Forslag til besvarelse eksamen TFY4260 Cellebiologi og cellulær ...
Forslag til besvarelse eksamen TFY4260 Cellebiologi og cellulær ...
Forslag til besvarelse eksamen TFY4260 Cellebiologi og cellulær ...

Create successful ePaper yourself
Turn your PDF publications into a flip-book with our unique Google optimized e-Paper software.
<strong>Forslag</strong> <strong>til</strong> <strong>besvarelse</strong> <strong>eksamen</strong> <strong>TFY4260</strong> <strong>Cellebiol<strong>og</strong>i</strong> <strong>og</strong> cellulær biofysikk<br />
23. mai 2012<br />
Oppgave 1: Plasmamembranen. Aksjonspotensial. Transport av neurotransmittor<br />
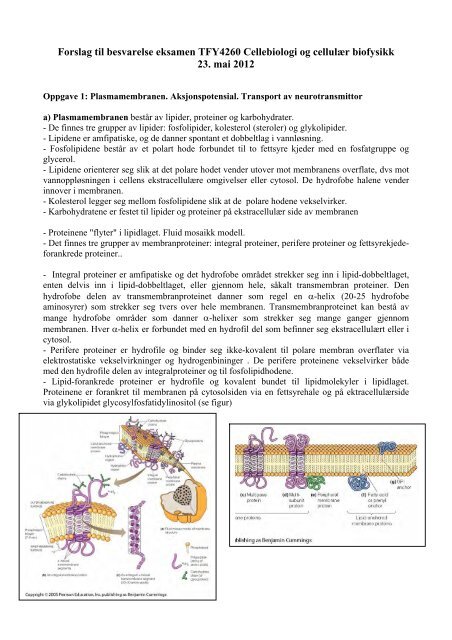
a) Plasmamembranen består av lipider, proteiner <strong>og</strong> karbohydrater.<br />
- De finnes tre grupper av lipider: fosfolipider, kolesterol (steroler) <strong>og</strong> glykolipider.<br />
- Lipidene er amfipatiske, <strong>og</strong> de danner spontant et dobbeltlag i vannløsning.<br />
- Fosfolipidene består av et polart hode forbundet <strong>til</strong> to fettsyre kjeder med en fosfatgruppe <strong>og</strong><br />
glycerol.<br />
- Lipidene orienterer seg slik at det polare hodet vender utover mot membranens overflate, dvs mot<br />
vannoppløsningen i cellens ekstracellulære omgivelser eller cytosol. De hydrofobe halene vender<br />
innover i membranen.<br />
- Kolesterol legger seg mellom fosfolipidene slik at de polare hodene vekselvirker.<br />
- Karbohydratene er festet <strong>til</strong> lipider <strong>og</strong> proteiner på ekstracellulær side av membranen<br />
- Proteinene "flyter" i lipidlaget. Fluid mosaikk modell.<br />
- Det finnes tre grupper av membranproteiner: integral proteiner, perifere proteiner <strong>og</strong> fettsyrekjedeforankrede<br />
proteiner..<br />
- Integral proteiner er amfipatiske <strong>og</strong> det hydrofobe området strekker seg inn i lipid-dobbeltlaget,<br />
enten delvis inn i lipid-dobbeltlaget, eller gjennom hele, såkalt transmembran proteiner. Den<br />
hydrofobe delen av transmembranproteinet danner som regel en -helix (20-25 hydrofobe<br />
aminosyrer) som strekker seg tvers over hele membranen. Transmembranproteinet kan bestå av<br />
mange hydrofobe områder som danner -helixer som strekker seg mange ganger gjennom<br />
membranen. Hver -helix er forbundet med en hydrofil del som befinner seg ekstracellulært eller i<br />
cytosol.<br />
- Perifere proteiner er hydrofile <strong>og</strong> binder seg ikke-kovalent <strong>til</strong> polare membran overflater via<br />
elektrostatiske vekselvirkninger <strong>og</strong> hydr<strong>og</strong>enbininger . De perifere proteinene vekselvirker både<br />
med den hydrofile delen av integralproteiner <strong>og</strong> <strong>til</strong> fosfolipidhodene.<br />
- Lipid-forankrede proteiner er hydrofile <strong>og</strong> kovalent bundet <strong>til</strong> lipidmolekyler i lipidlaget.<br />
Proteinene er forankret <strong>til</strong> membranen på cytosolsiden via en fettsyrehale <strong>og</strong> på ektracellulærside<br />
via glykolipidet glycosylfosfatidylinositol (se figur)<br />
b) Aksjonspotensial
) Aksjonspotensialet<br />
Ionestrømmer fra foregående aksjonspotensial induserer en depolarisering <strong>og</strong> et gradert potensial<br />
