</strong> altura máxima en cada cota altitudinal, para las localida<strong>de</strong>s don<strong>de</strong>se realizó el estudio.Figura 12. Relación entre el diámetro <strong>de</strong> la roseta y la altitud, en la Zona <strong>de</strong> la 23.Figura 13. Relación entre el diámetro <strong>de</strong> la roseta y la altitud, en la localidad Guan<strong>de</strong>ra.Figura 14. Relación entre el diámetro <strong>de</strong> la roseta y la altitud, en El Vola<strong>de</strong>ro.Figura 15. Variación en los diez diámetros mayores por altitud, al aumentar la elevación,en localidad <strong>de</strong> La 23.Figura 16. Variación en los diez diámetros mayores por altitud, al aumentar la elevación,en la localidad Guan<strong>de</strong>ra.Figura 17. Cambio en los diez diámetros mayores por altitud, al aumentar la elevación, enEl Vola<strong>de</strong>ro.vii
Figura 18. Variación <strong>de</strong>l número <strong>de</strong> capítulos por inflorescencia en un gradientealtitudinal, para la localidad <strong>de</strong> la 23.Figura 19. Variación <strong>de</strong>l número <strong>de</strong> capítulos por inflorescencia en un gradientealtitudinal, para la localidad <strong>de</strong>l Vola<strong>de</strong>ro.Figura 20. Cambio en la <strong>de</strong>nsidad <strong>de</strong> adultos <strong>de</strong> acuerdo a la altitud, en las treslocalida<strong>de</strong>s <strong>de</strong> estudio.Figura 21. Variación en la <strong>de</strong>nsidad <strong>de</strong> juveniles con respecto a la altitud, en la Zona <strong>de</strong> la23.Figura 22. Variación en la <strong>de</strong>nsidad <strong>de</strong> juveniles con la altitud para la localidadGuan<strong>de</strong>ra.Figura 23. Variación en la <strong>de</strong>nsidad <strong>de</strong> juveniles con la altitud para El Vola<strong>de</strong>ro.Figura 24. Relación entre las clases <strong>de</strong> altura y la proporción <strong>de</strong> individuos, p, en las treslocalida<strong>de</strong>s <strong>de</strong> estudio.Figura 25. Se muestran las líneas <strong>de</strong> ten<strong>de</strong>ncia para las 3 localida<strong>de</strong>s <strong>de</strong> páramo.Figura 26. Relación entre las clases <strong>de</strong> altura y el logaritmo <strong>de</strong> la proporción (log1000lx)para la especie E. pychnopylla.Figura 27. Relación entre las clases <strong>de</strong> altura y la proporción <strong>de</strong> supervivientes (lx), en lastres localida<strong>de</strong>s.Figura 28. Relación entre las clases <strong>de</strong> altura y la tasa <strong>de</strong> mortalidad, qx, en las treslocalida<strong>de</strong>s <strong>de</strong> estudio, y a nivel <strong>de</strong> la especie.Figura 29. Relación entre la altura <strong>de</strong> la paja, Calamagrostis sp. y la altura <strong>de</strong> la roseta,en los tres sitios <strong>de</strong> estudio.Figura 30. Relación entre la altura <strong>de</strong> Calamagrostis sp. y el diámetro <strong>de</strong> la roseta, en lostres sitios <strong>de</strong> estudio.Figura 31. Relación entre la altura <strong>de</strong> la paja, Calamagrostis sp., y el número <strong>de</strong> juvenilesen las tres localida<strong>de</strong>s <strong>de</strong> estudio.Figura 32. Relación entre la cantidad <strong>de</strong> Puya sp., y el número <strong>de</strong> juveniles en las treslocalida<strong>de</strong>s.Figura 33. Mo<strong>de</strong>lo <strong>de</strong>l posible comportamiento <strong>de</strong> la estructura <strong>de</strong> eda<strong>de</strong>s <strong>de</strong> la población<strong>de</strong> Espeletia killipii sometida a disturbios continuos.viii
1. <strong>REVISION</strong> <strong>MONOGRAFICA</strong>1.1 Ecosistema páramo1.1.1 DescripciónLa palabra páramo viene <strong>de</strong>l latín paramus y fue utilizada por los conquistadoresespañoles para <strong>de</strong>signar las áreas <strong>de</strong>l norte <strong>de</strong> los An<strong>de</strong>s que eran altas, inhóspitas, frías ycon vientos y lluvias constantes, que al parecer les recordaba a la Península Ibérica(Luteyn, 1999). El ecosistema <strong>de</strong> páramo, hace parte <strong>de</strong> las montañas tropicales <strong>de</strong>América y se encuentra distribuido <strong>de</strong> manera discontinua entre las latitu<strong>de</strong>s <strong>de</strong> 11°N y8°S (Luteyn, 1999). Está concentrado especialmente en la parte norte <strong>de</strong> Sur América:Venezuela, Colombia y Ecuador, con algunas zonas en Centroamérica y el norte <strong>de</strong> Perú.Es el último cinturón <strong>de</strong> vegetación y se encuentra, generalmente, sobre el límite superior<strong>de</strong>l bosque húmedo altomontano. Al encontrarse por encima <strong>de</strong> la vegetación <strong>de</strong> bosqueandino, el ecosistema se ve fragmentado, formando islas. Estas islas, como las <strong>de</strong>scribióCarlquist (1974), son equiparables con las oceánicas, pues las partes frías y altas seencuentran aisladas unas <strong>de</strong> otras, por mares <strong>de</strong> bosques tropicales en las partes bajas. Lasislas encontradas en los páramos se comportan como verda<strong>de</strong>ras islas en dos aspectosprincipales: el área <strong>de</strong> la isla pue<strong>de</strong> <strong>de</strong>terminar el número aproximado <strong>de</strong> especies que seencuentran en ella, y la distancia entre islas pue<strong>de</strong> pre<strong>de</strong>cir el número <strong>de</strong> en<strong>de</strong>mismos(Vuilleumier, 1970). Las islas representan una oportunidad <strong>de</strong> estudiar varios procesosevolutivos, como radiaciones adaptativas, por ser entes geográficos con límites <strong>de</strong>finidos,por poseer flujo genético reducido con otras islas, por su tamaño pequeño y por poseeraltos niveles <strong>de</strong> en<strong>de</strong>mismo (Emerson, 2002).Para llegar a esta distribución actual, los páramos han sufrido una historia compleja ydinámica, don<strong>de</strong> cada isla pue<strong>de</strong> ser resultado <strong>de</strong> un proceso diferente (Van <strong>de</strong>r Hammeny Cleef, 1986). Los ciclos <strong>de</strong> glaciaciones ocurridos en el Pleistoceno tuvieron un granimpacto en las condiciones <strong>de</strong> la vegetación llevando a expansiones y contracciones <strong>de</strong>lecosistema, favoreciendo la especiación alopátrica (Simpson y Todzia, 1990). Elaislamiento <strong>de</strong> islas y su eventual contacto, la diversidad <strong>de</strong> nichos y ambientes, y la
Sánchez, 2004. Morfometría y Demografía <strong>de</strong> E. pycnophylla. 2migración <strong>de</strong> especies <strong>de</strong>s<strong>de</strong> diferentes lugares han contribuido a la evolución <strong>de</strong> estaflora (Van <strong>de</strong> Hammen y Cleef, 1986), y aunque este ecosistema es geológicamentereciente, 2–4 millones <strong>de</strong> años, se consi<strong>de</strong>ra como uno <strong>de</strong> los ecosistemas <strong>de</strong> montaña quemuestran mayor diversidad y en<strong>de</strong>mismo en el mundo (Luteyn, 1999). Se propone que laflora <strong>de</strong> páramo tiene orígenes múltiples por adaptación <strong>de</strong> elementos tropicales <strong>de</strong> bajaselevaciones, por inmigración <strong>de</strong> las zonas templadas o por especiación <strong>de</strong> elementosautóctonos (Van <strong>de</strong>r Hammen y Cleef, 1986, Luteyn, 1999).El aislamiento geográfico <strong>de</strong> estas islas, su alto grado <strong>de</strong> en<strong>de</strong>mismo y diversidad, hacen<strong>de</strong>l ecosistema un lugar biogeográficamente único (Luteyn, 1999). En el páramo se pue<strong>de</strong>estudiar la evolución en un sentido amplio, don<strong>de</strong> se observan cambios ambientales,evolución <strong>de</strong> taxa, inmigración <strong>de</strong> taxa y adaptaciones.Según Cuatrecasas (en Luteyn,1999; Van <strong>de</strong>r Hammen y Cleef, 1986), el páramo, enrealidad está constituido por 3 zonas según la altitud y la estructura <strong>de</strong> la vegetación.Estas 3 zonas se <strong>de</strong>nominan: subpáramo, el páramo propiamente dicho y superpáramo.Así mismo el páramo se pue<strong>de</strong> clasificar <strong>de</strong>pendiendo <strong>de</strong> las características topográficas,la exposición al viento y las condiciones climáticas, en páramo atmosféricamente húmedoy atmosféricamente seco. Según estas características las elevaciones a las que ocurre cadazona varían: el límite entre el bosque y el páramo pue<strong>de</strong> ser más alto en la parte húmedaque en la seca (Van <strong>de</strong>r Hammen y Cleef, 1986).A<strong>de</strong>más <strong>de</strong> la ubicación topográfica <strong>de</strong>l páramo, el ecosistema presenta condicionesambientales drásticas; un conjunto <strong>de</strong> factores climáticos, edafológicos ygeomorfológicos condicionan la presencia <strong>de</strong> los organismos que han logrado adaptarse aestos ambientes extremos (Vargas, 1998a).Dentro <strong>de</strong> las características particulares <strong>de</strong>l ecosistema se encuentra una baja variaciónen la temperatura media entre estaciones, una gran variación en las condicionesclimáticas diarias, altos niveles <strong>de</strong> radiación UV, fuertes vientos, baja presiónatmosférica, bajos niveles en la presión parcial <strong>de</strong> oxígeno y la presencia <strong>de</strong> fuertes
Sánchez, 2004. Morfometría y Demografía <strong>de</strong> E. pycnophylla. 3heladas (Vargas, 1998a; Luteyn, 1999). El patrón <strong>de</strong> “verano cada día e invierno cadanoche” (Hedberg en Luteyn, 1999) es una <strong>de</strong> las características más <strong>de</strong>terminantes y laque mayor impacto tiene en la evolución <strong>de</strong> las plantas tropicales <strong>de</strong> este ecosistema(Smith y Young, 1987). Los suelos constituyen otra característica interesante: sonrelativamente jóvenes y poco <strong>de</strong>sarrollados, almacenan gran<strong>de</strong>s cantida<strong>de</strong>s <strong>de</strong> agua, sonpobres en nutrientes y su pH es ácido (Baruch, 1979, Vargas, 1998a). Todas estascondiciones abióticas llevan, como consecuencia, a que el crecimiento y <strong>de</strong>sarrollo sealento, la productividad primaria sea baja, a una lenta <strong>de</strong>scomposición <strong>de</strong> biomasa muerta,una difícil absorción radicular <strong>de</strong> nutrientes, severas quemaduras en las plantas y a que lasucesión tome mucho tiempo (Castaño, 1995; Luteyn, 1999).Varias <strong>de</strong> las adaptaciones morfológicas y fisiológicas que se encuentran en el páramo, sehan <strong>de</strong>sarrollado como un medio <strong>de</strong> escape o como una solución para tolerar lasfluctuaciones extremas en el clima diario (Smith y Young, 1987). Las formas <strong>de</strong>crecimiento constituyen un ejemplo <strong>de</strong> las adaptaciones que han sufrido diferentesespecies <strong>de</strong> plantas, como respuesta a las presiones que ejerce el ecosistema. Dentro <strong>de</strong>estas formas <strong>de</strong> crecimiento se encuentran las rosetas (acaules o caulescentes), lospajonales, cojines, arbustos micrófilos y enanos, y las geofitas (Hedberg y Hedberg,1979; Luteyn, 1999).Sin embargo, estas formas <strong>de</strong> crecimiento no son exclusivas <strong>de</strong>l páramo. Resultainteresante la recurrencia en el hábito <strong>de</strong> roseta y la correlación entre la fisiología y lashistorias <strong>de</strong> vida en muchos lugares <strong>de</strong>l Nuevo y Viejo mundo (Sur América, Este <strong>de</strong>Africa y Hawai). Esto soporta la i<strong>de</strong>a <strong>de</strong> evolución convergente, pues estas característicasson aparentemente adaptaciones para ambientes tropicales <strong>de</strong> alta montaña (Knox yPalmer, 1995). Ejemplos <strong>de</strong> evolución convergente en rosetas gigantes, se encuentran enlas lobelias (Lobeliae), y los Dendrosenecio <strong>de</strong> Africa, la alianza <strong>de</strong> los “Silverswords”en Hawai y en Espeletia, y Puya, en América.Como se pue<strong>de</strong> observar, el páramo es un ecosistema muy interesante a nivel científico;representa una oportunidad para estudiar y enten<strong>de</strong>r varios procesos biológicos. A su vez,
Sánchez, 2004. Morfometría y Demografía <strong>de</strong> E. pycnophylla. 4tiene un papel fundamental para la regulación <strong>de</strong> la hidrología regional y constituye unafuente <strong>de</strong> agua potable. Sin embargo se encuentra amenazado por las activida<strong>de</strong>shumanas, que han generado una gran cantidad <strong>de</strong> disturbios que alteran el ecosistema.1.1.2 Disturbio antrópicoA pesar <strong>de</strong> que la presencia humana en las zonas andinas se remonta a unos 18,000 añosantes <strong>de</strong>l presente (AP), la ocupación humana y el establecimiento en el páramo sonhechos relativamente recientes (González y Cár<strong>de</strong>nas, 1995). Al parecer, los páramos seexplotan <strong>de</strong>s<strong>de</strong> hace más <strong>de</strong> 1400 años por parte <strong>de</strong> comunida<strong>de</strong>s indígenas, pero el usoprincipal era como lugar sagrado, como corredores o áreas <strong>de</strong> cacería (Vargas, 1998b;Luteyn, 1999). Sin embargo, con la llegada <strong>de</strong> los españoles, en el siglo XVI, el impactosobre el ecosistema aumentó significativamente. La introducción <strong>de</strong> animales exóticoscomo vacas, caballos, ovejas y cabras, y la influencia <strong>de</strong>l sistema agrícola europeo generóla principal fuerza <strong>de</strong> <strong>de</strong>strucción <strong>de</strong> los páramos y bosques (González y Cár<strong>de</strong>nas, 1995;Vargas, 1998b; Luteyn, 1999).A partir <strong>de</strong>l siglo XVIII, la utilización económica <strong>de</strong>l páramo se volvió extensiva,aumentando el número <strong>de</strong> cabezas <strong>de</strong> ganado, la ocupación humana y por lo tanto lanecesidad <strong>de</strong> cultivos. Este proceso se aceleró enormemente durante los últimos 50 años,don<strong>de</strong> la presión <strong>de</strong>mográfica y las reformas agrarias, favorecieron una ocupación aúnmás extensiva (González y Cár<strong>de</strong>nas, 1995; Luteyn, 1999).Como se pue<strong>de</strong> observar, la mayor fuerza <strong>de</strong>structora <strong>de</strong>l ecosistema o el mayorproblema, es <strong>de</strong>mográfico. Debido al aumento en número y <strong>de</strong>nsidad <strong>de</strong> individuoshumanos, ha surgido la necesidad <strong>de</strong> ocupar nuevos lugares para suplir susrequerimientos. Esta ocupación va acompañada, principalmente, por un aumento <strong>de</strong>ganado, mayores áreas <strong>de</strong> cultivo y <strong>de</strong> métodos para aumentar la calidad <strong>de</strong> alimento paralos animales. A esto se le pue<strong>de</strong> sumar la tala <strong>de</strong> bosque para ma<strong>de</strong>ra, la cacería, el uso <strong>de</strong>fertilizantes en los cultivos, apertura <strong>de</strong> carreteras, minería, turismo, la plantación <strong>de</strong>especies exóticas como el pino, etc.
Sánchez, 2004. Morfometría y Demografía <strong>de</strong> E. pycnophylla. 5A pesar <strong>de</strong> existir una gran cantidad <strong>de</strong> disturbios, los que causan el mayor impacto sobreel ecosistema <strong>de</strong> páramo son el uso indiscriminado <strong>de</strong>l fuego y la gana<strong>de</strong>ría (Hofste<strong>de</strong>,1995; Vargas, 1998b; Hofste<strong>de</strong>, 2001). El fuego se utiliza en el páramo, para dospropósitos (Hofste<strong>de</strong>, 1995; Luteyn, 1999). El primero es como medio <strong>de</strong> <strong>de</strong>forestación,para proveer nuevas áreas <strong>de</strong> pastizal para el ganado, o para ampliar las zonas <strong>de</strong> cultivo.La segunda razón es para proveer al ganado, nuevos brotes, ricos en nutrientes.Como se pue<strong>de</strong> ver, el uso <strong>de</strong>l fuego suele ir acompañado <strong>de</strong> la gana<strong>de</strong>ría, causandodisturbios a diferentes niveles según lo <strong>de</strong>scribió Vargas (1998b). El autor <strong>de</strong>fine tresniveles:- Nivel geomorfológico: las quemas generalmente se realizan en la época seca; elfuego elimina la cobertura vegetal, haciendo que el suelo que<strong>de</strong> sujeto a<strong>de</strong>secación y a congelación en las noches. Esto genera procesos <strong>de</strong> solifluxión, locual favorece la erosión (Hofste<strong>de</strong>, 1995; Castaño, 1995; Luteyn, 1999; Hofste<strong>de</strong>,2001). Luego, al llegar el invierno, las pisadas <strong>de</strong>l ganado crean charcos, queafectan las características hidráulicas <strong>de</strong>l suelo (Castaño, 1995; Hofste<strong>de</strong>, 2001).El suelo se compacta, afectando el <strong>de</strong>sarrollo <strong>de</strong> la vegetación, y procesos <strong>de</strong><strong>de</strong>scomposición y respiración <strong>de</strong> éste (Singh y Grupta en Vargas, 1998b). Elefecto combinado <strong>de</strong> pastoreo y quema <strong>de</strong>struye el proceso <strong>de</strong> formación <strong>de</strong>cavida<strong>de</strong>s <strong>de</strong>l suelo, haciendo que éste no pueda retener tanta agua (Luteyn,1999). Los cambios en hidromorfia, producen <strong>de</strong>svíos en los cursos naturales <strong>de</strong>agua, formando áreas pantanosas (Vargas, 1998b).- Nivel <strong>de</strong>l subsistema suelo-vegetación: por las bajas temperaturas típicas <strong>de</strong>lecosistema, la <strong>de</strong>scomposición <strong>de</strong> materia orgánica es lenta, por falta <strong>de</strong> insectos yotros invertebrados (termitas, hormigas, escarabajos y gusanos), y por la bajaactividad metabólica <strong>de</strong> hongos y bacterias (Janzen, 1973; Smith, 1980; Vargas,1998b). La acumulación <strong>de</strong> materia orgánica aumenta la capacidad <strong>de</strong> retención<strong>de</strong> agua por parte <strong>de</strong>l suelo, pero lo hace más blando y por lo tanto, susceptible alas pisadas <strong>de</strong>l ganado. Al realizar quemas recurrentes y pastoreo intensivo, lasucesión se <strong>de</strong>tiene y se da mortalidad selectiva <strong>de</strong> algunas plantas, creando
Sánchez, 2004. Morfometría y Demografía <strong>de</strong> E. pycnophylla. 6parches (Hobbs y Huenneke, 1992). Esto genera cambios en la estructurapoblacional, y en muchos casos, si el nivel <strong>de</strong> disturbio es alto, la estructuravegetacional cambia, hacia pastizales y muchas especies <strong>de</strong> plantas se pier<strong>de</strong>n(Vargas, 1998; Luteyn, 1999; Hofste<strong>de</strong>, 2001). Se altera la diversidad tanto anivel local (alfa) como a nivel regional (beta) (Janzen, 1973; Luteyn, 1999).- Nivel <strong>de</strong> procesos: con las quemas se alteran los ciclos <strong>de</strong> nutrientes. Muchos <strong>de</strong>los nutrientes se retienen en la necromasa, la cual los recicla. Al quemar, seliberan masivamente nutrientes, <strong>de</strong> los cuales, sólo una pequeña fracción esutilizable, aumentando temporalmente la productividad (Vargas, 1998b). Conrespecto a los suelos, al realizar quemas reiteradas y pastoreo intensivo, se generauna <strong>de</strong>scomposición rápida, <strong>de</strong>bido a que el suelo se vuelve seco, y pier<strong>de</strong> lacapacidad <strong>de</strong> retener nutrientes (Hofste<strong>de</strong>, 1995; Verweij, 1995). A corto plazoéstos son utilizables, sin embargo se pier<strong>de</strong>n rápido <strong>de</strong>l suelo, por erosión,volatilización o inmovilización (Luteyn, 1999). Las quemas, disminuyen ysimplifican los ciclos <strong>de</strong> nutrientes; el pastoreo, reduce sus reservas en lavegetación, y posteriormente causa redistribución <strong>de</strong>sproporcionada, por las hecesy la orina (Vargas, 1998b).A nivel general, las ventajas <strong>de</strong> la quema y el pastoreo son a corto plazo. A largo plazo setienen <strong>de</strong>sventajas en la retención <strong>de</strong> agua y nutrientes, así como en la eventual pérdida<strong>de</strong> diversidad y cambios en las estructuras poblacionales. Según Verweij (1995), eltiempo mínimo para que se recupere el ecosistema luego <strong>de</strong> una quema, es <strong>de</strong> 3 a 10años; sin embargo, la frecuencia suele ser <strong>de</strong> 2–4 años (Hofste<strong>de</strong>, 2001). Con elincremento <strong>de</strong> la población humana y su impacto <strong>de</strong>mográfico, la frecuencia <strong>de</strong> lasquemas está aumentando más allá <strong>de</strong> lo que el ecosistema pue<strong>de</strong> resistir haciendo que nopueda recuperarse en su totalidad, lo que genera gran<strong>de</strong>s inquietu<strong>de</strong>s a nivel <strong>de</strong>conservación. A<strong>de</strong>más se ha visto que el disturbio promueve la invasión por parte <strong>de</strong>especies no nativas e invasoras; a su vez, cambios en el tamaño, frecuencia y tipo <strong>de</strong>disturbio, pue<strong>de</strong> hacer que las especies nativas no puedan adaptarse o establecerse, y porlo tanto se vea afectada la diversidad y composición <strong>de</strong>l ecosistema (Hobbs y Huenneke,1992).