som igjen danner aksjonspotensialet dersom endringen i membranpotensial er større enn<br />
terskelverdien for dannelse av aksjonspotensial:<br />
Aksjonspotensialet dannes ved at:<br />
1.Spenningsfølsomme Na+ kanaler åpnes endringen i membranpotensial<br />
Na+ strømmer inn i cellen<br />
membranpotensialet depolariseres<br />
spenningsfølsomme Na+ kanaler åpnes <strong>og</strong><br />
membranpot. depol. ytterligere.<br />
2. Na+ kanaler inaktiveres.<br />
Dette skjer som en forsinket respons på endringen i membranpot<br />
Idet Na kanalene åpnes trigges <strong>og</strong>så inaktivering <strong>og</strong> påfølgende<br />
lukking av Na kanalene samt åpning av K+ kanaler (se figur)<br />
3. Spenningsfølsomme K+ kanaler åpnes<br />
K+ strømmer ut av cellen<br />
membranpotensialet går mot hvilepotensialet,<br />
hyperpolariseres, før hvilepotensialet gjenopprettes.<br />
Aksjonspotensialet forplanes langs axonet ved at ionestrømmer<br />
går langs axonet <strong>og</strong> depolariserer neste segment av axonet uten myelin eller <strong>til</strong> Ranviers knute i<br />
axon med myelin. Dette fører <strong>til</strong> at spenningsfølsomme Na+ kanaler <strong>og</strong> K+ kanaler åpnes <strong>og</strong> et nytt<br />
aksjonpotensial dannes.<br />
c) Transport langs axon<br />
Skjer ved at neurotransmittorer transporteres i vesikler langs miktotubulus.<br />
Vesikler transporters langs mikrotubulus ved hjelp av motoriske mikrotubulus assosierte proteiner som<br />
kinesin <strong>og</strong> cytoplasmatisk dynein. ( Kinesin går fra minus enden mot pluss enden, <strong>og</strong> cytoplasmatisk<br />
dynein går motsatt vei).<br />
De motoriske mikrotubulus assosierte proteinene består av to tunge kjeder tvunnet rundt hverandre, <strong>og</strong><br />
to lette kjeder ved COOH enden.<br />
Amino-enden av proteinene har to globulære hoder med ATPase aktivitet.<br />
Disse globulære hodene bindes <strong>til</strong> mikrotubulus.<br />
Vesikkelen bindes <strong>til</strong> COOH enden.<br />
2<br />
De motoriske proteinene beveger seg langs mikrotubulus ved bruk av ATP:<br />
1. ATP bindes <strong>til</strong> de motoriske proteinene konformasjonsendring av proteinene<br />
2. ATP ADP + Pi konformasjonsendring <strong>og</strong><br />
3. De motoriske proteinene beveger seg langs mikrotubulus<br />
4. ADP <strong>og</strong> Pi frigjøres konformasjonen gjenopprettes<br />
Det hele gjentas <strong>og</strong> gir en bevegelse i en retning
Oppgave 2: Proteintransport <strong>og</strong> modifisering (Vekttall 2)<br />
a) Glatt ER:<br />
• Lipidsyntese<br />
- Fosfolipider i membraner<br />
- Steroidehormoner fra kolesterol<br />
- Lipoprotein-partikler som frakter lipider i blodbanen<br />
• Detoxifiseringsreaksjoner<br />
- vannuløselig giftstoffer → vannløselige skilles ut i urinen.<br />
Skjer ved hydroksylering som avhenger av protein i cytokrom P450<br />
familien<br />
• Karbohydrat metabolisme<br />
- I absorptiv <strong>til</strong>stand (rett retter matinntak): glukose → gluk<strong>og</strong>en i lever celler<br />
- I postabsoptiv <strong>til</strong>stand: gluk<strong>og</strong>en → glukose<br />
• Lager av Ca 2+<br />
- Ca 2+ - ATPase pumper Ca 2+ inn i ER. I<br />
muskelceller sarcoplasmatisk reticulum.<br />
Ca 2+ må ut i cytosol for muskelkontraksjon.<br />
Ru ER:<br />
• Syntese av proteiner<br />
1) Membranproteiner<br />
2) Vannløselige proteiner:<br />
Proteiner som skal skilles ut av cellen<br />
Proteiner i lumen av organeller<br />
• Karbohydrater festes <strong>til</strong> proteiner<br />
• Kvalitetskontroll av proteiner: sjekker folding av proteiner <strong>og</strong> korreksjon for gal folding<br />
• Danner multisubenhet proteiner<br />
Hvilke funksjoner har glykosylering av proteiner:<br />
Intracellulært:<br />
- Markør for riktig protein folding<br />
- Markør for proteinsortering<br />
- Hindrer at proteaser bryter ned proteinet<br />
Ekstracellulært / i plasmamembranen:<br />
- Gjenkjennelse mellom celler<br />
- Reseptor for ekstracellulære molekyler<br />
- Hindrer angrep på cellen<br />
b) Protein sortering<br />
Transport av proteiner <strong>til</strong> membranen av endoplasmatisk reticulum:<br />
Proteiner som skal <strong>til</strong> ER har en signalsekvens (består av ca 20 aminosyrer) som oftest utgjør den<br />
første sekvensen på peptidkjeden. Se figuren.<br />
Denne signalsekvensen gjenkjennes av en signal-gjenkjennende partikkel (SRP) i cytosol, <strong>og</strong> binder<br />
seg <strong>til</strong> signalsekvensen. SRP sirkulerer mellom cytosol <strong>og</strong> membranen på ru ER. Proteinsyntesen<br />
stanser/går saktere så lenge SRP er bundet <strong>til</strong> signalsekvensen (skyldes sannsynligvis at partikkelen<br />
blokkerer for bindingsstedet for neste t-RNA).<br />
SRP fører ribosomet med peptidkjeden som har signalsekvensen <strong>til</strong> ER. På membranen på ru ER sitter<br />
en reseptor for SRP. Reseptoren er assosiert med et poreprotein som polypeptidkjeden kan gå gjennom.<br />
SRP binder seg <strong>til</strong> sin reseptor <strong>og</strong> dermed bindes ribosomet <strong>og</strong> peptidet <strong>til</strong> ER. Se figuren. GTP bindes<br />
<strong>til</strong> både SRP <strong>og</strong> SRP reseptor. Dette gjør at translasjonen fortsetter <strong>og</strong> signalsekvensen setter inn i pore<br />
3
proteinet <strong>og</strong> kanalen i poreproteinet åpnes. GTP hydrolyseres <strong>til</strong> GDP. Dette medfører at SRP løsner <strong>og</strong><br />
går <strong>til</strong>bake <strong>til</strong> cytosol. Proteinsyntesen fortsetter så på ER.<br />
4<br />
Proteinet passerer ER membranen:<br />
Proteinet passerer gjennom en kanal assosiert med reseptoren for signal-gjenkjennende partikkel. Poren<br />
er dynamisk dvs den åpnes når signalsekvensen som er hydrofob plasseres i membranen på ER.<br />
Proteinet syntetiseres <strong>og</strong> går samtidig gjennom poren (såkalt co-translasjon)<br />
Når hele proteinet har passert ER membranen, <strong>og</strong> det bare er bundet <strong>til</strong> membranen ved signalpeptidet.<br />
En peptidase "klipper" peptidkjeden av ved signalpeptidet. Signalsekvensen degraderes <strong>til</strong> aminosyrer<br />
<strong>og</strong> proteinet folder seg <strong>til</strong> sin native form.<br />
c) De to modellene for transport av lipider <strong>og</strong> proteiner gjennom golgi er:<br />
1. Stationary cisternae modell: Hvor hvert kompartment<br />
(Cisterna) av golgi er en stabil struktur. Trafikkering gjennom golgi<br />
foregår i denne modellen via transportvesikler som skilles ut fra en<br />
cisterna <strong>og</strong> vandrer videre <strong>til</strong> den neste med sitt innhold. Proteiner<br />