Sánchez, 2004. Morfometría y Demografía <strong>de</strong> E. pycnophylla. 7Dentro <strong>de</strong>l ecosistema <strong>de</strong> páramo, existen algunas especies <strong>de</strong> plantas que son notablespor su dominancia. Al estar el ecosistema afectado por el disturbio, las poblaciones seven afectadas a su vez, y en muchos lugares, se ha perdido la cobertura por parte <strong>de</strong> éstas.Dentro <strong>de</strong> las especies se encuentran diferentes especies <strong>de</strong> pastos y <strong>de</strong> Espeletia.1.2 El género EspeletiaEn el páramo, una <strong>de</strong> las especies más típicas y emblemáticas son las pertenecientes algénero Espeletia. Este género, llamado comúnmente frailejón, fue nombrado porHumboldt y Bonpland en 1809, en honor a Don José <strong>de</strong> Espeletia virrey <strong>de</strong>l Nuevo Reino<strong>de</strong> Granada (Standley, 1915).Taxonómicamente, el género hace parte <strong>de</strong> la familia Asteraceae, tribu Heliantheae,subtribu Espeletiinae (Cuatrecasas, 1986). El grupo Espeletiinae compren<strong>de</strong> ocho génerosy más <strong>de</strong> 100 especies endémicas <strong>de</strong> Venezuela, Colombia y Ecuador. Los frailejonesocurren generalmente, por encima <strong>de</strong> los 3000 m <strong>de</strong> altitud y presenta diferentes formas<strong>de</strong> crecimiento relacionadas con el tipo <strong>de</strong> vegetación don<strong>de</strong> se encuentran (Cuatrecasas,1979). Según Smith y Koch (1935), Espeletia no tiene especies ampliamente distribuidas,pero cada grupo <strong>de</strong> páramos se encuentra habitado por una o más especies. A pesar <strong>de</strong>tener una amplia distribución en los páramos <strong>de</strong> estos tres países, y ser un elemento tan<strong>de</strong>stacado en el ecosistema, existen algunos cuestionamientos con respecto a su origen.El origen <strong>de</strong>l género Espeletia es incierto: se propone que un taxón proto-Espeletia sufrióun proceso <strong>de</strong> radiación adaptativa acelerado, colonizando todo el rango <strong>de</strong> hábitats posty periglaciales, <strong>de</strong>s<strong>de</strong> <strong>de</strong>siertos fríos hasta pantanos permanentes, dando lugar a una granvariedad <strong>de</strong> formas <strong>de</strong> vida y patrones <strong>de</strong> reproducción (Monasterio y Sarmiento, 1991).Este taxón ancestral, al parecer tiene su origen en el inicio <strong>de</strong>l periodo glacial <strong>de</strong>lPleistoceno temprano o en el Plioceno tardío cuando las condiciones <strong>de</strong> páramo frío yhúmedo prevalecían en la Cordillera Occi<strong>de</strong>ntal <strong>de</strong> Venezuela (Cuatrecasas, 1986). Suprecursor se localizaría en los bosques <strong>de</strong> montaña tropical <strong>de</strong> Venezuela, pero suevolución se llevaría a cabo en el páramo, constituyendo un elemento autóctono,
Sánchez, 2004. Morfometría y Demografía <strong>de</strong> E. pycnophylla. 8endémico <strong>de</strong>l ecosistema. La evolución <strong>de</strong>l grupo es relativamente reciente, y se estimaque sólo llegó a Ecuador, hace unos pocos miles <strong>de</strong> años.Según Cuatrecasas (1986), el género está muy bien <strong>de</strong>finido y se diferencia claramente, anivel morfológico, <strong>de</strong> los otros géneros <strong>de</strong> la subtribu. Sin embargo, estudios molecularesrecientes (Rauscher, 2000) muestran que las relaciones <strong>de</strong>ntro <strong>de</strong> Espeletiinae no sonclaras, y que muchos <strong>de</strong> los géneros son polifiléticos y/o parafiléticos según el gen <strong>de</strong>ITS, entre ellos Espeletia. Aún así, se consi<strong>de</strong>ra el género como unidad taxonómica útil(pero no monofilética), para estudios a nivel ecológico, y fisiológico. Aún queda muchopor enten<strong>de</strong>r <strong>de</strong> las relaciones filogenéticas <strong>de</strong>l complejo <strong>de</strong> géneros, y en especial <strong>de</strong>Espeletia. Esto se <strong>de</strong>be a la gran diversidad morfológica en formas <strong>de</strong> crecimiento yespecialización ecológica, que lo hacen un bueno candidato para estudios <strong>de</strong> procesosevolutivos como especiación y adaptación, así como para estudios <strong>de</strong> adaptaciónfisiológica y dinámica poblacional, entre otros.Espeletia es un género polimórfico y con gran plasticidad fenotípica entre los individuos<strong>de</strong> la misma especie, sobre todo en un gradiente altitudinal, o según el régimen <strong>de</strong>temperatura en el que crece la planta. Existen varias formas <strong>de</strong> crecimiento <strong>de</strong>ntro <strong>de</strong>lgénero, <strong>de</strong>finidas por Cuatrecasas (1979) como caulirrósulas policárpicas que pue<strong>de</strong>n seraltas y monocaules, sésiles y rizomatosas o sésiles y tuberosas. Según Hallé, et al. (1978),la estructura básica <strong>de</strong> Espeletia se acomoda al mo<strong>de</strong>lo <strong>de</strong> Corner. El mo<strong>de</strong>lo se <strong>de</strong>finecomo crecimiento por un meristema apical producido por un eje sin ramificación, coninflorescencias laterales.Algunas características típicas <strong>de</strong>l género son: ausencia <strong>de</strong> reproducción vegetativa(Baruch y Smith, 1979; Berry y Calvo, 1994), capítulos monoicos que varían en eltamaño y número <strong>de</strong> flores (Berry y Calvo, 1989), semillas pesadas sin mecanismo <strong>de</strong>dispersión –por lo que se cree que la dispersión es por gravedad, agua y ocasionalmenteser arrastradas por vientos mo<strong>de</strong>rados o fuertes– (Cuatrecasas, 1986; Sobrevila, 1989;Berry y Calvo, 1994), número <strong>de</strong> cromosomas, N=19, mecanismo <strong>de</strong>
Sánchez, 2004. Morfometría y Demografía <strong>de</strong> E. pycnophylla. 9autoincompatibilidad esporofítico fuerte (Sobrevila, 1989; Berry y Calvo, 1989) yten<strong>de</strong>ncia a la hibridación.Las especies <strong>de</strong>l género son gregarias y poseen especializaciones para climas fríos yhúmedos. Con respecto a las adaptaciones que le han permitido tener éxito ecológico, seencuentran la dinámica <strong>de</strong> crecimiento, reproducción, y los patrones <strong>de</strong> alocación <strong>de</strong>energía y biomasa (Smith, 1980; Monasterio, 1986; Monasterio y Sarmiento, 1991).Entre estas adaptaciones, al parecer, dos han sido fundamentales: el mantenimiento <strong>de</strong>una temperatura favorable (en especial en las zonas <strong>de</strong> crecimiento) y el balance hídrico,y las características que aumentan la posibilidad <strong>de</strong> los individuos <strong>de</strong> permanecer en lossitios <strong>de</strong> colonización, como es la presencia <strong>de</strong> bancos <strong>de</strong> semillas (Guariguata y Azocar;1988; Ochoa, 1994). El método más común entre las especies, para resistir temperaturaspor <strong>de</strong>bajo <strong>de</strong>l punto <strong>de</strong> congelamiento, es el <strong>de</strong> superenfriamiento (Squeo, et al., 1991).La floración se da en la época húmeda y la transición entre la época lluviosa y la seca; ladispersión <strong>de</strong>l fruto inicia en la estación lluviosa y continua durante la época seca y lagerminación se inicia al empezar la época lluviosa. Existe una gran variabilidad en lastasas <strong>de</strong> floración interanuales, ya que la secuencia reproductiva es asincrónica e irregular(Estrada y Monasterio, 1991; Cavelier et al., 1992).Espeletia posee a nivel popular muchos usos diferentes. La resina <strong>de</strong> E. humbertii y E.neriifolia se usan para hacer incienso; E. wed<strong>de</strong>llii se usa para curar problemas <strong>de</strong>respiración; las raíces <strong>de</strong> E. schultzii para hacer mantequilla, y las hojas para empacarla(Smith, 1981). En general se <strong>de</strong>staca por sus cualida<strong>de</strong>s sudoríficas, antibronquiales yantigripales. La <strong>de</strong>cocción <strong>de</strong> la hoja sirve contra el asma y afecciones pulmonares(Torrenegra y Tellez, 1999). También se utilizan para la construcción <strong>de</strong> casas y comofuente <strong>de</strong> leña (Verweij y Kok, 1992). Sin embargo, la colección extensiva <strong>de</strong> plantaspara estos propósitos, combinado con el pastoreo, pue<strong>de</strong> haber llevado a la ausencia <strong>de</strong>Espeletia en algunas zonas, según un estudio realizado en el Parque Nacional Natural LosNevados, Colombia (Verweij y Kok, 1995). Excepto por algunos estudios (Verweij yKok, 1992; Hofste<strong>de</strong> 1995; Premauer, 1999), no existe mucha información sobre elefecto real <strong>de</strong>l fuego y pastoreo en las poblaciones <strong>de</strong> Espeletia.
Sánchez, 2004. Morfometría y Demografía <strong>de</strong> E. pycnophylla. 10Se ha observado que bajo condiciones sin disturbio antrópico, la mayor mortalidad <strong>de</strong>individuos ocurre a nivel <strong>de</strong> juveniles y plántulas, y se encuentran las rosetas <strong>de</strong> mayoraltura (Estrada y Monasterio, 1988; Hofste<strong>de</strong>, 1995; Premauer, 1999). En estascondiciones, la tasa <strong>de</strong> crecimiento, se relaciona linearmente con la altura inicial <strong>de</strong>l tallo,y varía entre 1.9 y 8.8 cm/año, según la especie (Smith, 1980; Cavelier et al., 1992;Verweij y Kok, 1992). Sin embargo, estas tasas varían significativamente entre losindividuos, bajo el efecto <strong>de</strong> disturbio. Al parecer, las tasas <strong>de</strong> crecimiento en juveniles, yla germinación, se ven estimuladas luego <strong>de</strong> una quema; pero este aumento, se pue<strong>de</strong> vercomo una respuesta a una mayor tasa <strong>de</strong> mortalidad en estas clases y en los adultos másgran<strong>de</strong>s (Smith, 1981; Verweij y Kok, 1992, Kovár, 2001). Se estima que la mortalidadpue<strong>de</strong> aumentar hasta 2 veces más que en condiciones normales (Verweij y Kok, 1992).Según Hofste<strong>de</strong> (1995), en lugares con pastoreo intensivo y fuego, la estructura <strong>de</strong> lapoblación se ve afectada (se observan bajas <strong>de</strong>nsida<strong>de</strong>s <strong>de</strong> juveniles) y se pier<strong>de</strong> un granporcentaje <strong>de</strong> la biomasa aérea total. Debido al efecto <strong>de</strong>l disturbio, se pier<strong>de</strong> una granparte <strong>de</strong> la necromasa, lo que <strong>de</strong>scubre los mersitemas apicales, y se pier<strong>de</strong> el aislamientoque provee esta capa para las altas radiaciones y las temperaturas congelantes, haciendo aEspeletia más vulnerable.Existen otros factores, adicionales al disturbio antrópico, que pue<strong>de</strong>n condicionar laestructura <strong>de</strong> una población. Como se mencionó anteriormente, el gradiente altitudinaltiene un efecto a nivel fenotípico. La altitud pue<strong>de</strong> <strong>de</strong>terminar el establecimiento <strong>de</strong> lasplántulas, la tasa <strong>de</strong> crecimiento, el éxito reproductivo, entre otros (Smith, 1980).
Sánchez, 2004. Morfometría y Demografía <strong>de</strong> E. pycnophylla. 111.3 Efecto <strong>de</strong>l gradiente altitudinal en la estructura poblacionalEl efecto <strong>de</strong>l gradiente altitudinal fue reportado por Humboldt en 1805 y ha sido unfenómeno que ha sido ampliamente estudiado en los bosques tropicales (Beard, 1955;Grubb et al., 1963; Giles, 1975; Lawton, 1982; Wilson et al., 1987; Whitmore, 1991;Run<strong>de</strong>l et al., 1994; Richards, 1996; Sarmiento y Frolich, 2002). En general la ten<strong>de</strong>nciaobservada en estos bosques, es que al ir aumentando la elevación, la altura <strong>de</strong> las plantasdisminuye formando lo que se conoce como bosques enanos o elfin woodland (Fig. 1).Cuando las altitu<strong>de</strong>s son bastante altas, se observa la <strong>de</strong>saparición <strong>de</strong> árboles altos , dandopaso a formaciones arbustivas o pequeños árboles que se <strong>de</strong>nominan krummholz en laliteratura alemana (Wilson et al., 1987).En algunos estudios realizados en bosques <strong>de</strong> Ecuador se observó que no sólo la altura <strong>de</strong>la planta se ve afectada por el gradiente altitudinal, pero también el diámetro promedio <strong>de</strong>las plantas (Grubb et al., 1963); también existen otros aspectos que cambian con laaltitud, como el tamaño <strong>de</strong> las hojas, que cambian <strong>de</strong> mesófilas a micrófilas (Giles, 1975;Whitmore, 1991) y la composición florística <strong>de</strong>l bosque en general.METROSDISMINUCIÓN EN LA ALTURA DE LAS PLANTASFIGURA 1. Representa la variación en los tipos <strong>de</strong> bosque y sus alturas, al aumentar el gradiente altitudinal.El Bosque Montano Bajo, como su nombre lo indica, ocurre a bajas elevaciones, mientras que el Páramo seencuentra en las elevaciones mayores. (Modificado <strong>de</strong> Beard, 1955)Las razón por la que ocurre una ten<strong>de</strong>ncia a la reducción <strong>de</strong> tamaño al aumentar la altitudno se entien<strong>de</strong> muy claramente. Giles (1975), propone que las posibles razones por lacual el tamaño <strong>de</strong> las plantas se ve reducido, es por una falta <strong>de</strong> nutrientes por las bajas
Sánchez, 2004. Morfometría y Demografía <strong>de</strong> E. pycnophylla. 12tasas <strong>de</strong> <strong>de</strong>scomposición, la presencia <strong>de</strong> niebla, que pue<strong>de</strong> bloquear la transpiración, lasheladas y el viento. Otros autores proponen que la temperatura es la causante <strong>de</strong> laten<strong>de</strong>ncia (Beard, 1955), el estrés mecánico, o la falta <strong>de</strong> agua. Richards (1996), proponeque es un conjunto <strong>de</strong> todos lo factores climáticos – temperatura, precipitación, humedadatmosférica, velocidad <strong>de</strong>l viento, radiación- lo que lleva a que se <strong>de</strong>n estas formaciones.Un estudio realizado en una especie <strong>de</strong> Araliaceae (Lawton, 1982), indica que la razónpor la que pueda ocurrir la formación <strong>de</strong> bosques enanos, es <strong>de</strong>bido a una adaptaciónontogenética ya sea por plasticidad fenotípica o por diferenciación genética. Se cree quela exposición constante a lugares con viento, e inclinados, <strong>de</strong>be causar en las plantas unestrés en el cual se retarda la elongación y se favorece el incremento <strong>de</strong>l diámetro <strong>de</strong>ltronco, una hipótesis conocida como tigmomorfogénesis.En las montañas tropicales, don<strong>de</strong> existe la presencia <strong>de</strong> páramo, luego <strong>de</strong> la línea <strong>de</strong>bosque se encuentra una zona <strong>de</strong>nominada subpáramo (<strong>de</strong>finida por Cuatrecasas enLuteyn,1999). Esta zona, que correspon<strong>de</strong> a la parte más baja <strong>de</strong>l páramo, es un ejemplodon<strong>de</strong> se pue<strong>de</strong> observar la ten<strong>de</strong>ncia altitudinal. Esta zona es la más difícil <strong>de</strong> <strong>de</strong>finir<strong>de</strong>bido a que se compone por varias especies encontradas en los bosques circundantes, <strong>de</strong>arbustos y elementos típicos <strong>de</strong> páramo, como pastizales. Adicionalmente, es difícil<strong>de</strong>finirla en un gradiente altitudinal, ya que el disturbio antrópico ha modificado loslímites <strong>de</strong>l bosque, o los ha <strong>de</strong>saparecido, haciendo que el subpáramo se haya extendidohorizontal y verticalmente (Luteyn, 1999). La línea <strong>de</strong>l bosque, por lo tanto, parecenhaber disminuido su altitud <strong>de</strong>bido a la interferencia humana.Algunos ejemplos <strong>de</strong> especies <strong>de</strong> bosque encontradas y que respon<strong>de</strong>n al efectoaltitudinal en la zona son: Oreopanax y Schefflera (Araliaceae), Diplostephium yGynoxys (Asteraceae), Buddleja (Buddlejaceae), Weinmannia (Cunoniaceae), Bejaria(Ericaceae), Escallonia (Escalloniaceae), Miconia (Melastomataceae), Myrsine(Myrsinaceae), Hesperomeles (Rosaceae), (Luteyn, 1999), Clusia (Clusiaceae) yGaia<strong>de</strong>ndron (Loranthaceae).
Sánchez, 2004. Morfometría y Demografía <strong>de</strong> E. pycnophylla. 13A pesar <strong>de</strong> que la ten<strong>de</strong>ncia tien<strong>de</strong> a ser muy clara para varias especies <strong>de</strong> bosque,algunas otras parecen no cumplirla. La mayoría <strong>de</strong> las especies <strong>de</strong> frailejones, tien<strong>de</strong>n atener un patrón en la altura <strong>de</strong>l tallo diferente, es <strong>de</strong>cir, que al ir aumentando el gradientealtitudinal, las especies tien<strong>de</strong>n a aumentar su tamaño. El efecto <strong>de</strong> este gradiente en laestructura poblacional y ecología <strong>de</strong> Espeletia, es un fenómeno que ha sido pocoestudiado hasta el momento (Baruch, 1979; Smith, 1980; Berry y Calvo, 1989).Smith (1980), realizó un estudio en Venezuela para observar los cambios en la estructura<strong>de</strong> la población <strong>de</strong> E. schultzii, al aumentar la altitud. Para esta especie encontró,cumpliendo la ten<strong>de</strong>ncia general <strong>de</strong> Espeletia, que al estar más cerca <strong>de</strong>l superpáramo, losindividuos aumentaban su altura total promedio y a la vez, se encontraban los individuoscon mayor altura máxima. Para po<strong>de</strong>r explicar la razón <strong>de</strong> esta ten<strong>de</strong>ncia, plantea unaserie <strong>de</strong> hipótesis para probar cuál o cuales <strong>de</strong> éstas tiene mayor peso. Dentro <strong>de</strong> estashipótesis, se encuentran:- La altura <strong>de</strong> las plantas adultas aumenta al aumentar la altitud- Al disminuir el establecimiento <strong>de</strong> plántulas a mayores altitu<strong>de</strong>s, ocurre menoscompetencia entre adultos y juveniles, aumentando la longevidad.- La cobertura <strong>de</strong> plantas vasculares disminuye al aumentar la elevación, lo queresulta en una menor competencia interespecífica y por lo tanto aumenta lalongevidad.Dentro <strong>de</strong> las excepciones en el género, Smith (1980) menciona que sólo existen 3:(1) las especies arborescentes, (2) E. wed<strong>de</strong>llii y (3) E. hartwegiana. En este momento, seconsi<strong>de</strong>raba que E. hartwegiana tenía una distribución <strong>de</strong>s<strong>de</strong> Colombia hasta Ecuador(hoy en día E. hartwegiana ha sido <strong>de</strong>smembrada en varias especies, entre ellas E.pycnophylla). Se reporta que esta especie, por condiciones propias <strong>de</strong> los lugares quehabita, tien<strong>de</strong> a violar el patrón observado para el género. Para probar la vali<strong>de</strong>z <strong>de</strong> laten<strong>de</strong>ncia observada en el género, es interesante realizar más estudios en otras especies <strong>de</strong>Espeletia, para corroborar el planteamiento <strong>de</strong> Smith. Más aún en las especies que seencuentra lejanamente emparentadas con E. schultzii (Rauscher, 2000), y en aquellas queparecen no cumplirlo, como E. pycnophylla.
Sánchez, 2004. Morfometría y Demografía <strong>de</strong> E. pycnophylla. 14A<strong>de</strong>más <strong>de</strong> la altura, el diámetro <strong>de</strong> la roseta es otro caracter que generalmente se utilizacomo medida <strong>de</strong> crecimiento. Es interesante observar el efecto <strong>de</strong> la altitud no sólo en laaltura <strong>de</strong> las rosetas, sino también en el diámetro <strong>de</strong> la roseta, para tener una i<strong>de</strong>a <strong>de</strong>cómo se ve afectado el crecimiento en general.1.4 Demografía y Crecimiento Poblacional: Tablas <strong>de</strong> vidaLas tablas <strong>de</strong> vida proveen una fuente <strong>de</strong> información importante para enten<strong>de</strong>r lospatrones <strong>de</strong> mortalidad en una <strong>de</strong>terminada población u organismo (Krohne, 1998)Muchas veces se utilizan las distribuciones por tamaño o por edad, para indicar el estado<strong>de</strong> salud <strong>de</strong> una población (Condit et al., 1998). En estos casos, si el número <strong>de</strong> juvenileses muy alto relativo al número <strong>de</strong> adultos, se dice que la población está estable o quizásen crecimiento. En el caso contrario, un número reducido <strong>de</strong> juveniles indica que lapoblación pue<strong>de</strong> estar en <strong>de</strong>clive; muchos estudios <strong>de</strong> <strong>de</strong>mografía <strong>de</strong> poblaciones son, porlo tanto, esenciales para enten<strong>de</strong>r porqué las especies <strong>de</strong>clinan. Al enten<strong>de</strong>r mejor ladinámica <strong>de</strong>mográfica <strong>de</strong> una población se pue<strong>de</strong>n diseñar programas <strong>de</strong> conservación y<strong>de</strong>cisiones <strong>de</strong> manejo <strong>de</strong> una especie (Byers y Meagher, 1997).Los estudios <strong>de</strong>mográficos son importantes en el entendimiento <strong>de</strong> la dinámica <strong>de</strong> laspoblaciones <strong>de</strong> plantas. Existen algunos estudios realizados en diferentes especies condiferentes objetivos: para enten<strong>de</strong>r el efecto <strong>de</strong> la fragmentación <strong>de</strong>l bosque en loscambios <strong>de</strong>mográficos (Jules, 1998), para enten<strong>de</strong>r la dinámica <strong>de</strong> plantas raras (Allphiny Harper, 1997; Byers y Meagher, 1997; Rudas, 1998), para enten<strong>de</strong>r la supervivencia yestructura <strong>de</strong>mográfica en plantas sometidas a disturbio continuo (Crisp y Lange, 1976;Cavelier et al., 1992), entre otros.Todos estos estudios preten<strong>de</strong>n, mediante un análisis poblacional, enten<strong>de</strong>r elcomportamiento <strong>de</strong> una especie para po<strong>de</strong>r <strong>de</strong>sarrollar estrategias <strong>de</strong> conservación, oenten<strong>de</strong>r el impacto que puedan tener factores extrínsecos.
Sánchez, 2004. Morfometría y Demografía <strong>de</strong> E. pycnophylla. 15En Espeletia existen algunos estudios para enten<strong>de</strong>r la estructura poblacional. Smith(1981), realizó un estudio en 4 especies <strong>de</strong> frailejones en Venezuela. En estas especies seobserva la misma ten<strong>de</strong>ncia <strong>de</strong>mográfica: muchos individuos en las primeras clases <strong>de</strong>tamaño, y luego una caída abrupta hacia las clases <strong>de</strong> mayor altura. Otro estudio realizadopor Cavelier et al. (1992), en Colombia, revela la misma estructura poblacional, aunquela caida entre las clases <strong>de</strong> tamaños no son tan drásticas. Por útlimo, en dos estudiosrealizados en dos especies <strong>de</strong> frailejón (E. hartwegiana y E.killipii), se pretendió enten<strong>de</strong>rel impacto <strong>de</strong>l disturbio, en el patrón <strong>de</strong> la estructura <strong>de</strong>mográfica. Verweij y Kok (1992),mostraron que a nivel <strong>de</strong>mográfico, los lugares recientemente quemados, tienen tasas <strong>de</strong>mortalidad muy altas, y que esta tasa aumenta con la altura <strong>de</strong> la roseta. El otro estudio,realizado en E. killipii (Premauer, 1999), preten<strong>de</strong> enten<strong>de</strong>r el efecto <strong>de</strong>l disturbio en ladistribución <strong>de</strong> las clases <strong>de</strong> altura. En este caso, compara un sitio sin disturbio con uno<strong>de</strong> disturbio mo<strong>de</strong>rado pero en los dos casos se han perdido clases <strong>de</strong> altura. A<strong>de</strong>más <strong>de</strong>estos estudios no existen más análisis poblacionales para el género. Tampoco existemucha información <strong>de</strong> cómo cambia la estructura <strong>de</strong>mográfica con un gradientealtitudinal.
Sánchez, 2004. Morfometría y Demografía <strong>de</strong> E. pycnophylla. 162. OBJETIVOS2.1 Objetivo generalEstudiar la variación morfométrica y la estructura <strong>de</strong>mográfíca <strong>de</strong> Espeletia pycnophyllaCuatrec, en un gradiente altitudinal, en la Provincia <strong>de</strong> Carchi, Ecuador2.2 Objetivos específicos- Determinar el efecto <strong>de</strong>l gradiente altitudinal en la variación <strong>de</strong> algunos caracteresmorfológicos, como altura, diámetro <strong>de</strong> la roseta y el número <strong>de</strong> capítulos porinflorescencia.- Determinar el efecto <strong>de</strong>l gradiente altitudinal en la <strong>de</strong>nsidad <strong>de</strong> juveniles, adultosy el total <strong>de</strong> individuos.- Construir tablas <strong>de</strong> vida para enten<strong>de</strong>r la dinámica poblacional <strong>de</strong> la especie- Determinar el efecto <strong>de</strong> la comunidad vegetal en las medidas <strong>de</strong> morfometría(altura y diámetro <strong>de</strong> la roseta), y en la <strong>de</strong>nsidad <strong>de</strong> juveniles, adultos e individuostotales.
Sánchez, 2004. Morfometría y Demografía <strong>de</strong> E. pycnophylla. 173. MATERIALES Y METODOS3.1 Espeletia pycnophylla CuatrecasasEspeletia pycnophylla, es una roseta caulescente, perenne, que se encuentra en elsurocci<strong>de</strong>nte Colombiano y en Ecuador. Posee hojas <strong>de</strong>nsamente pubescentes, anchas,obovadas-oblongas y sésiles. Las inflorescencias son <strong>de</strong>snudas, pues las hojas <strong>de</strong> la parteproximal se han perdido. Es la especie que posee la distribución más al sur, y la másreciente a nivel evolutivo (Cuatrecasas, 1986).Esta especie muestra mucha variación regional: E. pycnophylla ssp pycnophylla, seencuentra en el surocci<strong>de</strong>nte colombiano, E. pycnophylla ssp angelensis, se encuentra alnorte <strong>de</strong> Ecuador y E. pycnophylla ssp llanganatensis en la Cordillera <strong>de</strong> los Llanganates.3.2 Area <strong>de</strong> estudioEn Ecuador se encuentran diferentes tipos <strong>de</strong> páramo (Mena Vásconez y Medina, 2001);<strong>de</strong>ntro <strong>de</strong> estos tipos, se encuentra el páramo <strong>de</strong> frailejón, que se restringe a la parte norteEcuador, a las provincia <strong>de</strong> Carchi y Sucumbíos, con una pequeña población en el ParqueNacional Llanganates. En la provincia <strong>de</strong> Carchi, esta planta cubre extensiones <strong>de</strong> variosmiles <strong>de</strong> hectáreas, mientras que en el Llanganates su extensión no exce<strong>de</strong> las 5hectáreas. Los páramos <strong>de</strong>l Carchi se <strong>de</strong>nominan <strong>de</strong> frailejones pues se encuentra<strong>de</strong>nsamente poblados por la subespecie endémica Espeletia pycnophylla ssp. angelensis.Este estudio se restringió a los páramos <strong>de</strong>l norte <strong>de</strong> Ecuador, en la Provincia <strong>de</strong> Carchi.Los suelos <strong>de</strong> estos páramos son andosoles <strong>de</strong>saturados negros, y muy orgánicos en lashondonadas (Zebrowski y Sourdat, 1997). Esto quiere <strong>de</strong>cir que son jóvenes, <strong>de</strong>rivados<strong>de</strong> cenizas volcánicas, muy fértiles y húmedos. En la provincia <strong>de</strong> Carchi el cultivo máscomún es el <strong>de</strong> papa (Zebrowski y Sourdat, 1999).Dentro <strong>de</strong> esta área se utilizaron tres localida<strong>de</strong>s para tomar los datos. La primera <strong>de</strong> éstasfue la Estación biológica Guan<strong>de</strong>ra (Fig. 2), parte <strong>de</strong> la Fundación Jatun Sacha. La
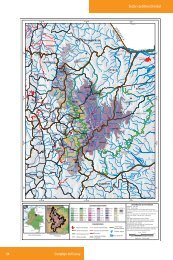
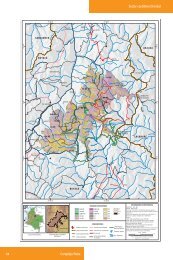
Sánchez, 2004. Morfometría y Demografía <strong>de</strong> E. pycnophylla. 18fundación posee cuatro estaciones biológicas en el Ecuador que promueven laconservación, investigación, educación, el trabajo con las comunida<strong>de</strong>s y el ecoturismo.La reserva biológica Guan<strong>de</strong>ra, creada en 1994, se encuentra ubicada en el lado oriental<strong>de</strong>l valle interandino, <strong>de</strong>l norte <strong>de</strong>l Ecuador; pertenece al Cantón Huaca, parroquia <strong>de</strong>Mariscal Sucre, a unos 11km al este <strong>de</strong>l pueblo San Gabriel. Su extensión es <strong>de</strong> 1000hectáreas y cubre altitu<strong>de</strong>s entre 3200–4000 m <strong>de</strong> altitud. La temperatura mínima seencuentra entre 0,5–7,5°C y en promedio son 4,0°C; la temperatura máxima estáalre<strong>de</strong>dor <strong>de</strong> 10,0–19,0°C y en promedio 15,0°C. La precipitación es aproximadamente1100 mm anual y la mayoría ocurre <strong>de</strong> mayo a septiembre <strong>de</strong>bido a la influenciaamazónica. A<strong>de</strong>más <strong>de</strong> la precipitación, algunos datos señalan que el agua <strong>de</strong> neblinapue<strong>de</strong> ser un fuente significante <strong>de</strong> agua en ciertos hábitats. Al parecer esta cantidad y laduración <strong>de</strong>l invierno han ido disminuyendo en las últimas siete décadas.Las otras dos localida<strong>de</strong>s se encuentran <strong>de</strong>ntro <strong>de</strong> la Reserva Ecológica El Angel, quehace parte <strong>de</strong> las zonas protegidas por el gobierno ecuatoriano. La reserva cuenta con15715Ha, y hace parte <strong>de</strong>l cantón Espejo, Parroquia La Libertad (Comuna <strong>de</strong> LaLibertad, 2001). El páramo El Angel, cuya cuenca se extien<strong>de</strong> sobre los 3100 m <strong>de</strong>altitud, se caracteriza por la presencia <strong>de</strong> Espeletia, acompañado por Festuca, Stipa yCalamagrostis. Este páramo se consi<strong>de</strong>ra una reserva ecológica nacional y se <strong>de</strong>staca porsu importancia como fuente <strong>de</strong> recursos hídricos (Fundación para el DesarrolloAgropecuario, 1998; Neill, 1999; Mena Vásconez y Medina, 2001).Según los datos registrados en la estación <strong>de</strong> Chapatán, entre los años <strong>de</strong> 1984 y 1993,(Noreste <strong>de</strong> la Reserva), la temperatura máxima se encuentra alre<strong>de</strong>dor <strong>de</strong> los 20°C, lamínima por <strong>de</strong>bajo <strong>de</strong> los 0°C y las temperaturas medias entre los 4 y 8°C. Laprecipitación anual es <strong>de</strong> 800 a 2000 mm y la humedad relativa <strong>de</strong>l 80%. La época secase inicia en mayo y dura generalmente hasta agosto, y la época lluviosa inicia enseptiembre hasta abril. Los límites <strong>de</strong> la reserva son: al norte con Colombia, el VolcánChiles, 78°30'10" longitud oeste, 0°43'55" latitud norte; al este, 77°53'40" longitud oeste,
Sánchez, 2004. Morfometría y Demografía <strong>de</strong> E. pycnophylla. 190°40'18" latitud norte, y al oeste 78°02'15" longitud oeste, 0°40'20" latitud norte(Comuna <strong>de</strong> La Libertad, 2001).La reserva se encuentra dividida en 6 zonas, 4 comunas y 2 asociaciones. De estas zonasse utilizaron dos, la Asociación Agrícola y Gana<strong>de</strong>ra 23 <strong>de</strong> Julio (Zona <strong>de</strong> La 23) y<strong>de</strong>ntro <strong>de</strong> la comuna <strong>de</strong> la Libertad, la zona <strong>de</strong> las lagunas <strong>de</strong>l Vola<strong>de</strong>ro (Fig. 3).FIGURA 2. Mapa base <strong>de</strong> la Reserva <strong>de</strong> Guan<strong>de</strong>ra (Fuente: Cartas topográficas Instituto Geográfico MilitarQuito, Realizado por: Laboratorio <strong>de</strong> SIG-ECOPAR, Datum horizontal PSAD 56, Zona 17N).
Sánchez, 2004. Morfometría y Demografía <strong>de</strong> E. pycnophylla. 20FIGURA 3. Localida<strong>de</strong>s <strong>de</strong> la Asociación Agrícola y Gana<strong>de</strong>ra 23 <strong>de</strong> Julio (<strong>de</strong>limitada en negro) y la zona<strong>de</strong> las lagunas <strong>de</strong>l Vola<strong>de</strong>ro (Fuente: Cartas topográficas Instituto Geográfico Militar Quito)
Sánchez, 2004. Morfometría y Demografía <strong>de</strong> E. pycnophylla. 213.3 MuestreoLos datos se tomaron durante los meses <strong>de</strong> Julio y Agosto <strong>de</strong>l año 2003, en cada una <strong>de</strong>las localida<strong>de</strong>s <strong>de</strong>scritas anteriormente. Se utilizaron transectos <strong>de</strong> 30m 2 (10x3),dispuestos horizontalmente a favor <strong>de</strong> la pendiente, al azar, en un intervalo <strong>de</strong> 100maltitudinales. En cada cota altitudinal, se realizaron las mediciones en cuatro transectos.Es <strong>de</strong>cir, que para cada cota, se cubrió un área <strong>de</strong> 120m 2 en total. En los transectos secontroló la altitud (registrándola con un altímetro), se tomaron los datos <strong>de</strong> la pendiente(clinómetro) y los puntos <strong>de</strong> GPS (Garmin 12xL, versión 2.01). Así mismo se anotó eltipo <strong>de</strong> vegetación dominante en el transecto (<strong>de</strong>finida como la presencia <strong>de</strong> paja, puya yarbustos) y la cobertura vegetal (Anexo 1 y 2).Según las características <strong>de</strong> cada localidad, i.e elevación <strong>de</strong> las montañas, y lasfacilida<strong>de</strong>s para tomar los datos, se muestrearon más o menos cotas altitudinales. Para elsitio <strong>de</strong> estudio, Guan<strong>de</strong>ra, se tomaron los datos en 4 cotas altitudinales, entre 3500 y3800 m <strong>de</strong> altitud, cubriendo un área total <strong>de</strong> 480m 2 . Para el Vola<strong>de</strong>ro entre 3600 y 3800cubriendo 360m 2 y para la zona <strong>de</strong> la 23 entre 3600 y 4000 m <strong>de</strong> altitud, un área <strong>de</strong>600m 2 (Tabla 1).TABLA 1. Descripción <strong>de</strong>l rango altitudinal y el área muestreada <strong>de</strong> cada una <strong>de</strong> las localida<strong>de</strong>s estudiadasLocalidadRango altitudinal(m <strong>de</strong> altitud)Area muestreada(m 2 )Zona <strong>de</strong> la 23 3600–4000 600Guan<strong>de</strong>ra 3500–3800 480El Vola<strong>de</strong>ro 3600–3800 360Las mediciones morfológicas que se realizaron en cada uno <strong>de</strong> los transectos fueron:altura máxima <strong>de</strong> la planta, diámetro <strong>de</strong> la roseta y número <strong>de</strong> capítulos porinflorescencia (para El Vola<strong>de</strong>ro y La 23), en los individuos adultos (Fig.4). Un individuoadulto se <strong>de</strong>fine según su altura, y el valor es por encima <strong>de</strong> 30cm (Verweij y Kok, 1992;
Sánchez, 2004. Morfometría y Demografía <strong>de</strong> E. pycnophylla. 22en el estudio no se observó ninguna roseta inferior a 30 cm con flores)). Los individuosmenores a esta altura se consi<strong>de</strong>raron juveniles y sólo se tuvo en cuenta su <strong>de</strong>nsidadpoblacional.FIGURA 4. Descripción <strong>de</strong> las partes básicas <strong>de</strong> un Espeletia, y medidas morfométricas que se tomaron enel estudio. (Modificado <strong>de</strong> Hofste<strong>de</strong>, 1995).3.4 Análisis <strong>de</strong> datos3.4.1 MorfometríaLos datos <strong>de</strong> morfometría se utilizan para comprobar una <strong>de</strong> las hipótesis planteadas porSmith (1980). Esta hipótesis plantea un aumento en la altura <strong>de</strong> Espeletia schultzii, alaumentar la elevación. Para esto se analizó la altura máxima por transecto y porelevación, la altura promedio <strong>de</strong> las plantas >30cm por elevación y por transecto, las 10plantas con mayor altura por transecto y por elevación.A su vez se utilizaron otros datos <strong>de</strong> morfología, que podrían verse afectados por laaltitud como el diámetro <strong>de</strong> la roseta y el número <strong>de</strong> capítulos por inflorescencia. Losanálisis para el diámetro <strong>de</strong> la roseta son los mismos que se hicieron para la altura. Conrespecto a los capítulos por inflorescencia, se analizó el número total y el promedio poraltitud.
Sánchez, 2004. Morfometría y Demografía <strong>de</strong> E. pycnophylla. 233.4.2 Estructura poblacional3.4.2.1 Densidad <strong>de</strong> individuosLa segunda hipótesis se relaciona con una disminución en el establecimiento <strong>de</strong> plántulasa mayores altitu<strong>de</strong>s, lo que resulta en menos competencia entre adultos y juvenilesaumentando la longevidad. Esto lo corrobora Smith al ver que el número <strong>de</strong> juvenilesdisminuye con la altitud. Para probar esta hipótesis, se analizaron no sólo las <strong>de</strong>nsida<strong>de</strong>s<strong>de</strong> juveniles, sino también la <strong>de</strong> los adultos y el número <strong>de</strong> individuos para ver si existealguna relación <strong>de</strong> <strong>de</strong>nsida<strong>de</strong>s con la altitud. Los análisis se realizaron teniendo en cuentael número total por transecto y por altitud.3.4.2.2 Tablas <strong>de</strong> VidaSe construyeron tablas <strong>de</strong> vida estáticas para cada transecto, para cada altitud, para lalocalidad y la especie. Estas tablas sirven para saber la estructura <strong>de</strong> una población en unmomento dado <strong>de</strong>l tiempo, es <strong>de</strong>cir, para tener una i<strong>de</strong>a general <strong>de</strong> la población; son muycomunes en los individuos iteropáreos. Para la construcción <strong>de</strong> las tablas <strong>de</strong> vida seutilizaron varias fuentes (Barbour et al., 1980; Begon et al., 1996; Caughley, 1966;Krebs, 1989; Krohne, 1998; Lowe 1969; Stiling, 1999). Los parámetros fueron:- Nx: número <strong>de</strong> individuos que viven al inicio <strong>de</strong> cada intervalo <strong>de</strong> altura x.- Proporción <strong>de</strong> individuos: número <strong>de</strong> individuos sobre el total.- lx: proporción <strong>de</strong> individuos que sobreviven al inicio <strong>de</strong> cada intervalo.- dx: número <strong>de</strong> individuos que mueren durante el intervalo x x+1- qx: tasa <strong>de</strong> mortalidad durante el intervalo x x+1Las clases se calcularon basándose en la altura <strong>de</strong> las plantas a intervalos regulares <strong>de</strong>30cm. La primera clase se <strong>de</strong>finió por las alturas entre 0-30cm (todos los individuosjuveniles), la clase 2: 31-60cm, y así sucesivamente hasta la clase 9 que se <strong>de</strong>finió comolos individuos mayores a 240cm (basado en el estudio <strong>de</strong> Verweij Y Kok, 1992; Cavelieret al., 1992; Premauer, 1999).Para el qx, es importante resaltar que siempre la última clase se toma con tasa <strong>de</strong>mortalidad <strong>de</strong> cero; el valor <strong>de</strong> cero también se utiliza cuando la tasa <strong>de</strong> mortalidad esnegativa, es <strong>de</strong>cir cuando hay más individuos en la clase siguiente.
Sánchez, 2004. Morfometría y Demografía <strong>de</strong> E. pycnophylla. 243.4.3 Efecto <strong>de</strong> la comunidad vegetal en las poblaciones <strong>de</strong> Espeletiapycnophylla3.4.3.1 Cobertura vegetalLos datos <strong>de</strong> cobertura vegetal preten<strong>de</strong>n corroborar otra <strong>de</strong> las hipótesis planteada porSmith (1980): la cobertura por plantas vasculares <strong>de</strong>crece al aumentar la altitud lo queresulta en menos competencia y por lo tanto aumento en la longevidad. Smith observóque el porcentaje <strong>de</strong> suelo <strong>de</strong>scubierto se relacionaba con la altura promedio <strong>de</strong> lasplantas en forma positiva. En este estudio se analizó el porcentaje <strong>de</strong> suelo cubierto portransecto y el promedio por cota altitudinal, en un gradiente <strong>de</strong> elevación.3.4.3.2 Efecto <strong>de</strong> otras especies vegetalesExisten otros factores que pue<strong>de</strong>n influir en la estructura morfométrica y <strong>de</strong>mográfica <strong>de</strong>la población, a<strong>de</strong>más <strong>de</strong> la altitud. La presencia <strong>de</strong> otras especies vegetales pue<strong>de</strong> causarun efecto en estas variables. Para observar este efecto se <strong>de</strong>scribió la vegetación presenteen cada uno <strong>de</strong> los transectos (Anexo 2).En general, se vio que el pajonal fue la vegetación más dominante en los transectos; encada uno <strong>de</strong> los transectos se tomó una medida general <strong>de</strong> su altura y se codificaron convalores discretos así:- 0: alturaalturaSe relacionó esta variable con las medidas <strong>de</strong> altura y diámetro <strong>de</strong> la roseta, ya quepue<strong>de</strong>n tener una influencia en el crecimiento y <strong>de</strong>sarrollo <strong>de</strong> éstas.A su vez se realizó un análisis con las <strong>de</strong>nsida<strong>de</strong>s <strong>de</strong> juveniles y adultos. En este análisisse utilizó la altura <strong>de</strong> la paja y la <strong>de</strong>nsidad <strong>de</strong> Puya sp. Para <strong>de</strong>terminar la <strong>de</strong>nsidad <strong>de</strong>puya se codificaron valores discretos <strong>de</strong> la siguiente manera: 0 la ausencia, 1 la presenciamo<strong>de</strong>rada (menos <strong>de</strong> 5 individuos), y 2 la presencia abundante.
Sánchez, 2004. Morfometría y Demografía <strong>de</strong> E. pycnophylla. 253.4.4 Análisis estadísticoTodos los análisis estadísticos se realizaron con el software SPSS 10.0 para Windows yStatistix 7.0 para Windows. Para probar la distribución <strong>de</strong> los datos, se realizó el Shapiro-Wilk Test. Se utilizaron pruebas estadísticas no paramétricas en las que los valores <strong>de</strong>P
Sánchez, 2004. Morfometría y Demografía <strong>de</strong> E. pycnophylla. 264. RESULTADOSEn la localidad <strong>de</strong> Guan<strong>de</strong>ra se encontraron 902 juveniles, 623 adultos, para un total <strong>de</strong>1525 individuos en los 480 m 2 . En La 23 se registraron 1083 juveniles, 876 adultos, paraun total <strong>de</strong> 1959 individuos en 600 m 2 . En el Vola<strong>de</strong>ro, fueron 598 juveniles, 534 adultos,haciendo un total <strong>de</strong> 1132 individuos en 360 m 2 (Tabla 1).Para la especie, en las tres localida<strong>de</strong>s muestreadas, se registraron 2583 juveniles, 2033adultos, para un total <strong>de</strong> 4616 individuos en 1440 m 2 .4.1 Morfometría4.1.1 Altura <strong>de</strong> la rosetaEn la Zona <strong>de</strong> la 23, las alturas <strong>de</strong> la roseta no mostraron una ten<strong>de</strong>ncia altitudinal clara(Fig. 5; Tabla 2). Se encontraron algunas diferencias entre las alturas para cada elevación,pero no reflejaron el gradiente (Tabla 3). Así mismo, en las otras dos localida<strong>de</strong>s, no seencontró ninguna ten<strong>de</strong>ncia altitudinal y no hubo diferencias significativas entre lasaltitu<strong>de</strong>s (Fig. 6 y 7; Tabla 2 y 3).Con respecto a las 10 alturas mayores por altitud, se encontró una ligera ten<strong>de</strong>nciapositiva en la Zona <strong>de</strong> la 23, mientras que en el Vola<strong>de</strong>ro la ten<strong>de</strong>ncia fue negativa. EnGuan<strong>de</strong>ra, aunque ningún resultado es significativo, se ve una ligera ten<strong>de</strong>ncia negativa(Figura 9; Tabla 2 y 3). Se observaron diferencias significativas en las localida<strong>de</strong>s <strong>de</strong> La23 y El Vola<strong>de</strong>ro (Fig. 8 y 10; Tabla 3); en la Zona <strong>de</strong> la 23, las diferencias altitudinalesfueron entre 3600 y 3800 m <strong>de</strong> altitud (Fig. 8), y en El Vola<strong>de</strong>ro, entre 3600 y las otrasdos cotas altitudinales (Fig. 10).Las correlaciones <strong>de</strong> la altura máxima, reflejaron una ten<strong>de</strong>ncia positiva pero nosignificativa en la Zona <strong>de</strong> la 23. En la localidad El Vola<strong>de</strong>ro, la ten<strong>de</strong>ncia fue negativa ysignificativa (Fig. 11; Tabla 2). En la localidad <strong>de</strong> La 23, la altura máxima fue <strong>de</strong> 325 cmen 4000 m <strong>de</strong> altitud; en Guan<strong>de</strong>ra, <strong>de</strong> 276 cm a 3500 m <strong>de</strong> altitud y en El Vola<strong>de</strong>ro, <strong>de</strong>348 cm a 3600 m <strong>de</strong> altitud.