som skal <strong>til</strong> ulike deler av golgi, sekreteres ut eller sendes <strong>til</strong>bake <strong>til</strong><br />
ER blir fraktet med vesikler <strong>til</strong> sin lokalisering.<br />
2. Cisternal maturation modell: Hver cisterna går gradvis<br />
over fra å være i cis-golgi netverket <strong>til</strong> å bli trans-golgi i en<br />
modningsprosess. I modningsprosessen <strong>til</strong>føres hvert kompartment<br />
de nødvendige enzymer <strong>og</strong> lipider i en konstant prosess, <strong>og</strong> de<br />
holdes igjen i riktig kompartment av signaler i proteinene selv.<br />
Eksperimentelle data støtter begge modellene, men i den senere tid<br />
har fluorescens mikroskopi avslørt at modell 2 er den mest<br />
sannsynlige. Dette <strong>og</strong>så pga, størrelsen på en del molekyler som<br />
umulig kunne fraktes med små vesikler som er forutsetningen for<br />
modell 1.
5<br />
Oppgave 3<br />
a) cAMP aktivering<br />
Syntese av cAMP krever at enzymet adenylate cyclase aktiveres av G protein. G protein har fått sitt<br />
navn fordi det har GDP/GTP bundet <strong>til</strong> seg. G proteinet består av 3 polypeptidkjeder, , , <strong>og</strong> , <strong>og</strong><br />
er bundet <strong>til</strong> cytoplasmatisk side av plasmamembranen. Når GDP er bundet <strong>til</strong> -kjeden er de tre<br />
kjedene bundet <strong>til</strong> hverandre, <strong>og</strong> G proteinet er inaktivt. Når GTP erstatter GDP dissosierer -<br />
kjeden fra resten av G proteinet, <strong>og</strong> -kjeden aktiveres. Se figur.<br />
Adrenalin binder seg <strong>til</strong> sin reseptor på cellens plasmamembran. Dett er en reseptor av typen G-<br />
protein forbundet reseptor.<br />
reseptoren aktiveres, bindes <strong>til</strong> G proteinet <strong>og</strong> katalyserer reaksjonen som erstatter GDP bundet<br />
<strong>til</strong> G proteinet med GTP<br />
-kjeden løsner fra resten av G proteinet <strong>og</strong> bindingsstedet for adenylate cyclase på -kjeden<br />
eksponeres<br />
-kjeden bindes <strong>til</strong> enzymet adenylate cyclase som aktiveres<br />
adenylate cyclase katalyserer reaksjonen ATP cAMP. Se figur.<br />
Varighet av cAMP aktivering:<br />
-kjeden har GTPase aktivitet <strong>og</strong> hydrolyserer GTP GDP+Pi svært fort.<br />
Men så lengde adenalin sitter bundet <strong>til</strong> sin reseptor er denne aktivert <strong>og</strong> et nytt GTP vil erstatte<br />
GDP. Dvs at så lenge reseptoren er aktivert vil adenylate cyclase være aktiv <strong>og</strong> produsere cAMP.<br />
Funksjonen <strong>til</strong> cAMP: c-AMP virker ved å aktivere c-AMP avhengige protein kinaser, såkalte A-<br />
kinaser. Disse kinasene består av et proteinkompleks. Når c-AMP bindes gjennomgår<br />
proteinkomplekset en konformasjonsendring som frigjør <strong>og</strong> aktiverer de funksjonelle subenhetene av<br />
enzymet. Enzymet fosforylerer igjen andre intracellulære proteiner.<br />
b) Forsterkning av responsen<br />
Figuren nedenfor viser hvilke trinn der en forsterkning foregår.<br />
Ikke forsterkning i trinnet cAMP <strong>til</strong> kinase aktivering. 2 cAMP/ kinase
6<br />
- Mange G-protein kan aktiveres<br />
- Hvert G-protein kan aktiverer mange<br />
adenylate cyclaser<br />