Sánchez, 2004. Morfometría y Demografía <strong>de</strong> E. pycnophylla. 27En Guan<strong>de</strong>ra ninguna <strong>de</strong> las correlaciones entre altura <strong>de</strong> la planta y elevación fueronsignificativas. Sin embargo se observa que las ten<strong>de</strong>ncias son siempre negativas (Tabla2).Al analizar las altitu<strong>de</strong>s comunes <strong>de</strong> las 3 localida<strong>de</strong>s (i.e. 3600, 3700 y 3800 m <strong>de</strong>altitud), se encontraron diferencias significativas entre 3600 m <strong>de</strong> altitud y las otras doscotas (prueba <strong>de</strong> Kruskal Wallis, N=1398, χ 2 =19.0458, p
Sánchez, 2004. Morfometría y Demografía <strong>de</strong> E. pycnophylla. 28máximo por altitud. También se mostró una diferencia significativa entre los 10diámetros por altitud, entre 3600 y las altitu<strong>de</strong>s <strong>de</strong> 3700 y 3800 m <strong>de</strong> altitud (Fig. 17;Tabla 2 y 3).Al comparar las tres localida<strong>de</strong>s por sus altitu<strong>de</strong>s comunes –3600, 3700 y 3800 m <strong>de</strong>altitud– , se observó una ten<strong>de</strong>ncia no significativa, aunque hay diferencias entre 3700 m<strong>de</strong> altitud y las otras dos cotas altitudinales (Prueba <strong>de</strong> Kruskal Wallis, N= 1399,χ 2 =17.0532, p0.1) (Anexo 3).4.1.3 Capítulos por inflorescenciaEn la Zona <strong>de</strong> la 23, se ve una ten<strong>de</strong>ncia negativa significativa (Tabla 2 y 3): al aumentarla elevación se ven menos capítulos por inflorescencia. Los datos altitudinales muestrandiferencias significativas entre 3600, 3700, 3800 y 4000 m <strong>de</strong> altitud (Fig. 18). El análisis<strong>de</strong> los promedios muestra un valor <strong>de</strong> correlación más fuerte (Tabla 2 y 3, Anexo 3). Parael Vola<strong>de</strong>ro, la relación fue significativa para el análisis con el número total <strong>de</strong> datos(Tabla 2 y 3). Se encontraron diferencias entre 3700 y 3800 m <strong>de</strong> altitud (Fig. 19).El análisis <strong>de</strong> las altitu<strong>de</strong>s comunes, mostró una relación significativa y negativa (Prueba<strong>de</strong> Kruskal Wallis, N=1085, χ 2 =111.11, p
Sánchez, 2004. Morfometría y Demografía <strong>de</strong> E. pycnophylla. 29altitudinales para los juveniles, entre 3800 y 3900 m <strong>de</strong> altitud, para La 23 (Fig. 21; Tabla5).En el sitio Guan<strong>de</strong>ra, aunque ninguno <strong>de</strong> los análisis mostró relación estadísticamentesignificativa con la altitud (Tabla 4 y 5), se observan ten<strong>de</strong>ncias hacia un incremento <strong>de</strong>juveniles (Fig. 22) y el total <strong>de</strong> individuos con la altitud. En el Vola<strong>de</strong>ro tampoco seobservó ninguna relación significativa entre la <strong>de</strong>nsidad <strong>de</strong> juveniles con la altitud (Fig.23; Tabla 4 y 5). Sólo se observaron diferencias entre las altitu<strong>de</strong>s <strong>de</strong> 3700 con las <strong>de</strong>3600 y 3800 m <strong>de</strong> altitud, en el análisis <strong>de</strong> la <strong>de</strong>nsidad <strong>de</strong> individuos totales y laelevación.4.2.2 Tablas <strong>de</strong> VidaSi se compara el número <strong>de</strong> individuos en los tres sitios <strong>de</strong> estudio, se pue<strong>de</strong> observar quetienen distribuciones similares, aunque por ser los tamaños <strong>de</strong> muestra diferentes, no sonmuy apreciables. En este caso resulta mejor comparar las proporciones <strong>de</strong> individuos, yaque éstas permiten saber cual es aporte <strong>de</strong> los individuos en cada clase <strong>de</strong> altura (Fig. 24).Utilizando este parámetro se ve más claramente que las tres localida<strong>de</strong>s tienendistribuciones muy similares en cada clase <strong>de</strong> altura. La primera clase, es <strong>de</strong>cir la <strong>de</strong>juveniles, tiene la proporción más alta; luego van disminuyendo hasta la clase 5, y a partir<strong>de</strong> esta clase, los valores tien<strong>de</strong>n a estabilizarse.Las tablas <strong>de</strong> vida fueron construidas basándose en los parámetros anteriormente<strong>de</strong>scritos (Tabla 6, Anexo 4 y 5). En algunos transectos se pudo observar variación en laspendientes <strong>de</strong> las curvas <strong>de</strong> supervivencia (a 4000 m <strong>de</strong> altitud en La 23, y 3800 m <strong>de</strong>altitud en Guan<strong>de</strong>ra), así como en las cotas altitudinales (en la Zona <strong>de</strong> la 23, entre 3600y las cotas <strong>de</strong> 3700, 3800 y 4000 m <strong>de</strong> altitud); sin embargo las ten<strong>de</strong>ncias son muyconsistentes. En ninguna <strong>de</strong> las localida<strong>de</strong>s se observó que la estructura <strong>de</strong>mográficavaríe con el gradiente altitudinal.Al unir los datos <strong>de</strong> las altitu<strong>de</strong>s <strong>de</strong> cada localidad, no se vieron diferencias entre laspendientes <strong>de</strong> las líneas <strong>de</strong> Guan<strong>de</strong>ra, el Vola<strong>de</strong>ro y la Zona <strong>de</strong> la 23, y todas tuvieron
Sánchez, 2004. Morfometría y Demografía <strong>de</strong> E. pycnophylla. 30relaciones altamente significativas (r 2 =0.9749, p
Sánchez, 2004. Morfometría y Demografía <strong>de</strong> E. pycnophylla. 31No se observó, en ninguna <strong>de</strong> las tres localida<strong>de</strong>s, alguna correlación entre porcentaje <strong>de</strong>suelo <strong>de</strong>scubierto y altura <strong>de</strong> las plantas. Sin embargo se observó relación entre elporcentaje <strong>de</strong> suelo <strong>de</strong>scubierto y las alturas mayores en El Vola<strong>de</strong>ro (Tabla 7).4.3.2 Efecto <strong>de</strong> otras especies vegetales4.3.2.1 Relación con altura y diámetro <strong>de</strong> la rosetaEl efecto <strong>de</strong> la altura <strong>de</strong> la paja, Calamagrostis sp., en la altura <strong>de</strong> las rosetas, se observóen las localida<strong>de</strong>s <strong>de</strong>l Páramo <strong>de</strong>l Angel; la Zona <strong>de</strong> la 23 y El Vola<strong>de</strong>ro mostraroncorrelaciones altamente significativas, don<strong>de</strong> la ten<strong>de</strong>ncia es hacia un aumento <strong>de</strong> laaltura <strong>de</strong> los frailejones, al aumentar la altura <strong>de</strong> la paja (Tabla 7 y 8; Anexo 3). En lalocalidad <strong>de</strong> Guan<strong>de</strong>ra no se observó ninguna ten<strong>de</strong>ncia clara; sin embargo, al unir losdatos para la especie, es <strong>de</strong>cir los resultados <strong>de</strong> las tres localida<strong>de</strong>s, se pue<strong>de</strong> ver unaten<strong>de</strong>ncia clara, y significativa (r=0.170, p
Sánchez, 2004. Morfometría y Demografía <strong>de</strong> E. pycnophylla. 32Espeletia (Tabla 7). Con la cantidad <strong>de</strong> Puya sp., no se observó ninguna relaciónsignificativa, aunque la ten<strong>de</strong>ncia es negativa (Fig. 32). En el Vola<strong>de</strong>ro sólo se observóuna relación significativa y positiva, entre la <strong>de</strong>nsidad <strong>de</strong> Puya sp. y el número <strong>de</strong> adultos(Tabla 7). Para el sitio Guan<strong>de</strong>ra se observó que la <strong>de</strong>nsidad <strong>de</strong> puya y paja afectaronsignificativamente la <strong>de</strong>nsidad <strong>de</strong> juveniles (Fig. 31 y 32; Tabla 7).Al unir los datos <strong>de</strong> las tres localida<strong>de</strong>s, se observó correlación entre la paja y la <strong>de</strong>nsidad<strong>de</strong> individuos totales y <strong>de</strong> juveniles (r=-0.531, p
Sánchez, 2004. Morfometría y Demografía <strong>de</strong> E. pycnophylla. 33TABLASTabla 2: Resultados <strong>de</strong> los coeficientes <strong>de</strong> correlación <strong>de</strong> Spearman para los datos <strong>de</strong> Morfometría. * indicalos valores que son significativos con p
Sánchez, 2004. Morfometría y Demografía <strong>de</strong> E. pycnophylla. 34Tabla 4: Resultados <strong>de</strong> los coeficientes <strong>de</strong> correlación <strong>de</strong> Spearman para los datos <strong>de</strong> Estructura <strong>de</strong>Población.* indica los valores que son significativos con p
Sánchez, 2004. Morfometría y Demografía <strong>de</strong> E. pycnophylla. 35Tabla 6: Tabla <strong>de</strong> vida para vida para toda la especie, utilizando los datos obtenidos <strong>de</strong> las tres localida<strong>de</strong>s(N=4616).Número <strong>de</strong> Proporción <strong>de</strong> Mortalidad <strong>de</strong>Clases supervivientes individuos Probabilidad individuos Tasa <strong>de</strong><strong>de</strong> altura al inicio <strong>de</strong> la clase en cada clase <strong>de</strong> supervivencia entre clases mortalidad(x) (Nx) (p) (lx) (dx) (qx)1 2583 0.559 1.0 1422 0.552 1161 0.251 0.449 803 0.693 358 0.077 0.139 156 0.444 202 0.044 0.078 62 0.315 140 0.030 0.054 73 0.526 67 0.014 0.026 22 0.337 45 0.0097 0.017 18 0.48 27 0.0058 0.010 -6 09 33 0.0071 0.013 0 0Tabla 7: Resultados <strong>de</strong> los coeficientes <strong>de</strong> correlación <strong>de</strong> Spearman para los datos <strong>de</strong>l Efecto <strong>de</strong> comunidad.* indica los valores que son significativos con p
Sánchez, 2004. Morfometría y Demografía <strong>de</strong> E. pycnophylla. 36Tabla 8: Resultados <strong>de</strong> la prueba <strong>de</strong> Kruskal Wallis para los datos <strong>de</strong>l Efecto <strong>de</strong> la comunidad. * indica losvalores que son significativos con p
Sánchez, 2004. Morfometría y Demografía <strong>de</strong> E. pycnophylla. 37FIGURAS400Altura <strong>de</strong> la roseta (cm)3002001002425118 147226265291431335 343 317365337 386 324493 372 423387 461 320326418682537664611598 604501519 499 633605 603 516 559 631517649 632 556673 577763683737740724 730718 720 736739735 709 8290N =16215118018919436003700380039004000Altitud (m)FIGURA 5: Relación entre las alturas <strong>de</strong> la roseta y la altitud en la zona <strong>de</strong> la 23.300167301Altura <strong>de</strong> la roseta (cm)200100149150159 108 14172282255254231192239193194 280367336467430510 5305144174250N =1881111121493500360037003800Altitud (m)FIGURA 6: Relación entre las alturas <strong>de</strong> las plantas <strong>de</strong> la localidad Guan<strong>de</strong>ra y la altitud.
Sánchez, 2004. Morfometría y Demografía <strong>de</strong> E. pycnophylla. 38400Altura <strong>de</strong> la roseta (cm)30020010016 3852151133 13856170148 172141167 180186 89282338213286311506509519 4344890N =212136185360037003800Altitud (m)FIGURA 7: Relación entre las alturas <strong>de</strong> las plantas <strong>de</strong> El Vola<strong>de</strong>ro y la altitud.400Altura <strong>de</strong> la roseta (cm)300200112100N =101010101036003700380039004000Altitud (m)FIGURA 8: Variación <strong>de</strong> la diez alturas mayores por altitud al aumentar la elevación, para la Zona <strong>de</strong> la 23
Sánchez, 2004. Morfometría y Demografía <strong>de</strong> E. pycnophylla. 39300Altura <strong>de</strong> la roseta (cm)200100N =101010103500360037003800Altitud (m)FIGURA 9: Variación <strong>de</strong> la diez alturas mayores por altitud al aumentar la elevación, en el sitio <strong>de</strong> estudioGuan<strong>de</strong>ra6005005Altura <strong>de</strong> la roseta (cm)40030027200100N =101010360037003800Altitud (m)FIGURA 10: Variación <strong>de</strong> la diez alturas mayores por altitud al aumentar la elevación, en El Vola<strong>de</strong>ro
Sánchez, 2004. Morfometría y Demografía <strong>de</strong> E. pycnophylla. 40375350Altura máxima (cm)3253002752502252003400 3500 3600 3700 3800 3900 4000 4100AltitudFIGURA 11: Valores <strong>de</strong> altura máxima en cada cota altitudinal, para las localida<strong>de</strong>s don<strong>de</strong> se realizó el estudio., datos correspondientes a la localidad <strong>de</strong> la 23; , datos <strong>de</strong> la localidad, Guan<strong>de</strong>ra y , datos <strong>de</strong> lalocalidad El Vola<strong>de</strong>ro.10080160161188186762Diámetro <strong>de</strong> la roseta (cm)6040207410N =16215118018919436003700380039004000Altitud (m)FIGURA 12: Relación entre el diámetro <strong>de</strong> la roseta y la altitud, en la Zona <strong>de</strong> la 23.
Sánchez, 2004. Morfometría y Demografía <strong>de</strong> E. pycnophylla. 41100357Diámetro <strong>de</strong> la roseta (cm)806040200N =1881111121493500360037003800AltitudFIGURA 13: Relación entre el diámetro <strong>de</strong> la roseta y la altitud, en la localidad Guan<strong>de</strong>ra.100490Diámetro <strong>de</strong> la roseta (cm)806040200N =213136185360037003800Altitud (m)FIGURA 14: Relación entre el diámetro <strong>de</strong> la roseta y la altitud, en El Vola<strong>de</strong>ro.
Sánchez, 2004. Morfometría y Demografía <strong>de</strong> E. pycnophylla. 42100Diámetro <strong>de</strong> la roseta (cm)9080706094050N =101010101036003700380039004000Altitud (m)FIGURA 15: Variación en los diez diámetros mayores por altitud, al aumentar la elevación, en localidad <strong>de</strong> La23.100Diámetro <strong>de</strong> la roseta (cm)90807060752850N =101010103500360037003800Altitud (m)FIGURA 16: Variación en los diez diámetros mayores por altitud, al aumentar la elevación, en la localidadGuan<strong>de</strong>ra.
Sánchez, 2004. Morfometría y Demografía <strong>de</strong> E. pycnophylla. 431002028Diámetro <strong>de</strong> la roseta (cm)90807060N =101010360037003800Altitud (m)FIGURA 17: Cambio en los diez diámetros mayores por altitud, al aumentar la elevación, en El Vola<strong>de</strong>ro.30Número capítulos/inflorescencia2010130191 296 397 179468395 264651 662 652 664 653 663 650 535 654636 606 637 598 635623 666 593 605 665596 661 597 668 537 630 595 560 604 667 624538 533 559 594 634628 694 690 691 629615 626 696 621 692 620 638 639 616 603 576 619 515 693719 720 718723 729 722725 759 758 768 739 772 773831 829 830 823899 825 900 8320N =1663411918915436003700380039004000AltitudFIGURA 18: Variación <strong>de</strong>l número <strong>de</strong> capítulos por inflorescencia en un gradiente altitudinal, para la localidad<strong>de</strong> la 23.
Sánchez, 2004. Morfometría y Demografía <strong>de</strong> E. pycnophylla. 441816Número capítulos/inflorescencia1412108648 3710 11 293174262 49N =82130175360037003800AltitudFIGURA 19: Variación <strong>de</strong>l número <strong>de</strong> capítulos por inflorescencia en un gradiente altitudinal, para la localidad<strong>de</strong>l Vola<strong>de</strong>ro.Densidad <strong>de</strong> adultos/120m 22202102001901801701601501401301201103400 3500 3600 3700 3800 3900 4000 4100Altitud (m)FIGURA 20: Cambio en la <strong>de</strong>nsidad <strong>de</strong> adultos <strong>de</strong> acuerdo a la altitud, en las tres localida<strong>de</strong>s <strong>de</strong> estudio. , datoscorrespondientes a la localidad <strong>de</strong> la 23; , datos <strong>de</strong> la localidad, Guan<strong>de</strong>ra y , datos <strong>de</strong> la localidad ElVola<strong>de</strong>ro.
Sánchez, 2004. Morfometría y Demografía <strong>de</strong> E. pycnophylla. 45120100Densidad juveniles/30m2806040200N =4444436003700380039004000Altitud (m)FIGURA 21: Variación en la <strong>de</strong>nsidad <strong>de</strong> juveniles con respecto a la altitud, en la Zona <strong>de</strong> la 23.300Densidad juveniles/30m22001000-100N =44443500360037003800Altitud (m)FIGURA 22: Variación en la <strong>de</strong>nsidad <strong>de</strong> juveniles con la altitud para la localidad Guan<strong>de</strong>ra.
Sánchez, 2004. Morfometría y Demografía <strong>de</strong> E. pycnophylla. 468070Densidad juveniles/30m26050403020N =444360037003800Altitud (m)FIGURA 23: Variación en la <strong>de</strong>nsidad <strong>de</strong> juveniles con la altitud para El Vola<strong>de</strong>ro.0.7Proporción <strong>de</strong> individuos0.60.50.40.30.20.1Guan<strong>de</strong>raLa 23El Vola<strong>de</strong>ro01 2 3 4 5 6 7 8 9Clases <strong>de</strong> alturaFIGURA 24:Relación entre las clases <strong>de</strong> altura y la proporción <strong>de</strong> individuos, p, en las tres localida<strong>de</strong>s <strong>de</strong> estudio.Las clases <strong>de</strong> altura se encuentran construidas en intervalos regulares <strong>de</strong> 30 cm.
Sánchez, 2004. Morfometría y Demografía <strong>de</strong> E. pycnophylla. 473.532.5Log 1000lx21.510.500 1 2 3 4 5 6 7 8 9 10Clases <strong>de</strong> alturaFIGURA 25: Se muestran las líneas <strong>de</strong> ten<strong>de</strong>ncia para las 3 localida<strong>de</strong>s <strong>de</strong> páramo. , datos correspondientes a lalocalidad <strong>de</strong> la 23 (m=-0.2513, r2=0.9269, p
Sánchez, 2004. Morfometría y Demografía <strong>de</strong> E. pycnophylla. 48Proporción <strong>de</strong> supervivientes(lx)1.210.80.60.40.201 2 3 4 5 6 7 8 9Clases <strong>de</strong> alturaGuan<strong>de</strong>raLa 23El Vola<strong>de</strong>roFIGURA 27: Relación entre las clases <strong>de</strong> altura y la proporción <strong>de</strong> supervivientes (lx), en las tres localida<strong>de</strong>s. Lasclases <strong>de</strong> altura se encuentran construidas en intervalos regulares <strong>de</strong> 30 cm.QxLa 230.80.70.60.50.40.30.20.100 1 2 3 4 5 6 7 8 9 10aClases <strong>de</strong> alturaQxGuan<strong>de</strong>ra0.80.70.60.50.40.30.20.100 1 2 3 4 5 6 7 8 9 10bClases <strong>de</strong> alturaQx0.80.70.60.50.40.30.20.100 1 2 3 4 5 6 7 8 9 10cClases <strong>de</strong> alturaQx0.80.70.60.50.40.30.20.100 1 2 3 4 5 6 7 8 9 10dClases <strong>de</strong> alturaFIGURA 28: Relación entre las clases <strong>de</strong> altura y la tasa <strong>de</strong> mortalidad, qx, en las tres localida<strong>de</strong>s <strong>de</strong> estudio, y anivel <strong>de</strong> la especie. a) relación para la localidad <strong>de</strong> la 23, b) relación para Guan<strong>de</strong>ra, c) Vola<strong>de</strong>ro d) para laespecie. Las clases <strong>de</strong> altura se encuentran construidas en intervalos regulares <strong>de</strong> 30 cm.
Sánchez, 2004. Morfometría y Demografía <strong>de</strong> E. pycnophylla. 49400Altura <strong>de</strong> la roseta (cm)30020010013826831385 6825371395 13106641365724718 7201243 129712661364 611 698 1287 699 7191244 1256 709 12281400 1281 829499 862 633 1302 1372 871 715 603 516 559 1296 1313 501 1293 830 1408 598 787 764 1394 6041227 810605 631 8031363 697 12941254763431737 1710 1214365 17761757 147 287 1470 1141 735 442 1923 1759 1640 1314 739 13301663 1939 1350 1148 271 1348 1517 736 1839 175 965 730 1664 387 461 1559 291 493 24 1558 265 386 1876 740423 372 1745282418255 1919 11951333 12161774 312492726 396 16011496 1804 390 309892 91492811582261027335 343 3171009 101415761089 9321046 337 1162 3240N =4850111113732Altura Calamagrostis sp.FIGURA 29: Relación entre la altura <strong>de</strong> la paja, Calamagrostis sp. y la altura <strong>de</strong> la roseta, en los tres sitios <strong>de</strong>estudio.1001365175 1450 160 1460186 17661216 188Diámetro <strong>de</strong> la roseta (cm)806040201302140013641377 1281 12970N =4850111113732Altura Calamagrostis sp.FIGURA 30: Relación entre la altura <strong>de</strong> Calamagrostis sp. y el diámetro <strong>de</strong> la roseta, en los tres sitios <strong>de</strong>estudio.
Sánchez, 2004. Morfometría y Demografía <strong>de</strong> E. pycnophylla. 50Número juveniles12010080604020Guan<strong>de</strong>raLa 23El Vola<strong>de</strong>ro00 1 2Altura Calamagrostis sp.FIGURA 31: Relación entre la altura <strong>de</strong> la paja, Calamagrostis sp., y el número <strong>de</strong> juveniles en las treslocalida<strong>de</strong>s <strong>de</strong> estudio. 0 se refiere a la altura 1 m.Número juveniles12010080604020Guan<strong>de</strong>raLa 23El Vola<strong>de</strong>ro00 1 2Puya sp.FIGURA 32: Relación entre la cantidad <strong>de</strong> Puya sp., y el número <strong>de</strong> juveniles en las tres localida<strong>de</strong>s. 0 serefiere a la ausencia, 1 a presencia mo<strong>de</strong>rada y 2 a presencia abundante.
Sánchez, 2004. Morfometría y Demografía <strong>de</strong> E. pycnophylla. 515. DISCUSIONPara enten<strong>de</strong>r la evolución y adaptación <strong>de</strong> las plantas a su ambiente, es importanteenten<strong>de</strong>r los efectos <strong>de</strong> la variación <strong>de</strong>l microhábitat en la morfología, fisiología y ladinámica poblacional <strong>de</strong> una especie. El gradiente altitudinal provee una oportunidadúnica para estudiar esto, ya que está asociado con cambios en temperatura, humedad,vegetación y otros caracteres ambientales. Por lo tanto es muy interesante preguntarsecomo cambia una especie a lo largo <strong>de</strong> un gradiente altitudinal.5.1 Morfometría5.1.2 Altura <strong>de</strong> la rosetaSmith (1980) observó una clara ten<strong>de</strong>ncia altitudinal para la altura <strong>de</strong> las rosetas <strong>de</strong> E.schultzii; en ésta, la altura <strong>de</strong> las plantas tien<strong>de</strong> a aumentar conforme aumenta laelevación. Esta hipótesis, según el autor, se cumple para la mayoría <strong>de</strong> especies <strong>de</strong>Espeletia, con sólo algunas excepciones. Smith reporta estas excepciones comoobservaciones, como posibles <strong>de</strong>sviaciones a una ten<strong>de</strong>ncia general. De acuerdo con estasobservaciones, Espeletia pycnophylla <strong>de</strong>bería cumplir un patrón diferente al género, es<strong>de</strong>cir, las rosetas más altas se encontrarían en las altitu<strong>de</strong>s más bajas. Al parecer podría<strong>de</strong>berse al efecto protector <strong>de</strong> la línea <strong>de</strong> bosque.Un estudio realizado en E. pycnophylla (Kovář, 2001), no encontró ninguna relaciónentre la altura <strong>de</strong> la roseta y elevación. Al parecer, según lo propone el autor, el impacto<strong>de</strong>l fuego pue<strong>de</strong> oscurecer la relación. El efecto <strong>de</strong>l disturbio en la altura <strong>de</strong> las plantas,ha sido <strong>de</strong>scrito previamente (Verweij y Kok, 1992; Hofste<strong>de</strong>, 1995; Premauer, 1999), yen general, las plantas más altas se encuentran en los lugares que no han sido sometidos adisturbio antrópico. Sin embargo el estudio realizado por Kovář (2001), se hizo con el fin<strong>de</strong> observar, más exactamente, el efecto <strong>de</strong>l disturbio en esta especie y no <strong>de</strong> estudiar elefecto <strong>de</strong> un gradiente altitudinal. No existen en realidad estudios que prueben lasobservaciones que hizo Smith en las especies que parecen no cumplir su hipótesis.
Sánchez, 2004. Morfometría y Demografía <strong>de</strong> E. pycnophylla. 52En este estudio no se encontró ninguna relación estadísticamente significativa para estecaracter morfológico. En las tres localida<strong>de</strong>s <strong>de</strong> estudio, se encontraron diferentesten<strong>de</strong>ncias, y por lo tanto no se pue<strong>de</strong>n hacer conclusiones para la especie (Fig. 5, 6 y 7).Sólo en la Zona <strong>de</strong> la 23, en el análisis <strong>de</strong> las alturas mayores, se observó una ten<strong>de</strong>nciapositiva, reflejando la hipótesis <strong>de</strong> Smith y contradiciendo su observación para estaespecie (Fig. 8). Sin embargo la correlación es débil, a pesar <strong>de</strong> ser significativa, lo queno permite hacer conjeturas (Tabla 2). En el sitio <strong>de</strong> estudio El Vola<strong>de</strong>ro se encontró unaten<strong>de</strong>ncia negativa significativa, pero se <strong>de</strong>be recordar que en esta localidad, sólo sehicieron mediciones en tres altitu<strong>de</strong>s, por lo que la ten<strong>de</strong>ncia no es tan fiable.En dos <strong>de</strong> las localida<strong>de</strong>s, Guan<strong>de</strong>ra y El Vola<strong>de</strong>ro, se encontró la altura máxima en lasaltitu<strong>de</strong>s más bajas, apoyando las observaciones <strong>de</strong> Smith. Por el contrario, en la zona <strong>de</strong>la 23, este valor estuvo a 4000 m <strong>de</strong> altitud lo que concuerda con los resultados <strong>de</strong> Smith(Fig. 8, 9, 10 y 11). Al unir los datos para las tres altitu<strong>de</strong>s en común –3600, 3700 y 3800m <strong>de</strong> altitud– tampoco se pue<strong>de</strong> observar un patrón claro. Como se pue<strong>de</strong> ver, no existeuna ten<strong>de</strong>ncia inequívoca entre las tres localida<strong>de</strong>s, ni una para la especie, así como loobservó Kovář (2001).Existen varias razones por las que pue<strong>de</strong> no encontrarse el mismo patrón que en E.schultzii. Smith (1980), había <strong>de</strong>scrito que para las especies <strong>de</strong> Espeletia encontradas<strong>de</strong>s<strong>de</strong> el centro-sur <strong>de</strong> Colombia hasta Ecuador no se encontraba el patrón, dado queocurren en sitios muy húmedos cerca <strong>de</strong> la línea <strong>de</strong>l bosque y en sitios muy secos amayores elevaciones. Al estar cercanas al bosque tienen un abastecimiento continuo <strong>de</strong>agua, y por lo tanto pue<strong>de</strong>n crecer más rápido. En la única localidad don<strong>de</strong> se tuvo laoportunidad <strong>de</strong> realizar mediciones cercanas a la línea <strong>de</strong> bosque fue en Guan<strong>de</strong>ra. Esinteresante, que en este caso, se encontró la altura máxima en la elevación más baja (3500m <strong>de</strong> altitud). En las otras dos localida<strong>de</strong>s no existía la línea <strong>de</strong> bosque (frontera <strong>de</strong>colonización y agrícola); sin embargo, en la localidad <strong>de</strong> El Vola<strong>de</strong>ro también seencontró la mayor altura a 3600 m <strong>de</strong> altitud (Fig 11).
Sánchez, 2004. Morfometría y Demografía <strong>de</strong> E. pycnophylla. 53A pesar <strong>de</strong> que las observaciones <strong>de</strong> Smith (1980) predicen que esta especie no aumentala altura <strong>de</strong> las rosetas como respuesta a un gradiente altitudinal, existen otras razones porlas que se pudo ver oscurecida la ten<strong>de</strong>ncia. Las localida<strong>de</strong>s se han encontrado bajo lapresencia <strong>de</strong> disturbio por fuego. Las quemas pue<strong>de</strong>n causar una homogenización en elpáramo, y por lo tanto se pier<strong>de</strong>n las diferencias altitudinales. A<strong>de</strong>más, como ya semencionó anteriormente, el fuego tiene impacto en la altura <strong>de</strong> las plantas y pue<strong>de</strong>retrasar el crecimiento <strong>de</strong> éstas, haciéndolas <strong>de</strong> menor tamaño. Es <strong>de</strong>cir, la altitud ya nosería el único aspecto que estaría afectando la altura <strong>de</strong> las rosetas, sino un conjunto <strong>de</strong>factores que pue<strong>de</strong>n oscurecer la relación.Comparando la metodología utilizada en el presente estudio con la <strong>de</strong> Smith (1980),existen algunas diferencias que pue<strong>de</strong>n hacer que no sean comparables. Smith cubrió unrango altitudinal alre<strong>de</strong>dor <strong>de</strong> 800 metros, entre 3000 y 3800 m <strong>de</strong> altitud. En las treslocalida<strong>de</strong>s utilizadas en este estudio se cubrieron diferentes rangos altitudinales, pero entotal se abarcó un rango <strong>de</strong> 600 metros, entre 3500 y 4000 m <strong>de</strong> altitud. A<strong>de</strong>más <strong>de</strong> ser unrango altitudinal más corto que el estudiado previamente, las altitu<strong>de</strong>s son diferentes ysólo se superponen 300 metros. Pue<strong>de</strong> ser que en altitu<strong>de</strong>s menores, no estudiadas, elpatrón pueda reflejarse más claramente.En Venezuela, se encuentran un sinnúmero <strong>de</strong> especies <strong>de</strong> Espeletia; en Ecuador seencuentra únicamente una especie. Las diferencias en la estructura <strong>de</strong> las comunida<strong>de</strong>s <strong>de</strong>Espeletia pue<strong>de</strong>n tener un efecto <strong>de</strong> competencia intraespecífica en Venezuela, y por lotanto la respuesta frente los factores ambientales pue<strong>de</strong> ser distinta. También existendiferencias a nivel <strong>de</strong> suelos; en Ecuador los suelos son volcánicos, en Venezuela lossuelos tiene una gran influencia glacial.En general, los estudios se realizaron bajo condiciones diferentes, y pue<strong>de</strong> ser que esascondiciones tengan distintos impactos en las rosetas.
Sánchez, 2004. Morfometría y Demografía <strong>de</strong> E. pycnophylla. 545.1.2 Diámetro <strong>de</strong> la rosetaEl crecimiento, medido como altura total, no pareció verse afectado por el gradientealtitudinal. Sin embargo, existen otras medidas <strong>de</strong> crecimiento que pue<strong>de</strong>n reflejar elefecto <strong>de</strong> la elevación. El diámetro <strong>de</strong> la roseta está relacionado con la producción <strong>de</strong>hojas, y por lo tanto con la fotosíntesis. Este carácter pue<strong>de</strong> ser una medida indirecta <strong>de</strong> laproductividad <strong>de</strong> una planta (con respecto a su capacidad fotosintética), o una medida <strong>de</strong>la producción <strong>de</strong> biomasa. Así mismo, el diámetro <strong>de</strong> la roseta pue<strong>de</strong> indicar si haycambios en la disposición <strong>de</strong> las hojas, ya sea para proteger el meristema, o paraaumentar la intercepción <strong>de</strong> rayos solares. El diámetro <strong>de</strong> la roseta está involucrado convarios aspectos importantes para las plantas, por lo que es interesante observar si existeun efecto <strong>de</strong> la elevación en el tamaño <strong>de</strong> la roseta.Un estudio realizado en E. timotensis (Monasterio y Sarmiento, 1991) muestra que lasrosetas tienen hojas más verticales (es <strong>de</strong>cir tienen un menor ángulo <strong>de</strong> apertura), alaumentar el gradiente altitudinal; según se propone, esto ayuda a interceptar máseficientemente los rayos <strong>de</strong>l sol y por lo tanto mejora la asimilación por fotosíntesis. Asímismo, la roseta protege el meristema apical, pues su geometría forma una capa <strong>de</strong>asilamiento térmico.Al utilizar los valores <strong>de</strong> todos los diámetros <strong>de</strong> roseta medidos no se observa, en ninguna<strong>de</strong> las tres localida<strong>de</strong>s, una ten<strong>de</strong>ncia altitudinal clara (Fig. 12, 13 y 14). Tampoco cuandose unieron los datos <strong>de</strong> las tres altitu<strong>de</strong>s comunes. Sin embargo, algunos análisis <strong>de</strong> losdiámetros mayores reflejaron un gradiente altitudinal en las localida<strong>de</strong>s <strong>de</strong>l páramo <strong>de</strong>lAngel (El Vola<strong>de</strong>ro y la Zona <strong>de</strong> la 23) (Tabla 2; Fig. 15 y 17). Aunque no en todos estosanálisis se observa una ten<strong>de</strong>ncia significativa, en varios <strong>de</strong> ellos se ve, que al aumentarla elevación el tamaño máximo <strong>de</strong>l diámetro disminuye.La altitud pue<strong>de</strong> imponer un límite en el tamaño <strong>de</strong>l diámetro <strong>de</strong> las rosetas. Aunque laten<strong>de</strong>ncia para todas las plantas medidas (el análisis <strong>de</strong> todos los diámetros) no reflejanun patrón, los valores extremos pue<strong>de</strong>n dar información acerca <strong>de</strong>l efecto <strong>de</strong> la altitud. Unmenor diámetro, pue<strong>de</strong> indicar menor tamaño <strong>de</strong> las hojas y un ángulo <strong>de</strong> apertura <strong>de</strong>
Sánchez, 2004. Morfometría y Demografía <strong>de</strong> E. pycnophylla. 55éstas más pequeño. Un cambio <strong>de</strong> tamaño reflejaría la ten<strong>de</strong>ncia que generalmente ocurreen los bosques: al aumentar la elevación; las hojas son un caracter que tien<strong>de</strong> a versemodificado, cambiando su tamaño <strong>de</strong> mesófilas a micrófilas, y a mayores elevaciones ananófilas (Giles, 1975; Whitmore, 1991). Esto se <strong>de</strong>be a que al aumentar la elevación, lasradiaciones UV son mayores, y las temperaturas –entre otras condiciones–, son másextremas (Vargas 1998a; Luteyn, 1999). Con respecto a un cambio en el ángulo, podríaocurrir lo mismo que se observa en E. timotensis, don<strong>de</strong> las rosetas acomodarían sushojas para maximizar la fotosíntesis y proteger el meristema, <strong>de</strong>bido a la presión por lascondiciones ambientales.En la localidad <strong>de</strong> Guan<strong>de</strong>ra no se observó ninguna ten<strong>de</strong>ncia clara, lo que pue<strong>de</strong> <strong>de</strong>bersea factores propios <strong>de</strong>l lugar. En el caso <strong>de</strong>l Páramo <strong>de</strong>l Angel; al parecer, la altitud estásiendo un parámetro que impone un límite en el crecimiento <strong>de</strong> las hojas y en el grado <strong>de</strong>apertura <strong>de</strong> éstas. Pue<strong>de</strong>n existir otras variables que estén afectando el comportamiento<strong>de</strong> este caracter, pero se pue<strong>de</strong> ver una ten<strong>de</strong>ncia.5.1.3 Capítulos por inflorescenciaEl gradiente altitudinal pue<strong>de</strong> tener un efecto no sólo en el crecimiento <strong>de</strong> las plantas,sino también en su reproducción. Las condiciones climáticas pue<strong>de</strong>n tener un impacto enla producción <strong>de</strong> inflorescencias, capítulos y flores. El número <strong>de</strong> capítulos porinflorescencia pue<strong>de</strong> dar una i<strong>de</strong>a <strong>de</strong> cómo la elevación afecta alguno <strong>de</strong> los aspectosreproductivos <strong>de</strong> las rosetas.Un estudio previo realizado en E. pycnophylla (Kovář, 2001), muestra que el número <strong>de</strong>capítulos por inflorescencia se ve afectado por la altitud. Este estudio, sin embargo,presenta problemas en la metodología. El objetivo <strong>de</strong>l estudio era <strong>de</strong>terminar el efecto <strong>de</strong>ldisturbio en un gradiente altitudinal; para esto, es necesario comparar lugares disturbadoscon lugares sin disturbio y tener las mismas elevaciones. En el muestreo se tomaron tresaltitu<strong>de</strong>s sometidas a fuego (3600, 3850 y 4100 m <strong>de</strong> altitud) y una sola sin presencia <strong>de</strong>disturbio (4100 m <strong>de</strong> altitud). Las comparaciones con el sitio sin disturbio no son muy
Sánchez, 2004. Morfometría y Demografía <strong>de</strong> E. pycnophylla. 56válidas, ya que no se tiene un gradiente altitudinal, y a<strong>de</strong>más sólo hay un punto <strong>de</strong>comparación.En las medidas <strong>de</strong>l número <strong>de</strong> capítulos por inflorescencia, el autor llega a la conclusiónque este valor es mayor en los sitios con quemas que en los que no hay disturbio. Sintener en cuenta estas diferencias entre sitios con disturbio y sin disturbio, los valorestien<strong>de</strong>n a disminuir al aumentar la elevación (aunque a 4100 m <strong>de</strong> altitud bajo presencia<strong>de</strong> disturbio los valores tien<strong>de</strong>n a incrementarse). Los valores, sin embargo, no son muycomparables con los <strong>de</strong>l presente estudio, <strong>de</strong>bido a la diferencia en la división <strong>de</strong> clases(el autor subdivi<strong>de</strong> los individuos mayores a 30 cm en dos categorías: intermedios yadultos).En este estudio es muy clara la ten<strong>de</strong>ncia con el gradiente altitudinal. En las doslocalida<strong>de</strong>s don<strong>de</strong> se estudió, se vio una disminución en el número <strong>de</strong> capítulos alaumentar el gradiente altitudinal (Tabla 2; Fig. 18 y 19). Esta ten<strong>de</strong>ncia se pue<strong>de</strong> <strong>de</strong>ber alas características propias <strong>de</strong>l ecosistema. Así mismo, las plantas tien<strong>de</strong>n a utilizar susrecursos <strong>de</strong> manera que optimicen el crecimiento y la reproducción (Perrin y Civil, 1993;Ashman, 1994). A mayores elevaciones los recursos tien<strong>de</strong>n a ser más escasos y pue<strong>de</strong>nestarse utilizando más en el mantenimiento <strong>de</strong> la planta y su crecimiento, que en lareproducción. Al aumentar la altitud el <strong>de</strong>sarrollo tien<strong>de</strong> a ser más lento <strong>de</strong>bido a que laproductividad disminuye, la <strong>de</strong>scomposición es más lenta y la absorción <strong>de</strong> nutrientes escomplicada (Castaño, 1998; Luteyn, 1999). Este patrón pue<strong>de</strong> reflejar la plasticidad queposee esta especie bajo diferentes presiones ambientales. Otra razón por la que pue<strong>de</strong>encontrarse esta ten<strong>de</strong>ncia sería <strong>de</strong>bido a diferencias genéticas; sin embargo pensar enuna diferenciación genética <strong>de</strong>bido a un gradiente altitudinal resulta complicado ya quelas poblaciones no se encuentran muy distanciadas entre sí, y muy posiblemente existeflujo genético. A<strong>de</strong>más pue<strong>de</strong> ser que el número <strong>de</strong> inflorescencias por planta aumente oque el número <strong>de</strong> flores por capítulo sea mayor, para compensar la disminución en elnúmero <strong>de</strong> capítulos. En realidad no se pue<strong>de</strong> <strong>de</strong>cir que el potencial reproductivo <strong>de</strong> laespecie disminuya con la altitud sin antes ver otras características florales. Es másprobable pensar que las diferencias se <strong>de</strong>ben a la plasticidad fenotípica <strong>de</strong> la especie.
Sánchez, 2004. Morfometría y Demografía <strong>de</strong> E. pycnophylla. 57El cambio en el número <strong>de</strong> capítulos por inflorescencia indica una variación altitudinal enla arquitectura floral. Esto pue<strong>de</strong> tener implicaciones taxonómicas ya que los caracteresflorales tien<strong>de</strong>n a ser muy utilizados en la <strong>de</strong>scripción <strong>de</strong> las especies <strong>de</strong> Espeletia. Esnecesario, por lo tanto, conocer la variación en los caracteres florales al hacer<strong>de</strong>scripciones.En un estudio realizado para la misma especie, en Nariño – Colombia (Armero, com.pers.), se encontró que a una altitud <strong>de</strong> 3360 + 15 m, el número <strong>de</strong> capítulos porinflorescencia era <strong>de</strong> 7 + 2.7. Es interesante que este valor es menor que el obtenido para3600 m <strong>de</strong> altitud en Ecuador. Las diferencias encontradas se pue<strong>de</strong>n <strong>de</strong>ber a que son doslugares <strong>de</strong> estudio distintos, al efecto <strong>de</strong>l disturbio sobre las plantas o se pue<strong>de</strong> ser pordiferencias genéticas entre las poblaciones. Para probar el efecto <strong>de</strong> la elevación que seobservó en el presente estudio, habría que repetir en la localidad <strong>de</strong> Nariño un muestreoen gradiente altitudinal para ver si se cumple el patrón. Pue<strong>de</strong> ser que igualmentedisminuya el número <strong>de</strong> capítulos por inflorescencia con el gradiente, o que aumenten yen algún punto vuelvan a disminuir.5.2 Estructura Poblacional5.2.1 Densidad <strong>de</strong> individuosEstudios realizados en E. schultzii y E. pycnophylla (Smith, 1980; Kovář, 2001),muestran que el número <strong>de</strong> juveniles disminuye con la altitud <strong>de</strong>bido a factoresambientales y <strong>de</strong> competencia. Así mismo, existen otros factores que pue<strong>de</strong>n influir enestas <strong>de</strong>nsida<strong>de</strong>s. La <strong>de</strong>nsidad <strong>de</strong> individuos, tanto adultos como juveniles, es sensible aldisturbio y al estado <strong>de</strong> sucesión <strong>de</strong>l páramo. Aunque en los sitios sin disturbio antrópicose muestran las <strong>de</strong>nsida<strong>de</strong>s <strong>de</strong> juveniles más gran<strong>de</strong>s, y según lo reportado, el disturbiofuego tien<strong>de</strong> a disminuir estas <strong>de</strong>nsida<strong>de</strong>s (Verweij y Kok, 1992; Hofste<strong>de</strong>, 1995), lospáramos maduros poseen un número bajo <strong>de</strong> plántulas, haciendo que las distribuciones <strong>de</strong>clases sean más uniformes (Llambí et al., en prensa).
Sánchez, 2004. Morfometría y Demografía <strong>de</strong> E. pycnophylla. 58Con respecto al disturbio por fuego, al parecer su impacto es diferente en adultos yjuveniles. Los adultos tien<strong>de</strong>n a ser los más afectados, aumentando la mortalidad(Verweij y Kok, 1992; Laegaard, 1992), mientras que los juveniles se recuperan pronto.Los juveniles mueren inmediatamente luego <strong>de</strong> la quema, pero esta clase se reemplazarápidamente pues se estimula la germinación y se benefician <strong>de</strong>l espacio libre, ladisponibilidad <strong>de</strong> luz y nutrientes (Verweij y Kok, 1992).En este estudio no se tuvo en cuenta el efecto exacto <strong>de</strong>l disturbio como para relacionaresta variable <strong>de</strong> manera precisa con los patrones encontrados. Sin embargo, mediante lasobservaciones y anotaciones <strong>de</strong> campo se pudo <strong>de</strong>terminar que todos los transectos <strong>de</strong> lasdiferentes localida<strong>de</strong>s estuvieron sujetos a la presencia <strong>de</strong>l fuego. También se tuvo unai<strong>de</strong>a general <strong>de</strong> que tan reciente había sido el disturbio, por la presencia <strong>de</strong> cenizas en lostallos <strong>de</strong> las rosetas. El disturbio por pastoreo pudo estar presente en las localida<strong>de</strong>s,aunque no se observó presencia <strong>de</strong> estiércol.En los tres sitios <strong>de</strong> estudio se observa un comportamiento diferente <strong>de</strong> las <strong>de</strong>nsida<strong>de</strong>s <strong>de</strong>acuerdo a la altitud. Esto se pue<strong>de</strong> <strong>de</strong>ber a las condiciones ambientales propias <strong>de</strong> cadalugar y al efecto que pue<strong>de</strong> y pudo tener el disturbio en cada uno. A pesar <strong>de</strong> que en E.pycnophylla se reportó esta ten<strong>de</strong>ncia, en el presente estudio no se encuentra ningúnpatrón claro que permita hacer una generalización para la especie (Tabla 4; Fig. 21, 22 y23), ya que ninguno <strong>de</strong> los análisis para juveniles fueron estadísticamente significativos(Tabla 4). Al ser las <strong>de</strong>nsida<strong>de</strong>s <strong>de</strong> individuos tan sensibles al efecto <strong>de</strong>l disturbio, asícomo aspectos ambientales, pue<strong>de</strong> ser que una misma especie muestre comportamientodiferente según la presión a la que se encuentra sujeto.En el caso <strong>de</strong> la <strong>de</strong>nsidad <strong>de</strong> adultos, tampoco se observa una ten<strong>de</strong>ncia clara, ya que laslocalida<strong>de</strong>s se comportaron <strong>de</strong> diferente manera (Tabla 4; Fig. 20). Sólo en la Zona <strong>de</strong> la23 se pudo ver una relación clara entre la <strong>de</strong>nsidad y la altitud; en ésta, el número <strong>de</strong>adultos por área aumenta con la elevación. Esta ten<strong>de</strong>ncia no está <strong>de</strong> acuerdo con lo quese espera teóricamente. Sin embargo si el impacto <strong>de</strong>l disturbio es mayor en las altitu<strong>de</strong>s
Sánchez, 2004. Morfometría y Demografía <strong>de</strong> E. pycnophylla. 59menores, por la cercanía a las zonas pobladas, se podría esperar que el mayor número <strong>de</strong>rosetas se encuentren a mayor elevación, don<strong>de</strong> el impacto no sería tan fuerte.Al ver en <strong>de</strong>talle el patrón que se observa en la Figura 20 y 23 para la localidad <strong>de</strong> ElVola<strong>de</strong>ro, se observan, en las tres altitu<strong>de</strong>s, las mismas ten<strong>de</strong>ncias. Las mayores<strong>de</strong>nsida<strong>de</strong>s <strong>de</strong> juveniles y <strong>de</strong> adultos se encuentran en la menor altitud (i.e 3600 m <strong>de</strong>altitud), lo que estaría <strong>de</strong> acuerdo con la teoría. En 3700 m <strong>de</strong> altitud hay una disminucióndrástica en las <strong>de</strong>nsida<strong>de</strong>s tanto <strong>de</strong> juveniles como <strong>de</strong> adultos. Según las observaciones <strong>de</strong>campo, en este lugar se notó que el impacto <strong>de</strong>l disturbio había sido muy fuerte, lo quepodría explicar la disminución en la <strong>de</strong>nsidad <strong>de</strong> adultos.5.2.2 Tablas <strong>de</strong> VidaLa mayoría <strong>de</strong> estudios en <strong>de</strong>mografía <strong>de</strong> Espeletia se han realizado en una misma altitudy observando el efecto <strong>de</strong>l disturbio. Así como en morfología, la altitud pue<strong>de</strong> tener unefecto sobre la distribución <strong>de</strong> clases <strong>de</strong> eda<strong>de</strong>s <strong>de</strong> una población. Pue<strong>de</strong> existir un efecto<strong>de</strong>l gradiente <strong>de</strong> elevación en la <strong>de</strong>mografía <strong>de</strong> la población, llevando a que factorescomo la tasa <strong>de</strong> supervivencia y mortalidad se vean afectados por las condicionesclimáticas, que tien<strong>de</strong>n a ser más extremas al aumentar la altitud. Es interesante, por lotanto ver el comportamiento <strong>de</strong> una población <strong>de</strong> Espeletia bajo un gradiente altitudinal.Según los resultados <strong>de</strong> Verweij y Kok (1992), la curva a la cual se ajustaron la estructura<strong>de</strong> eda<strong>de</strong>s <strong>de</strong> E. hartwegiana en un sitio sin disturbio, era logarítmica. Este mismo patrónfue encontrado para E. barclayana (Cavelier et al,.1992). Esta curva es bastante comúnpara plantas, y lo que indica es un alto número juveniles que va <strong>de</strong>creciendologarítmicamente hasta tener unos pocos adultos, <strong>de</strong> mayores alturas.Las curvas logarítmicas suelen encontrarse en sitios sin disturbio don<strong>de</strong> se consi<strong>de</strong>ra quela población está estable o incluso en crecimiento. El disturbio tiene efecto en ladistribución <strong>de</strong> las clases haciendo que algunas disminuyan drásticamente y que incluso,algunas <strong>de</strong>saparezcan (Verweij y Kok, 1992; Premauer, 1999). Las tasas <strong>de</strong> mortalidad
Sánchez, 2004. Morfometría y Demografía <strong>de</strong> E. pycnophylla. 60también se ven afectadas siendo más fuertes que en los sitios sin disturbio, y aumentandocon el tamaño <strong>de</strong> la roseta (Verweij y Kok, 1992). Al no haber disturbio, esta tasa sueleser constante a lo largo <strong>de</strong> todas las clases <strong>de</strong> altura (Verweij y Kok, 1992), o es mayor anivel <strong>de</strong> juveniles y plántulas (Estrada y Monasterio, 1991; Hofste<strong>de</strong>, 1995; Premauer,1999).En este estudio se observó una relación entre las clases <strong>de</strong> altura y el número <strong>de</strong>individuos (nx), la proporción <strong>de</strong> individuos (p) y la proporción <strong>de</strong> supervivientes (lx)(Fig. 24 y 27). Estos resultados son consistentes en las tres localida<strong>de</strong>s e indican que estasvariables van disminuyendo conforme aumenta la altura <strong>de</strong> las rosetas. Los tres sitios secomportaron <strong>de</strong> manera muy similar al comparar la proporción <strong>de</strong> individuos; este valorpermanece muy regular en cada clase. A pesar <strong>de</strong> que el número <strong>de</strong> individuos varíe, elaporte <strong>de</strong> cada clase a la siguiente se mantiene proporcionalmente igual así se encuentrenen diferentes lugares. La especie como tal, posee un patrón <strong>de</strong>mográfico bastante regular.Con respecto a las curvas <strong>de</strong> supervivencia se encontró, en todos los casos, –es <strong>de</strong>cir en elanálisis por transecto, por cota altitudinal, por localidad y para la especie–, curvaslogarítmicas (Fig. 25 y 26). A pesar <strong>de</strong> existir presencia <strong>de</strong> disturbio en los tres sitios <strong>de</strong>estudio, las poblaciones se comportaron como las poblaciones sin disturbio (Fig. 33). Seencontraron algunas diferencias a nivel <strong>de</strong> tasa <strong>de</strong> supervivencia en transectos <strong>de</strong> lamisma altitud y diferencias entre cotas altitudinales, sin embargo las ten<strong>de</strong>ncias son muyconsistentes y en ninguna <strong>de</strong> las localida<strong>de</strong>s se encontró un gradiente altitudinal en laestructura <strong>de</strong>mográfica <strong>de</strong> la población. Aunque las poblaciones se encuentren bajodiferentes condiciones, ya sean locales, <strong>de</strong> microhábitat o con diferente historia <strong>de</strong>disturbio, se observa el mismo comportamiento <strong>de</strong>mográfico. Pue<strong>de</strong> ser que, a pesar <strong>de</strong>que la especie se encuentra sujeta a la presencia <strong>de</strong> quemas, se recupere rápidamente o nose vea tan afectada por él ya que no se pier<strong>de</strong>n clases <strong>de</strong> altura como se encuentrareportado para E. killipii (Premauer, 1999). (Fig. 33)Las tasas <strong>de</strong> mortalidad para cada una <strong>de</strong> las localida<strong>de</strong>s, aunque no son constantes,tampoco aumentan con la clase <strong>de</strong> altura como se supone ocurre en los sitios con
Sánchez, 2004. Morfometría y Demografía <strong>de</strong> E. pycnophylla. 61disturbio (Fig. 28). Se observa que la tasa varía con la clase <strong>de</strong> altura, es <strong>de</strong>cir no hay unaclase que se caracterice por tener una mayor mortalidad. Al parecer la tasa <strong>de</strong> mortalidadse encuentra sujeta a las condiciones <strong>de</strong> cada lugar, ya que se comporta <strong>de</strong> diferentemanera en las localida<strong>de</strong>s. También es importante resaltar que las caídas tan abruptas sepue<strong>de</strong>n <strong>de</strong>ber a que las clases <strong>de</strong> alturas más gran<strong>de</strong>s tienen pocos individuos lo que lashace más sensibles a cambios bruscos, como en el caso <strong>de</strong> El Vola<strong>de</strong>ro. La especie tieneuna tasa <strong>de</strong> mortalidad, también variable, aunque se encuentra más concentrada en las dosprimeras clases <strong>de</strong> altura (Fig. 24). Este patrón , <strong>de</strong> mayor mortalidad en las primerasclases, es lo que se suele encontrar en los sitios sin disturbio antrópico. Nuevamente, estaespecie se estaría comportando como las especies en ausencia <strong>de</strong> disturbio.FIGURA 31: Tomada <strong>de</strong> Premauer (1999). Mo<strong>de</strong>lo <strong>de</strong>l posible comportamiento <strong>de</strong> la estructura <strong>de</strong> eda<strong>de</strong>s <strong>de</strong>la población <strong>de</strong> Espeletia killipii sometida a disturbios continuos. (a): Curva logarítmica <strong>de</strong> una poblaciónsin disturbio. (b): Efecto <strong>de</strong> disturbio continuo, don<strong>de</strong> se ven afectadas las primeras clases. (c, d, e): Efecto<strong>de</strong>l disturbio continuo; se empiezan a per<strong>de</strong>r clases. (f): Población extinta.Estos resultados, sin embargo, sólo muestran el estado <strong>de</strong> las poblaciones en un momentodado, es <strong>de</strong>cir sirven para tener una i<strong>de</strong>a general en el momento <strong>de</strong> hacer el estudio. Paratener una mejor i<strong>de</strong>a <strong>de</strong> las tasas <strong>de</strong> mortalidad y la dinámica poblacional, el estudio <strong>de</strong>be
Sánchez, 2004. Morfometría y Demografía <strong>de</strong> E. pycnophylla. 62exten<strong>de</strong>rse por más tiempo. Como lo reporta Allphin y Harper (1997), para una especie <strong>de</strong>la familia Asteraceae, nunca hay estabilidad en la distribución <strong>de</strong> las clases <strong>de</strong> altura.5.3 Efecto <strong>de</strong> comunidadExisten otros factores que pue<strong>de</strong>n influir en los patrones <strong>de</strong> morfometría y en la estructurapoblacional, a<strong>de</strong>más <strong>de</strong> la altitud. Entre estos se encuentran otras característicastopográficas (pendiente, exposición, etc), efecto <strong>de</strong>l disturbio, competencia con otrasespecies vegetales, entre otras. Dado que el efecto <strong>de</strong>l gradiente altitudinal no resultó sermuy claro en las medidas <strong>de</strong> crecimiento (altura y diámetro <strong>de</strong> la roseta), se utilizaron lasmedidas <strong>de</strong> cobertura vegetal y la presencia <strong>de</strong> otras especies dominantes, para ver elcomportamiento <strong>de</strong> la población <strong>de</strong> Espeletia frente a éstas.5.3.1 Cobertura vegetalSegún lo reportado por Smith (1980), la cobertura vegetal disminuye al aumentar laelevación. En este caso las localida<strong>de</strong>s <strong>de</strong> Guan<strong>de</strong>ra y El Vola<strong>de</strong>ro, mostraron la mismaten<strong>de</strong>ncia que reporta el autor. En la Zona <strong>de</strong> la 23 no se encontró <strong>de</strong> manera tan clara, loque se pue<strong>de</strong> <strong>de</strong>ber a factores locales, como las condiciones <strong>de</strong> microclima y la historia <strong>de</strong>disturbio en cada una <strong>de</strong> las cotas altitudinales. En esta localidad es interesante que elporcentaje <strong>de</strong> cobertura vegetal es más o menos el mismo en todas las altitu<strong>de</strong>s a excepción<strong>de</strong> 3800 m <strong>de</strong> altitud.El porcentaje <strong>de</strong> suelo <strong>de</strong>scubierto se correlacionó linearmente con la altura <strong>de</strong> las rosetas<strong>de</strong> E. schultzii (Smith, 1980). Esta conclusión es un poco esperada, ya que si la elevación serelaciona con la altura <strong>de</strong> las plantas, y la cobertura <strong>de</strong>l suelo lo hace con la altitud, pues laaltura <strong>de</strong> las plantas <strong>de</strong>bería relacionarse con el porcentaje <strong>de</strong> cobertura.En las tres localida<strong>de</strong>s <strong>de</strong> estudio, no se cumplió la relación entre altura <strong>de</strong> las plantas y elporcentaje <strong>de</strong> suelo <strong>de</strong>scubierto. Aunque en el caso <strong>de</strong> Guan<strong>de</strong>ra y El Vola<strong>de</strong>ro la relaciónentre altitud y cobertura fue negativa, así como la relación entre altitud y altura <strong>de</strong> la roseta,no hay ninguna relación entre cobertura y altura. Esto se <strong>de</strong>be a que las relaciones con laaltura total <strong>de</strong> las plantas no son claras. Al utilizar los valores mayores y el máximo <strong>de</strong>
Sánchez, 2004. Morfometría y Demografía <strong>de</strong> E. pycnophylla. 63altura <strong>de</strong> roseta, se observan algunas ten<strong>de</strong>ncias en El Vola<strong>de</strong>ro. En esta localidad, lasmayores alturas <strong>de</strong> la roseta no sólo estarían limitadas por la altitud sino también por elporcentaje <strong>de</strong> suelo <strong>de</strong>scubierto, lo que podría indicar la presencia <strong>de</strong> un conjunto <strong>de</strong>factores que limitan el crecimiento <strong>de</strong> las rosetas.5.3.2 Efecto <strong>de</strong> otras especies vegetalesLos datos <strong>de</strong> la altura y el diámetro <strong>de</strong> la roseta, como ya se mencionó anteriormente, notuvieron ninguna relación con el gradiente <strong>de</strong> elevación. Sin embargo se pue<strong>de</strong> pensar enque existen otros factores que están afectando la distribuciones <strong>de</strong> las alturas o los tamaños<strong>de</strong> los diámetros. La paja, Calamagrostis sp., junto con los frailejones, tienen dominio <strong>de</strong>lpaisaje florístico. Al ser tan dominante, pue<strong>de</strong> tener un efecto en las poblaciones <strong>de</strong>Espeletia pycnophylla tanto en el sentido <strong>de</strong>mográfico, como morfológico.Con respecto a los datos <strong>de</strong> morfometría se observó, que tanto la altura como el diámetro <strong>de</strong>la roseta, se ven afectados por la altura <strong>de</strong>l pajonal. En especial se ve una ten<strong>de</strong>ncia clara enlas localida<strong>de</strong>s <strong>de</strong>l Páramo <strong>de</strong>l Angel, es <strong>de</strong>cir en la Zona <strong>de</strong> la 23 y en El Vola<strong>de</strong>ro.Aunque en Guan<strong>de</strong>ra no se observó ningún patrón relacionado con la altura <strong>de</strong> la paja, alutilizar los datos <strong>de</strong> todas las alturas se pudo comprobar que sí existe una misma ten<strong>de</strong>nciapara la especie (Fig. 29 y 30): cuando la paja es más alta, los frailejones tien<strong>de</strong>n a tenermayor tamaño.Los datos <strong>de</strong> todas las alturas y diámetros <strong>de</strong> la roseta no están respondiendo a un gradientealtitudinal, pero podrían hacerlo a diferentes factores <strong>de</strong>l medio. Un mayor tamaño en lapaja y <strong>de</strong> la población <strong>de</strong> Espeletia podría indicar una misma historia <strong>de</strong> disturbio en unlugar dado, o se pue<strong>de</strong> <strong>de</strong>ber a características edafológicas. También se pue<strong>de</strong> pensar en unefecto <strong>de</strong> competencia, en especial por luz. Al ser la paja más alta, los individuos máspequeños <strong>de</strong> Espeletia podrían ser eliminados por la falta <strong>de</strong> luz, y así sólo se encuentranlos más altos.
Sánchez, 2004. Morfometría y Demografía <strong>de</strong> E. pycnophylla. 64Con respecto a la <strong>de</strong>nsidad, un estudio realizado para E. pycnophylla en el Volcán Chiles(Kovář, 2001), muestra que al haber una menor cobertura <strong>de</strong> paja, el número <strong>de</strong> juvenilestien<strong>de</strong> a ser mayor. Hofste<strong>de</strong> (1995), también reporta que al removerse las cobertura vegetalpor el fuego, el número <strong>de</strong> juveniles <strong>de</strong> E. hartwegiana es más alto. Esto podría explicarsebajo el efecto <strong>de</strong> competencia interespecífica, ya que las otras especies podrían competircon los juveniles <strong>de</strong> Espeletia por el espacio, luz y disponibilidad <strong>de</strong> nutrientes. Alaumentar la cobertura <strong>de</strong> estas especies, el espacio para nuevos juveniles se ve limitado.En las localida<strong>de</strong>s <strong>de</strong> estudio, se observaron diferentes impactos <strong>de</strong> otras especies vegetales,como Calamagrostis sp. y Puya sp. En Guan<strong>de</strong>ra fue claro que las dos especies serelacionan con la disminución <strong>de</strong> la <strong>de</strong>nsidad <strong>de</strong> juveniles; en La 23, aparentemente la pajaes la única que tiene una correlación significativa. Por el contrario, en El Vola<strong>de</strong>ro ninguna<strong>de</strong> las dos especies afectan la <strong>de</strong>nsidad <strong>de</strong> juveniles (Tabla 4; Fig. 30 y 31).Con respecto a la <strong>de</strong>nsidad <strong>de</strong> adultos, ninguno <strong>de</strong> los análisis relacionados a la altura <strong>de</strong> lapaja fueron significativos. La <strong>de</strong>nsidad <strong>de</strong> puya, por el contrario, mostró una correlaciónpositiva pero sólo en la localidad <strong>de</strong> El Vola<strong>de</strong>ro. Al parecer las <strong>de</strong>nsida<strong>de</strong>s <strong>de</strong> adultos nose encuentran muy afectadas por la presencia <strong>de</strong> otras especies (Tabla 4).El efecto <strong>de</strong> la elevación en la disminución <strong>de</strong> la altura <strong>de</strong> la paja se observa en las treslocalida<strong>de</strong>s, aunque Kovář (2001), reporta lo contrario. Se esperaría que si la altura <strong>de</strong> lapaja disminuye con la altitud y que aparentemente, la disminución en tamaño <strong>de</strong> la pajacorrespon<strong>de</strong> a una disminución en la altura y el diámetro <strong>de</strong> la roseta, y a un aumento en la<strong>de</strong>nsidad <strong>de</strong> juveniles, estas variables <strong>de</strong>berían verse afectadas a su vez por la altitud.Se esperaría entonces que la altura y el diámetro <strong>de</strong> la roseta disminuyera con la elevación yel número <strong>de</strong> juveniles aumentara con la altitud.Al parecer los frailejones no se ven afectados por la altitud <strong>de</strong> la misma manera que otrasespecies vegetales. Aunque pue<strong>de</strong> existir un factor <strong>de</strong> competencia interespecífica, pue<strong>de</strong>nexistir otros factores que sean más <strong>de</strong>terminantes en la estructura morfométrica y<strong>de</strong>mográfica. Con respecto a la <strong>de</strong>nsidad <strong>de</strong> juveniles, Smith (1980) reporta que para E.
Sánchez, 2004. Morfometría y Demografía <strong>de</strong> E. pycnophylla. 65schultzii, el número <strong>de</strong> juveniles disminuye con la altitud <strong>de</strong>bido al estrés ambiental y por elefecto <strong>de</strong> competencia. En este caso se podría <strong>de</strong>cir que la <strong>de</strong>nsidad <strong>de</strong> juveniles se pudover afectada por el efecto <strong>de</strong> otras especies, especialmente la paja, pero no se presentarelación con la altitud.
Sánchez, 2004. Morfometría y Demografía <strong>de</strong> E. pycnophylla. 666. CONCLUSIONES- No se encontró un efecto <strong>de</strong>l gradiente altitudinal en la altura <strong>de</strong> las rosetas. En cadauna <strong>de</strong> las localida<strong>de</strong>s se observó una ten<strong>de</strong>ncia diferente, lo que no permite haceruna generalización para E. pycnophylla. Esto está <strong>de</strong> acuerdo con lo reportadopreviamente para la especie, pero no corrobora la hipótesis planteada para lamayoría <strong>de</strong> las especies <strong>de</strong> Espeletia.- El diámetro <strong>de</strong> la roseta es otra medida <strong>de</strong> crecimiento en estas plantas, por lo quepue<strong>de</strong> verse afectado por la altitud. Se encontró, en las localida<strong>de</strong>s <strong>de</strong>l Páramo <strong>de</strong>lAngel, un efecto <strong>de</strong> la altitud en el tamaño máximo <strong>de</strong>l diámetro <strong>de</strong> la roseta. Laaltitud pue<strong>de</strong> estar limitando el tamaño que este caracter pue<strong>de</strong> alcanzar.- Existe un efecto altitudinal en el número <strong>de</strong> capítulos por inflorescencia: alaumentar la elevación el número <strong>de</strong> capítulos disminuye. Esto pue<strong>de</strong> indicar unadisminución en el potencial reproductivo <strong>de</strong> la especie, aunque <strong>de</strong>be corroborarsecon datos <strong>de</strong> número <strong>de</strong> inflorescencias y flores por capítulo. A su vez, pue<strong>de</strong>indicar plasticidad fenotípica <strong>de</strong> la especie y tener implicaciones taxonómicas.- El efecto <strong>de</strong>l gradiente altitudinal fue claro en sólo uno <strong>de</strong> los caracteresmorfológicos –capítulos por inflorescencia–, mientras que en los otros dos –altura ydiámetro <strong>de</strong> la roseta– no hay una ten<strong>de</strong>ncia clara. La variación morfológica pue<strong>de</strong>verse afectada por otras condiciones como el disturbio y la competencia.- Teóricamente se espera una disminución en la <strong>de</strong>nsidad <strong>de</strong> juveniles y adultos alaumentar la elevación. Sin embargo, las <strong>de</strong>nsida<strong>de</strong>s no respondieron <strong>de</strong> manerainequívoca en las tres localida<strong>de</strong>s. No se pudo <strong>de</strong>terminar un efecto <strong>de</strong> la altitud enestas <strong>de</strong>nsida<strong>de</strong>s.- Las Tablas <strong>de</strong> Vida reflejaron consistencia en cada una <strong>de</strong> las localida<strong>de</strong>sestudiadas. La especie tuvo un mismo comportamiento <strong>de</strong>mográfico sin importar las
Sánchez, 2004. Morfometría y Demografía <strong>de</strong> E. pycnophylla. 67condiciones <strong>de</strong> cada lugar. Las proporciones <strong>de</strong> individuos en las clase <strong>de</strong> alturafueron muy similares en cada sitio, lo que muestra una constancia en las clases.Todas las curvas <strong>de</strong> supervivencia fueron logarítimicas, y la tasa <strong>de</strong> mortalidad esvariable en cada una <strong>de</strong> las localida<strong>de</strong>s y para la especie aunque la mayormortalidad se encuentra concentrada en las primeras clases <strong>de</strong> altura.- El porcentaje <strong>de</strong> cobertura vegetal disminuyó con la elevación, en las localida<strong>de</strong>s <strong>de</strong>Guan<strong>de</strong>ra y El Vola<strong>de</strong>ro. No se encontró relación entre la altura <strong>de</strong> las rosetas y elporcentaje <strong>de</strong> cobertura vegetal. Al parecer no se pue<strong>de</strong> pensar en un efectocombinado <strong>de</strong> altitud y porcentaje <strong>de</strong> cobertura vegetal en la altura <strong>de</strong> las rosetas.- La altura <strong>de</strong> la paja, Calamagrostis sp., y la altura y diámetro <strong>de</strong> las rosetasmostraron una correlación positiva. Aunque los dos caracteres <strong>de</strong> E. pycnophylla nomostraron una relación concreta con la altitud, se observa que sí existe con la altura<strong>de</strong>l pajonal. Esto podría indicar una misma historia <strong>de</strong> disturbio o se pue<strong>de</strong> <strong>de</strong>ber acaracterísticas edafológicas. También se pue<strong>de</strong> pensar en un efecto <strong>de</strong> competencia.- El efecto <strong>de</strong> otras especies vegetales, como Calamagrostis sp. y Puya sp., fuediferente en cada localidad. En la Zona <strong>de</strong> la 23 la paja mostró correlación con la<strong>de</strong>nsidad <strong>de</strong> juveniles; mientras que en Guan<strong>de</strong>ra, la paja y puya se correlacionaroncon la <strong>de</strong>nsidad <strong>de</strong> juveniles. En el Vola<strong>de</strong>ro ninguna <strong>de</strong> las especies afectó la<strong>de</strong>nsidad <strong>de</strong> juveniles pero sí la <strong>de</strong> adultos.- Existe un efecto <strong>de</strong>l gradiente altitudinal en la altura <strong>de</strong> la paja. A medida queaumenta la elevación, la paja va reduciendo su altura. Aunque se observa relaciónentre la altura paja y la altitud, y entre la altura paja y la altura y diámetro <strong>de</strong> laroseta, los caracteres <strong>de</strong> los frailejones no varían con altitud. Existen por lo tanto,otras razones, como disturbio o condiciones locales, que están afectando estasmedidas.
Sánchez, 2004. Morfometría y Demografía <strong>de</strong> E. pycnophylla. 687. RECOMENDACIONES- Realizar este estudio en otras especies <strong>de</strong> Espeletia que no cumplan con la ten<strong>de</strong>nciareportada por Smith (1980). Estas especies son las que se encuentran distribuidas<strong>de</strong>s<strong>de</strong> el Centro <strong>de</strong> Colombia hasta Ecuador.- Realizar un estudio comparativo entre varias especies encontradas en una mismalocalidad, relacionándolas con su hábito y su hábitat.- Estudiar la variación altitudinal en otras rosetas gigantes, para comprobar si laforma <strong>de</strong> crecimiento tiene algún efecto sobre el comportamiento frente a laelevación.- Observar la variación altitudinal <strong>de</strong> caracteres reproductivos: número <strong>de</strong>inflorescencias por planta, <strong>de</strong> capítulos por inflorescencia, <strong>de</strong> flores por capítulos yproducción <strong>de</strong> semillas, para ver si existe una disminución <strong>de</strong>l potencialreproductivo con la elevación.- Si es posible, cubrir un rango altitudinal más amplio al realizar el estudio y realizarel estudio teniendo en cuenta el disturbio.- Hacer un estudio <strong>de</strong>mográfico por más tiempo en E. pycnophylla para ver si enrealidad la especie tiene un comportamiento tan estable. Así mismo realizar en otrasespecies <strong>de</strong>l género estudios <strong>de</strong>mográficos en un gradiente altitudinal.
Sánchez, 2004. Morfometría y Demografía <strong>de</strong> E. pycnophylla. 69LITERATURA CITADAAllphin L. y K. T. Harper. 1997. Demography and Life History Characteristics of the RareKachina Daisy (Erigeron kachinensis, Asteraceae). The American MidlandNaturalist. 138: 109-120.Ashman, T. 1994. A dynamic perspective on the physiological cost of reproduction inplants. The American Naturalist. 144: 300-316.Barbour, M. G., J. H. Burk y W. D. Pitts. 1980. Terrestrial Plant Ecology. TheBenjamin/Commings Publishing Company, Inc. USA. Pp. 59-70.Baruch, Z. y A. P Smith. 1979. Morphological and Physiological Correlates of NicheBreadth in Two Species of Espeletia (Compositae) in the Venezuelan An<strong>de</strong>s.Oecologia. 38: 71-82Beard, J. S. 1955. The Classification of Tropical American Vegetation-Types. Ecology. 36:89-100.Begon, M., J. L. Harper y C. R. Townsend. 1996. Ecology: individuals, populations andcommunities. Tercera edición. Blackwell Science Publishers. Cambridge. Pp. 147-162.Berry, P. E. y R. N. Calvo. 1989. Wind pollination, self-incompatibility, and altitudinalshifts in pollination systems in the high An<strong>de</strong>an genus Espeletia (Asteraceae).American Journal of Botany. 76: 1602-1614.Berry, P. E. y R. N. Calvo. 1994. An overview of the reproductive biology of Espeletia(Asteraceae) in the Venezuelan An<strong>de</strong>s., pp. 229-249 in Tropical AlpineEnvironments, edited by P. W. Run<strong>de</strong>l, A. P. Smith and F. C. Meinzer. CambridgeUniversity Press, Great Britain.
Sánchez, 2004. Morfometría y Demografía <strong>de</strong> E. pycnophylla. 70Byers D. L. y T. R. Meagher. 1997. A Comparison of Demographic Characteristics in arare and a common Species of Eupatorium. Ecological Applications. 7: 519-530.Carlquist, S. 1974. Island Biology. Columbia University Press, New York.Castaño, C. 1995. El hombre y el continum <strong>de</strong>l páramo, pp. 17-36 en El Páramo:Ecosistema a proteger, editado por Reyes, P. ECOAN, Bogotá.Caughley, G. 1966. Mortality patterns in Mammals. Ecology. 47: 906-918Cavelier, J., J. L. Machado, D. Valencia, J. Montoya, A. Laignelet, A. Hurtado, A. Varela,y C. Mejia. 1992. Leaf Demography and Growth Rates of Espeletia barclayanaCuatrec. (compositae), a Caulescent Rossete in a Colombian Paramo. Biotropica.24: 52-63.Comuna <strong>de</strong> La Libertad. 2001. Plan <strong>de</strong> manejo comunitario <strong>de</strong> los páramos <strong>de</strong> La Libertad.Documento no publicado. QuitoCondit, R., R. Sukumar, S. P. Hubbell, y R. B. Foster. 1998. Predicting Population Trendsfrom Size Distributions: A Direct Test in a Tropical Tree Community. TheAmerican Naturalist. 152: 495-509.Crisp M. D. y R. T. Lange. 1976. Age structure, distribution and survival un<strong>de</strong>r grazing ofthe arid-zone shrub Acacia burkittii. Oikos. 27: 86-92.Cuatrecasas, J. 1979. Growth forms of the Espeletiinae and their correlation to vegetationtypes of the high tropical An<strong>de</strong>s, pp. 397-410, in Tropical Botany, edited by Larsen,K. and L. B Holm-Nielsen. Aca<strong>de</strong>mic Press, London.
Sánchez, 2004. Morfometría y Demografía <strong>de</strong> E. pycnophylla. 71Cuatrecasas, J., 1986. Speciation and radiation of the Espeletiinae in the An<strong>de</strong>s, pp. 267-303 in High Altitu<strong>de</strong> Tropical Biogeography, edited by F. Vuilleumier and M.Monasterio. Oxford University Press, New York.Emerson, B. C. 2002. Evolution on oceanic islands: molecular phylogenetic approaches toun<strong>de</strong>rstanding pattern and process. Molecular ecology. 11: 951-966.Estrada, C. y M. Monasterio. 1988. Ecología poblacional <strong>de</strong> una roseta gigante, Espeletiaspicata Sch. Bip. (Compositae), <strong>de</strong>l páramo <strong>de</strong>sértico. Ecotropicos. 1: 25-39.Estrada, C. y M. Monasterio. 1991. Comportamiento reproductivo <strong>de</strong> una roseta gigante,Espeletia spicata Sch. Bip. (Compositae), <strong>de</strong>l páramo <strong>de</strong>sértico. Ecotropicos. 4: 1-17.Fundación para el <strong>de</strong>sarrollo agropecuario, “fundagro”. 1998. Un análisis global <strong>de</strong> lasituación actual y potencial para el <strong>de</strong>sarrollo <strong>de</strong> la cuenca <strong>de</strong>l río El Angel, enInforme final <strong>de</strong> activida<strong>de</strong>s Proyecto Manejo <strong>de</strong> Recursos, Carchi, Ecuador.Giles, E. Jr. 1975. Structure and climate in tropical rain forest. Annual Review of Ecologyand Systematics. 6:.67-86Grubb, P. J., J. R. Lloyd, T. D Pennington, T. C. Whitmore. 1963. A Comparison ofMontane and Lowland Rain Forest in Ecuador I. The Forest Structure,Physiognomy and Floristics. Journal of Ecology. 51: 567-601.González F. L. G. Y F. Cár<strong>de</strong>nas. 1995. El páramo un paisaje <strong>de</strong>shumanizado: el caso <strong>de</strong>las provincias <strong>de</strong>l Norte y Gutierrez (Boyacá – Colombia), pp 63-80 en El páramo:Ecosistema <strong>de</strong> alta montaña, editado por Reyes, P. ECOAN, Bogotá.Guariguata, M. R. y A. Azocar. 1988. Seed Bank Dynamics and Germination Ecology inEspeletia timotensis (Compositae), an An<strong>de</strong>an Giant Rosette. Biotropica. 20: 54-59.
Sánchez, 2004. Morfometría y Demografía <strong>de</strong> E. pycnophylla. 72Hallé, F. R., A. A. Ol<strong>de</strong>man and P. B. Tomlinson. 1978. Tropical Trees and Forests: AnArchitectural Analysis. Springer-Verlag Berlin. Germany.Hedberg, I. y O. Hedberg. 1979. Tropical-alpine life-forms of vascular plants. Oikos. 33:297-307.Hobbs, R. J. y L. F. Huenneke. 1992. Disturbance, Diversity, and Invasion: Implications forConservation. Conservation Biology. 6: 324-337.Hofste<strong>de</strong>, R. 1995. Effects of burning and grazing on a Colombian paramo ecosystem. Ph.D. Dissertation, University of Amsterdam.Hofste<strong>de</strong>, R. 2001. El impacto <strong>de</strong> las activida<strong>de</strong>s humanas sobre el páramo, pp. 161-185, enLos páramos <strong>de</strong> Ecuador. Particularida<strong>de</strong>s, Problemas y Perspectivas, editado porMena, P., G. Medina y R. Hofste<strong>de</strong>. Abya Yala/Proyecto Páramo. Quito.Janzen, D. H. 1973. Rate of Regeneration after a Tropical High Elevation Fire. Biotropica.5: 117-122.Jules E. S. 1998. Habitat Fragmentation and Demographic Change for a Common Plant:Trillium in Old-Growth Forest. Ecology. 79: 1645-1656.Knox, E. B. y J. D. Palmer. 1995. Chloroplast DNA variation and the recent radiation of thegiant senecios (Asteraceae) on the tall mountains of eastern Africa. Proc. Natl.Acad. Sci. USA. 92: 10349-10353.Kovář, P. 2001. Effects of burning on Espeletia pycnophylla stands in the páramo ofVolcán Chiles, Ecuador –a short-term case study, pp. 113-118, en The Ecology ofVolcán Chiles: high-altitu<strong>de</strong> ecosystem on the Ecuador-Colombian bor<strong>de</strong>r, editadopor P.M. Ramsay. Plymouth: Pebble & Shell. UK.
Sánchez, 2004. Morfometría y Demografía <strong>de</strong> E. pycnophylla. 73Krebs, C. J. 1989. Ecological methodology. Harper Collins Publishers. New York.Krohne, D. T. 1998. General Ecology. Wadsworth Publishing Company. USA.Laegaard, S. 1992. Páramos: why study them? pp. 151-170 en Páramo: An An<strong>de</strong>anEcosystem un<strong>de</strong>r Human Influence, editado por Balslev, H. And J. Luteyn.Aca<strong>de</strong>mic Press. London.Lawton, R. O. 1982. Wind stress and elfin stature in a montane rain forest tree: an adaptiveexplanation. American Journal of Bototany. 69: 1224-1230.Llambí, L. D., R. Law y A. Hodge. En prensa. Temporal changes in local spatial structureof late – successional species: establishment of an an<strong>de</strong>an caulescent rosette plant.Journal of Ecology.Lowe V. P. W. 1969. Population dynamics of the red <strong>de</strong>er (Cervus elaphus L.) on Rhum.Journal of Animal Ecology. 38: 425-457Luteyn, J. L. 1999. Páramos: a checklist of plant diversity, geographical distribution, andbotanical literature. New York Botanical Gar<strong>de</strong>n Press, Bronx, N.YMena Vásconez, P. y G. Medina. 2001. La biodiversidad en los Páramos en el Ecuador, pp.27-52, en Los páramos <strong>de</strong> Ecuador. Particularida<strong>de</strong>s, Problemas y Perspectivas,editado por Mena, P., G. Medina y R. Hofste<strong>de</strong>. Abya Yala/Proyecto Páramo.Quito.Monasterio, M. 1986. Adaptive strategies of Espeletia in the An<strong>de</strong>an <strong>de</strong>sert páramo, pp.49-80, in High Altitu<strong>de</strong> Tropical Biogeography, edited by F. Vuilleumier and M.Monasterio. Oxford University Press, New York.
Sánchez, 2004. Morfometría y Demografía <strong>de</strong> E. pycnophylla. 74Monasterio, M. y L. Sarmiento. 1991. Adaptive radiation of Espeletia in the cold An<strong>de</strong>antropics. Trends in Ecology & Evolution 6: 387-391.Neill, D. 1999. Vegetation, in Catalogue of the Vascular Plants of Ecuador, edited by P. MJørgensen and S. León-Yanez. Missouri Botanical Gar<strong>de</strong>n Press, St. Louis.Ochoa, K. K. 1994. Banco <strong>de</strong> semillas y aspectos ecofisiológicos <strong>de</strong> la germinación <strong>de</strong>Espeletia grandiflora H. B. K. en el páramo El Granizo. Trabajo <strong>de</strong> Grado, Facultad<strong>de</strong> Ciencias, Carrera Biología, Pontificia Universidad Javeriana. Bogotá.Perrin N. y R. M. Civil. 1993. Dynamic mo<strong>de</strong>ls of energy allocation and investment.Annual Review of Ecology and Systematics. 24: 379-410.Premauer, J. M. 1999. Efecto <strong>de</strong> diferentes regimenes <strong>de</strong> disturbio por quema y pastoreosobre la estructura horizontal y vertical <strong>de</strong> la vegetacion <strong>de</strong> paramo, ParqueNacional Natural Chingaza. Trabajo <strong>de</strong> Grado, Facultad <strong>de</strong> Ciencias, CarreraBiología, Universidad Nacional <strong>de</strong> Colombia. BogotáRauscher, J. T. 2000. Molecular systematics of the Espeletia complex: Evi<strong>de</strong>nce fromNRITS sequence on the evolution of an an<strong>de</strong>an adaptive radiation.. Ph.D. thesis,Washington University, Saint Louis, Missouri.Reyes, P. (ed.). 1998. El páramo: ecosistema a proteger. ECOAN. Bogotá.Reyes, P. (ed.). 1998. El páramo: ecosistema <strong>de</strong> alta montaña. ECOAN. Bogotá.Richards, P. W. 1996. The tropical rain forest at its altitudinal and latitudinal limits., pp.419-454, in The Tropical Rain Forest: an ecological study. Edited by Cambridge,University Press, Cambridge.Rudas C. M. 1998. Evaluación <strong>de</strong>l Estado Actual <strong>de</strong> una Población <strong>de</strong> la Palma <strong>de</strong> Cera,Ceroxylon sasaimae Galeano: Aportes a su Historia <strong>de</strong> Vida y Estudio
Sánchez, 2004. Morfometría y Demografía <strong>de</strong> E. pycnophylla. 75Demográfico. Trabajo <strong>de</strong> Grado, Facultad <strong>de</strong> Ciencias, Departamento <strong>de</strong> Biología,Universidad <strong>de</strong> los An<strong>de</strong>s. Bogotá.Run<strong>de</strong>l, P. W., A. P. Smith y F. C. Meinzer (eds.). 1994. Tropical alpine environments:Plant form and function. Cambridge University Press, Cambridge.Sarmiento, F. O. y L. M. Frolich. 2002. An<strong>de</strong>an Cloud Forest Tree Lines: Naturalness,Agriculture and the Human Dimension. Mountain Research and Development. 22:278-287.Simpson, B. B. y C. A. Todzia. 1990. Patterns and Processes in the <strong>de</strong>velopment of the highAn<strong>de</strong>an flora. American Journal of Botany. 77: 1419-1432.Smith, A. C. y M. F. Koch. 1935. The genus Espeletia: a study in phylogenetic taxonomy.Brittonia. 1: 479-532.Smith, A. P. 1980. The Paradox of plant height in An<strong>de</strong>an giant rosette species. Journal ofEcology. 68: 63-73.Smith A. P. 1981. Growth and Population Dynamics of Espeletia (Compositae) in theVenezuelan An<strong>de</strong>s. Smithsonian Contributions to Botany. 48. SmithsonianInstitution Press.Smith, A. P. y T. P. Young. 1987. Tropical alpine plant ecology. Annual Review ofEcology and Systematics. 18: 137-158.Sobrevila, C. 1989. Effects of pollen donors on seed formation in Espeletia schultzii(Compositae) populations at different altitu<strong>de</strong>s. Plant Systematics and Evolution.166: 45-67.
Sánchez, 2004. Morfometría y Demografía <strong>de</strong> E. pycnophylla. 76Squeo, F. A., F. Rada, A. Azocar, y G. Goldstein. 1991. Freezing tolerance and avoidancein high tropical An<strong>de</strong>an plants: Is it equally represented in species with differentplant height? Oecologia. 86: 378-382.Standley, P. C. 1915. The genus Espeletia. American Journal of Botany. 2: 468-486.Stiling, P. 1999. Ecology: theories and applications. Tercera Edición. Prentice Hall. USA.Pp. 150-161.Torrenegra, R. D. y A. N. Tellez. 1999. Química y actividad biológica <strong>de</strong> especies <strong>de</strong>lgénero Espeletia, Informe final. Proyecto <strong>de</strong> Investigación, Departamento <strong>de</strong>Química, Facultad <strong>de</strong> Ciencias, Pontificia Universidad Javeriana, Bogotá.Van <strong>de</strong>r Hammen, T., y A. M. Cleef. 1986. Development of the high An<strong>de</strong>an paramo floraand vegetation, pp. 153-201, in High Altitu<strong>de</strong> Tropical Biogeography, edited by F.Vuilleumier and M. Monasterio. Oxford University Press, New York.Vargas, O. 1998a. Impacto <strong>de</strong>l Fuego y Pastoreo sobre el Medio Ambiente Páramo, pp. 63-72, en El páramo: Ecosistema a proteger, editado por Reyes, P. ECOAN, Bogotá.Vargas, O. 1998b. Problemas para la conservación, protección y manejo <strong>de</strong> los páramos,pp. 29-48, en Caracterización y Manejo <strong>de</strong> las zonas <strong>de</strong> Páramo: memorias <strong>de</strong>lcurso, editado por el Ministerio <strong>de</strong>l Medio Ambiente – ICFES. Bogotá.Verweij, P. A. y K. Kok. 1992. Effects of fire and grazing on Espeletia hartwegianapopulations, pp. 215-229, en Páramo: An an<strong>de</strong>an ecosystem un<strong>de</strong>r humaninfluence, edited by Balslev H. and L. Luteyn. Aca<strong>de</strong>mic Press, London.Verweij, P. A. y K. Kok. 1995. Effects of fire and grazing on plant populations, pp. 97-126,en Spatial and temporal mo<strong>de</strong>lling of vegetation patterns. Burning and grazing in
Sánchez, 2004. Morfometría y Demografía <strong>de</strong> E. pycnophylla. 77the paramo of Los Nevados National Park, Colombia. Ph. D. Dissertation,University of Amsterdam.Verweij, P. A. 1995. Spatial and temporal mo<strong>de</strong>lling of vegetation patterns. Burning andgrazing in the paramo of Los Nevados National Park, Colombia. Ph. D.Dissertation, University of Amsterdam.Vuilleumier, F. 1970. Insular Biogeography in continental regions. I. The northern An<strong>de</strong>sof South America. The American Naturalist. 104: 373-388.Whitmore, T. C. 1991. What are tropical rain forests?, pp. 15-20, in An introduction toTropical Rain Forests, edited by T. C Whitmore. Oxford University Press, NewYork.Wilson, C., J. Grace, S. Allen y F. Slack. 1987. Temperature and stature: a study oftemperature in montane vegetation. Functional Ecology. 1: 405-413.Zar, J. H. 1999. Biostatistical Análisis. Cuarta Edición. Prentice Hall. USA.Zebrowski C. y M. Sourdat. 1997. Los factores <strong>de</strong> la pedogénesis y los suelos en Ecuador, enGeografía básica <strong>de</strong>l Ecuador, Tomo IV: Geografía Física, editado por Huttel C., C.Zebrowski y P. Gondard. Quito.Zebrowski C. y M. Sourdat. 1999. Paisajes agrarios <strong>de</strong>l Ecuador, en Geografía básica <strong>de</strong>l Ecuador,Tomo V: Geografía Agraria, editado por Huttel C., C. Zebrowski y P. Gondard. Quito.
Sánchez, 2004. Morfometría y Demografía <strong>de</strong> E. pycnophylla. 78ANEXOSANEXO 1. Descripción <strong>de</strong> los transectos.Zona la 23Rango altitudinal: 3600-4000 m <strong>de</strong> altitudTransecto Altitud + 5 Ubicación GPS-DMS Pendiente % Cobertura vegetalm <strong>de</strong> altitud Latitud (N) Longitud (W)3600 1 3600 0°42'44.9" 79°29'20.7" 38 702 3605 0°42'47.9" 79°29'20.7" 41 673 3605 0°42'51.5" 79°29'20.7" 40 614 3605 0°42'56.2" 79°29'20.7" 40 673700 1 3700 0°43'1.8" 79°29'20.7" 38 582 3705 0°42'59.5" 79°29'20.7" 41 673 3695 0°42'52.4" 79°29'20.7" 39 544 3700 0°42'49" 79°29'20.7" 39 703800 1 3800 0°43'37.5" 79°29'20.8" 40 942 3795 0°43'32.4" 79°29'20.8" 38 973 3795 0°43’50.2” 79°29'20.8" 40 974 3800 0°43'52.8" 79°29'20.8" 40 913900 1 3900 0°43'41.6" 79°29'20.8" 36 582 3900 0°43'48" 79°29'20.8" 42 733 3900 0°43'48.2" 79°29'20.8" 42 704 3895 0°43'47.4" 79°29'20.8" 40 644000 1 4000 0°44'0.7" 79°29'20.8" 40 612 3995 0°44'3.4" 79°29'20.8" 40 793 3995 0°44'21.3" 79°29'20.8" 39 54.54 3995 0°44'19.89" 79°29'20.8" 41 67El Vola<strong>de</strong>ro Rango altitudinal: 3600-3800 m <strong>de</strong> altitud3600 1 3600 0°41'3" 79°29'20.6" 36 942 3600 0°41'5.2" 79°29'20.6" 38 973 3595 0°41'12.7" 79°29'20.6" 38 914 3600 0°41'18.89" 79°29'20.6" 36 943700 1 3700 0°42'37" 79°29'20.7" 40 732 3705 0°42'34.1" 79°29'20.7" 42 763 3700 0°42'32.1" 79°29'20.7" 40 764 3700 0°42'26.8" 79°29'20.7" 42 733800 1 3800 0°42'21.6" 79°29'20.7" 40 642 3800 0°42'21.8" 79°29'20.7" 39 583 3800 0°42'26.9" 79°29'20.7" 38 85
Sánchez, 2004. Morfometría y Demografía <strong>de</strong> E. pycnophylla. 794 3800 0°42'26.1" 79°29'20.7" 40 54Guan<strong>de</strong>raRango altitudinal: 3500-3800 m <strong>de</strong> altitudTransecto Altitud + 5 Ubicación GPS-DMS Pendiente % Cobertura vegetalm <strong>de</strong> altitud Latitud (N) Longitud (W)3500 1 3495 0°36'18.6" 79°29'20.39" 39 972 3505 0°36'23.9" 79°29'20.39" 36 883 3505 0°36'32.4" 79°29'20.39" 36 914 3505 0°36'33" 79°29'20.39" 36 943600 1 3605 0°36'1.7" 79°29'20.39" 40 942 3605 0°36'6" 79°29'20.39" 39 943 3605 0°36'0.1" 79°29'20.39" 38 974 3605 0°35'54.9" 79°29'20.3" 42 943700 1 3700 0°35'55.9" 79°29'20.39" 40 882 3705 0°35'59.7" 79°29'20.39" 39 853 3700 0°36'1" 79°29'20.39" 40 894 3705 0°36'4.09" 79°29'20.39" 36 853800 1 3800 0°36'16.3" 79°29'20.39" 40 702 3800 0°36'13.6" 79°29'20.39" 38 763 3795 0°36'18.8" 79°29'20.39" 38 674 3795 0°36'24.7" 79°29'20.39" 40 70
Sánchez, 2004. Morfometría y Demografía <strong>de</strong> E. pycnophylla. 80ANEXO 2. Vegetación encontrada en cada uno <strong>de</strong> los transectos.Zona <strong>de</strong> la 23Altitud Transecto Tipo <strong>de</strong> vegetación3600 1En general la <strong>de</strong>nsidad <strong>de</strong> vegetación es poca: Calamagrostis sp. con altura
Sánchez, 2004. Morfometría y Demografía <strong>de</strong> E. pycnophylla. 81Guan<strong>de</strong>raAltitudTransecto Tipo <strong>de</strong> vegetación3500 1Alta <strong>de</strong>nsidad <strong>de</strong> vegetación: Calamagrostis sp. con altura >1m, muchas Puya sp., Corta<strong>de</strong>ria sp., arbustos <strong>de</strong>Asteraceae e Hypericum sp., y helechos2 Calamagrostis sp. con altura
Sánchez, 2004. Morfometría y Demografía <strong>de</strong> E. pycnophylla. 82ANEXO 3. Figuras Complementarias400Altura <strong>de</strong> la roseta (cm)300200100289 311325424406 41132924 443414 256440 453459 362229421 445432 228404 403 290 205 449 430 25 318 484166118 147 436312818549638 874749 704614 673 847588 8221002 1371906 914 8881374936908 957 895 11201064 1384 994 1299 943958 1032 1354 891897 1083989 13371063 1013 1217 1322 1255 1319 1163 1232 1183 13391270 11671303128612450N =485399514360037003800Altitu<strong>de</strong>s (m)FIGURA 1: Relación entre la altura <strong>de</strong> la roseta y la elevación, en las tres altitu<strong>de</strong>s comunes <strong>de</strong> las localida<strong>de</strong>s.100297510 831765 5121175Diámetro <strong>de</strong> la roseta (cm)806040200N =486399514360037003800Altitud (m)FIGURA 2: Relación entre el diámetro <strong>de</strong> la roseta y la elevación, en las tres altitu<strong>de</strong>s comunes <strong>de</strong> laslocalida<strong>de</strong>s.
Sánchez, 2004. Morfometría y Demografía <strong>de</strong> E. pycnophylla. 8310Núm prom.Capítulos/inflorescencia9876543500 3600 3700 3800 3900 4000 4100Altitud (m)FIGURA 3: Efecto <strong>de</strong> la altitud en el promedio <strong>de</strong>l número <strong>de</strong> capítulos por inflorescencia para la localidad <strong>de</strong> la23 (◊), El Vola<strong>de</strong>ro (□) y para la especie (∆).30Capítulos por inflorescencia201013011594 16 152 11395 13 72 121403 508 391 609255 275 680 286476 288 254 607 315101510169711013 940 934 1018 1047970 1033 1056 1019 1058 941 943 1038 1057874 1003 1041 864 875 942 866 876 865 747 1054 993 994 1014 863 992 862 1017 10550N =248471366360037003800Altitud (m)FIGURA 4: Relación entre el número <strong>de</strong> capítulos por inflorescencia y la elevación, en las tres altitu<strong>de</strong>s comunes<strong>de</strong> las localida<strong>de</strong>s.
Sánchez, 2004. Morfometría y Demografía <strong>de</strong> E. pycnophylla. 84100Porcentaje cobertura/30m29080706050N =4444436003700380039004000Altitud (m)FIGURA 5: Variación en el porcentaje <strong>de</strong> cobertura vegetal <strong>de</strong> acuerdo a la altitud, para la localidad <strong>de</strong> La 23.100Porcentaje cobertura/30m290807060N =44443500360037003800Altitud (m)FIGURA 6: Cambio en el porcentaje <strong>de</strong> cobertura vegetal con la altitud, en el sitio <strong>de</strong> estudio Guan<strong>de</strong>ra.
Sánchez, 2004. Morfometría y Demografía <strong>de</strong> E. pycnophylla. 85100Porcentaje cobertura/30m29080706050N =444360037003800Altitud (m)FIGURA 7: Variación en el porcentaje <strong>de</strong> cobertura vegetal <strong>de</strong> acuerdo a la altitud, para el sitio <strong>de</strong> estudio ElVola<strong>de</strong>ro.400Altura <strong>de</strong> la roseta (cm)300200100683682537664724718 720829 709719 699 611 698604810 501 830 598 787 764697 605 631 803708 517 766 800788 556 649 632 875 850 767 809 519 805 499 862 633 871 715 603 516 559763431737365740 386 265291 493 372 423 24730 387 461271 418736 282 175255739287 396 492 442312735726226335 343 3170N =34445973012Altura Calamagrostis sp.FIGURA 8: Relación entre la altura <strong>de</strong> la paja, Calamagrostis sp. y la altura <strong>de</strong> la roseta, en la Zona <strong>de</strong> la 23. 0 serefiere a la altura 1 m.
Sánchez, 2004. Morfometría y Demografía <strong>de</strong> E. pycnophylla. 86300301167Altura <strong>de</strong> la roseta (cm)200100367467149336150255430108510 254 530514 350 23161 192348 87 395 365417 134282159 140N =4659512Altura Calamagrostis sp.FIGURA 9: Relación entre la altura <strong>de</strong> la paja, Calamagrostis sp. y la altura <strong>de</strong> la roseta, en Guan<strong>de</strong>ra. 1 a laaltura entre 50 cm y 1m y 2 a la altura >1 m.400Altura <strong>de</strong> la roseta (cm)300200100506509519 43448936739042133889457 340 454474 272 472319438265447 25516 3852282151133 138213 560N =141187205012Altura Calamagrostis sp.FIGURA 10: Relación entre la altura <strong>de</strong> la paja y la altura <strong>de</strong> la roseta, en la localidad El Vola<strong>de</strong>ro. 0 se refiere ala altura <strong>de</strong> 1 m.
Sánchez, 2004. Morfometría y Demografía <strong>de</strong> E. pycnophylla. 87100188186160175Diámetro <strong>de</strong> la roseta (cm)806040200N =34445973012Altura Calamagrostis sp.FIGURA 11: Relación entre la altura <strong>de</strong> la paja y el diámetro <strong>de</strong> la roseta, en la Zona <strong>de</strong> la 23. 0 se refiere a laaltura 1 m.10035741 51Diámetro <strong>de</strong> la roseta (cm)806040200N =4651952Altura Calamagrostis sp.FIGURA 12: Relación entre la altura <strong>de</strong> la paja y el diámetro <strong>de</strong> la roseta, en la localidad <strong>de</strong> Guan<strong>de</strong>ra. 0 serefiere a la altura 1 m.
Sánchez, 2004. Morfometría y Demografía <strong>de</strong> E. pycnophylla. 88100489340Diámetro <strong>de</strong> la roseta (cm)806040200N =141187205012Altura Calamagrostis sp.FIGURA 13: Relación entre la altura <strong>de</strong> la paja y el diámetro <strong>de</strong> la roseta, en El Vola<strong>de</strong>ro. 0 se refiere a laaltura 1 m.120100Número total juveniles80604017200N =102710012Altura Calamagrostis sp.FIGURA 14: Relación entre la altura <strong>de</strong> la paja, Calamagrostis sp., y el número total <strong>de</strong> juveniles. 0 se refierea la altura 1 m.
Sánchez, 2004. Morfometría y Demografía <strong>de</strong> E. pycnophylla. 89ANEXO 4. Tablas <strong>de</strong> vida por altitud para cada una <strong>de</strong> las localida<strong>de</strong>sLocalidad: La 23Tabla 1: Tabla <strong>de</strong> vida para Espeletia pycnophylla a 3600 m <strong>de</strong> altitud (N=383)Número <strong>de</strong> Proporción <strong>de</strong> Mortalidad <strong>de</strong>Clases supervivientes Individuos Probabilidad individuos Tasa <strong>de</strong><strong>de</strong> altura al inicio <strong>de</strong> la clase en cada clase <strong>de</strong> supervivencia entre clases mortalidad(x) (Nx) (p) (lx) (dx) (qx)1 221 0.577 1.000 119 0.5382 102 0.266 0.462 72 0.7063 30 0.078 0.136 13 0.4334 17 0.044 0.077 8 0.4715 9 0.023 0.041 7 0.7786 2 0.005 0.009 1 0.5007 1 0.003 0.005 0 08 1 0.003 0.005 1 09 0 0 0 0 0Tabla 2: Tabla <strong>de</strong> vida para E. pycnophylla a 3700 m <strong>de</strong> altitud (N=373)Número <strong>de</strong> Proporción <strong>de</strong> Mortalidad <strong>de</strong>Clases supervivientes individuos Probabilidad individuos Tasa <strong>de</strong><strong>de</strong> altura al inicio <strong>de</strong> la clase en cada clase <strong>de</strong> supervivencia entre clases mortalidad(x) (Nx) (p) (lx) (dx) (qx)1 222 0.595 1.000 157 0.7072 65 0.174 0.293 32 0.4923 33 0.088 0.149 11 0.3334 22 0.059 0.099 5 0.2275 17 0.046 0.077 10 0.5886 7 0.019 0.032 3 0.4297 4 0.011 0.018 2 0.5008 2 0.005 0.009 1 0.5009 1 0.003 0.005 1 0Tabla 3: Tabla <strong>de</strong> vida para E. pycnophylla a 3800 m <strong>de</strong> altitud (N=333)Número <strong>de</strong> Proporción <strong>de</strong> Mortalidad <strong>de</strong>Clases supervivientes individuos Probabilidad individuos Tasa <strong>de</strong><strong>de</strong> altura al inicio <strong>de</strong> la clase en cada clase <strong>de</strong> supervivencia entre clases mortalidad(x) (Nx) (p) (lx) (dx) (qx)1 153 0.459 1.000 68 0.4442 85 0.255 0.556 38 0.4473 47 0.141 0.307 32 0.6814 15 0.045 0.098 9 0.6005 6 0.018 0.039 -5 06 11 0.033 0.072 9 0.8187 2 0.006 0.013 -4 08 6 0.018 0.039 -2 09 8 0.024 0.052 8 0
Sánchez, 2004. Morfometría y Demografía <strong>de</strong> E. pycnophylla. 90Tabla 4: Tabla <strong>de</strong> vida para E. pycnophylla a 3900 m <strong>de</strong> altitud (N=475)Número <strong>de</strong> Proporción <strong>de</strong> Mortalidad <strong>de</strong>Clases supervivientes individuos Probabilidad individuos Tasa <strong>de</strong><strong>de</strong> altura al inicio <strong>de</strong> la clase en cada clase <strong>de</strong> supervivencia entre clases mortalidad(x) (Nx) (p) (lx) (dx) (qx)1 286 0.602 1.000 142 0.4972 144 0.303 0.503 120 0.8333 24 0.051 0.084 16 0.6674 8 0.017 0.028 0 0.0005 8 0.017 0.028 6 0.7506 2 0.004 0.007 2 1.0007 0 0.000 0.000 -1 08 1 0.002 0.003 -1 09 2 0.004 0.007 2 0Tabla 5: Tabla <strong>de</strong> vida para E. pycnophylla a 4000 m <strong>de</strong> altitud (N=395)Número <strong>de</strong> Proporción <strong>de</strong> Mortalidad <strong>de</strong>Clases supervivientes individuos Probabilidad individuos Tasa <strong>de</strong><strong>de</strong> altura al inicio <strong>de</strong> la clase en cada clase <strong>de</strong> supervivencia entre clases mortalidad(x) (Nx) (p) (lx) (dx) (qx)1 201 0.509 1.000 72 0.3582 129 0.327 0.642 113 0.8763 16 0.041 0.080 -1 04 17 0.043 0.085 4 0.2355 13 0.033 0.065 4 0.3086 9 0.023 0.045 4 0.4447 5 0.013 0.025 3 0.6008 2 0.005 0.010 -1 09 3 0.008 0.015 3 0Localidad Guan<strong>de</strong>raTabla 6: Tabla <strong>de</strong> vida para E. pycnophylla a 3500 m <strong>de</strong> altitud (N=325)Número <strong>de</strong> Proporción <strong>de</strong> Mortalidad <strong>de</strong>Clases supervivientes individuos Probabilidad individuos Tasa <strong>de</strong><strong>de</strong> altura al inicio <strong>de</strong> la clase en cada clase <strong>de</strong> supervivencia entre clases mortalidad(x) (Nx) (p) (lx) (dx) (qx)1 137 0.422 1.000 44 0.3212 93 0.286 0.679 50 0.5383 43 0.132 0.314 17 0.3954 26 0.080 0.190 14 0.5385 12 0.037 0.088 4 0.3336 8 0.025 0.058 5 0.6257 3 0.009 0.022 1 0.3338 2 0.006 0.015 1 0.5009 1 0.003 0.007 1 0
Sánchez, 2004. Morfometría y Demografía <strong>de</strong> E. pycnophylla. 91Tabla 7: Tabla <strong>de</strong> vida para E. pycnophylla a 3600 m <strong>de</strong> altitud (N=268)Número <strong>de</strong> Proporción <strong>de</strong> Mortalidad <strong>de</strong>Clases supervivientes individuos Probabilidad individuos Tasa <strong>de</strong><strong>de</strong> altura al inicio <strong>de</strong> la clase en cada clase <strong>de</strong> supervivencia entre clases mortalidad(x) (Nx) (p) (lx) (dx) (qx)1 157 0.586 1.000 84 0.5352 73 0.272 0.465 55 0.7533 18 0.067 0.115 8 0.4444 10 0.037 0.064 6 0.6005 4 0.015 0.025 1 0.2506 3 0.011 0.019 2 0.6677 1 0.004 0.006 -1 08 2 0.007 0.013 2 09 0 0 0 0 0Tabla 8: Tabla <strong>de</strong> vida para E. pycnophylla a 3700 m <strong>de</strong> altitud (N=459)Número <strong>de</strong> Proporción <strong>de</strong> Mortalidad <strong>de</strong>Clases supervivientes individuos Probabilidad individuos Tasa <strong>de</strong><strong>de</strong> altura al inicio <strong>de</strong> la clase en cada clase <strong>de</strong> supervivencia entre clases mortalidad(x) (Nx) (p) (lx) (dx) (qx)1 340 0.754 1.000 284 0.8352 56 0.124 0.165 33 0.5893 23 0.051 0.068 9 0.3914 14 0.031 0.041 5 0.3575 9 0.020 0.026 3 0.3336 6 0.013 0.018 -2 07 8 0.018 0.024 7 0.8758 1 0.002 0.003 -1 09 2 0.004 0.006 2 0Tabla 9: Tabla <strong>de</strong> vida para E. pycnophylla a 3800 m <strong>de</strong> altitud (N=473)Número <strong>de</strong> Proporción <strong>de</strong> Mortalidad <strong>de</strong>Clases supervivientes individuos Probabilidad individuos Tasa <strong>de</strong><strong>de</strong> altura al inicio <strong>de</strong> la clase en cada clase <strong>de</strong> supervivencia entre clases mortalidad(x) (Nx) (p) (lx) (dx) (qx)1 268 0.567 1.000 144 0.5372 124 0.262 0.463 83 0.6693 41 0.087 0.153 23 0.5614 18 0.038 0.067 5 0.2785 13 0.027 0.049 10 0.7696 3 0.006 0.011 -1 07 4 0.008 0.015 4 1.0008 0 0.000 0.000 -2 09 2 0.004 0.007 2 0
Sánchez, 2004. Morfometría y Demografía <strong>de</strong> E. pycnophylla. 92Localidad El Vola<strong>de</strong>roTabla 10: Tabla <strong>de</strong> vida para E. pycnophylla a 3600 m <strong>de</strong> altitud (N=453)Número <strong>de</strong> Proporción <strong>de</strong> Mortalidad <strong>de</strong>Clases supervivientes individuos Probabilidad individuos Tasa <strong>de</strong><strong>de</strong> altura al inicio <strong>de</strong> la clase en cada clase <strong>de</strong> supervivencia entre clases mortalidad(x) (Nx) (p) (lx) (dx) (qx)1 240 0.530 1.000 116 0.4832 124 0.274 0.517 94 0.7583 30 0.066 0.125 12 0.4004 18 0.040 0.075 3 0.1675 15 0.033 0.063 10 0.6676 5 0.011 0.021 -2 07 7 0.015 0.029 1 0.1438 6 0.013 0.025 -2 09 8 0.018 0.033 8 0Tabla 11: Tabla <strong>de</strong> vida para E. pycnophylla a 3700 m <strong>de</strong> altitud (N=252)Número <strong>de</strong> Proporción <strong>de</strong> Mortalidad <strong>de</strong>Clases supervivientes individuos Probabilidad individuos Tasa <strong>de</strong><strong>de</strong> altura al inicio <strong>de</strong> la clase en cada clase <strong>de</strong> supervivencia entre clases mortalidad(x) (Nx) (p) (lx) (dx) (qx)1 116 0.460 1.000 52 0.4482 64 0.254 0.552 40 0.6253 24 0.095 0.207 4 0.1674 20 0.079 0.172 3 0.1505 17 0.067 0.147 14 0.8246 3 0.012 0.026 0 0.0007 3 0.012 0.026 2 0.6678 1 0.004 0.009 -3 09 4 0.016 0.034 4 0Tabla 12: Tabla <strong>de</strong> vida para E. pycnophylla a 3800 m <strong>de</strong> altitud (N=427)Número <strong>de</strong> Proporción <strong>de</strong> Mortalidad <strong>de</strong>Clases supervivientes individuos Probabilidad individuos Tasa <strong>de</strong><strong>de</strong> altura al inicio <strong>de</strong> la clase en cada clase <strong>de</strong> supervivencia entre clases mortalidad(x) (Nx) (p) (lx) (dx) (qx)1 242 0.567 1.000 140 0.5792 102 0.239 0.421 73 0.7163 29 0.068 0.120 12 0.4144 17 0.040 0.070 0 0.0005 17 0.040 0.070 9 0.5296 8 0.019 0.033 1 0.1257 7 0.016 0.029 4 0.5718 3 0.007 0.012 1 0.3339 2 0.005 0.008 2 0
Sánchez, 2004. Morfometría y Demografía <strong>de</strong> E. pycnophylla. 93ANEXO 5. Tablas <strong>de</strong> vida para cada una <strong>de</strong> las localida<strong>de</strong>sTabla 1: Tabla <strong>de</strong> vida para E. pycnophylla en la Zona <strong>de</strong> la 23 (N=1959)Número <strong>de</strong> Proporción <strong>de</strong> Mortalidad <strong>de</strong>Clases supervivientes individuos Probabilidad individuos Tasa <strong>de</strong><strong>de</strong> altura al inicio <strong>de</strong> la clase en cada clase <strong>de</strong> supervivencia entre clases mortalidad(x) (Nx) (p) (lx) (dx) (qx)1 1083 0.553 1.0 558 0.5152 525 0.268 0.485 375 0.7143 150 0.077 0.139 71 0.4734 79 0.040 0.073 26 0.3295 53 0.027 0.049 22 0.4156 31 0.016 0.029 19 0.6137 12 0.006 0.011 0 08 12 0.006 0.011 -2 09 14 0.007 0.013 1 0Tabla 2: Tabla <strong>de</strong> vida para E. pycnophylla en el sitio <strong>de</strong> estudio Guan<strong>de</strong>ra (N=1525)Número <strong>de</strong> Proporción <strong>de</strong> Mortalidad <strong>de</strong>Clases Supervivientes individuos Probabilidad individuos Tasa <strong>de</strong><strong>de</strong> altura al inicio <strong>de</strong> la clase en cada clase <strong>de</strong> supervivencia entre clases mortalidad(x) (Nx) (p) (lx) (dx) (qx)1 902 0.591 1.0 556 0.6162 346 0.227 0.384 221 0.6393 125 0.082 0.139 57 0.4564 68 0.045 0.075 30 0.4415 38 0.025 0.042 18 0.4746 20 0.013 0.022 4 0.2007 16 0.010 0.018 11 0.6888 5 0.003 0.006 0 09 5 0.003 0.006 5 0Tabla 3: Tabla <strong>de</strong> vida para E. pycnophylla en la localidad <strong>de</strong> El Vola<strong>de</strong>ro (N=1132)Número <strong>de</strong> Proporción <strong>de</strong> Mortalidad <strong>de</strong>Clases supervivientes individuos Probabilidad individuos Tasa <strong>de</strong><strong>de</strong> altura al inicio <strong>de</strong> la clase en cada clase <strong>de</strong> supervivencia entre clases mortalidad(x) (Nx) (p) (lx) (dx) (qx)1 598 0.528 1.0 308 0.5152 290 0.256 0.485 207 0.7143 83 0.073 0.139 28 0.3374 55 0.049 0.092 6 0.1095 49 0.043 0.082 33 0.6736 16 0.014 0.027 -1 07 17 0.015 0.028 7 0.4128 10 0.009 0.017 -4 09 14 0.012 0.023 14 0