- Hver adenylat cyclase kan syntetisere<br />
mange cAMP<br />
- Protein kinase A aktiverer mange<br />
fosforylase kinaser<br />
- Fosforylase kinase aktiverer mange<br />
glyc<strong>og</strong>en fosforylase<br />
-Fosforylase byter ned mange<br />
gluk<strong>og</strong>en <strong>til</strong> glucose-1-fosfa<br />
c) Tumor suppressor gener koder for:<br />
- p53 er cellens vakthund. Forhindrer at skadet DNA kan replikeres ved at celler med skadet DNA<br />
gjenomgår apoptose dersom DNA skaden ikke repareres. p53 fosforyleres <strong>og</strong> igangsetter to<br />
prosesser: arresterer cellen i G1 slik at den kan reparere DNA skaden. Dersom DNA skaden ikke<br />
repareres trigger p53 apoptose.<br />
-Rb er cellens bremse, bremser celledeling. Proteinet regulerer restriksjonspunktet <strong>og</strong> avgjør om<br />
cellen skal gå inni S-fase. Rb sjekker at nødvendige vekstbetingelser er oppfylt <strong>og</strong> egenskaper ved<br />
cellen som ingen DNA-skade <strong>og</strong> cellen må ha en viss størrelse. Dersom betingelsene er oppfylt<br />
fosforyleres Rb <strong>og</strong> enzymer nødvendig for å gjennomføre S-fase syntetiseres.<br />
- APC er et multiprotein ødeleggelse kompleks, involvert i Wnt sporet. APC bindes <strong>til</strong> proteinet<br />
Axin <strong>og</strong> glyc<strong>og</strong>en syntase kinase. Dette medfører fosforylering av β-catenin som merkes for<br />
nedbryting via proteasomer. Dette inaktiverer Wnt sporet. En mutasjon i APC gjør at β-catenin<br />
ikke brytes ned <strong>og</strong> Wnt sporet er alltid skrudd på <strong>og</strong> celle proliferasjon gjennomføres.<br />
Oppgave 4: Cytoskjelettet. Ekstracellulær matrix<br />
Strukturen av mikrovilli<br />
Tette bunter av aktin-filament holdt sammen av:<br />
- Aktinbindende proteiner: Fimbrin <strong>og</strong> villin<br />
-Festet <strong>til</strong> plamamembranen med myosin I <strong>og</strong> calmodulin<br />
-Plussenden utover i en tett amorf (ikke-krystallinsk) ukjent substans<br />
- Minusenden forankret i et nettverk i cytosol under plasmamembranen
7<br />
Strukturen av flimmerhår<br />
Flimmerhår består av mikrotubulus organisert i<br />
axonemer, 9 dubletter perifert i axonemet<br />
pluss to singletter sentralt som figur viser.<br />
To dubletter kan gli langs hverandre ved bruk<br />
av GTP bundet <strong>til</strong> proteinet dynein.<br />
Glidebevegelsen forhindres delvis a proteinet<br />
nexin som binder to dubletter sammen <strong>og</strong><br />
av ”eike” proteiner som binder dublettene <strong>til</strong><br />
de sentrale mikrotubulus.<br />
Hovedforskjeller:<br />
- Mikrovilli består av aktinfilament, flimmerhår av mikrotubulus<br />
Mikrovilli er statiske, flimmerhår dynamiske dvs de beveger seg.<br />
Funksjonen<br />
Mikrovilli: Øke absorpsjonsoverflaten : finnes på epitelcelle ri tarm der stor absorpsjonsfalte er<br />
nødvending. Mikrovilli på overflaten av epitelceller øker overflaten ca 20x.<br />
Flimmerhår: Beveger seg over epitelcellelag i luftveiene slik at støv <strong>og</strong> partikler fjernes.<br />
b) Fokalpunkter<br />
Cellens aktinfilament er forankret i ekstracellulær<br />
matrix (ECM) via fokalpunkter.<br />
Fokalpunktene består av aktinbindende proteiner<br />
(vinkulin <strong>og</strong> talin) som er festet <strong>til</strong><br />
transmembranproteiner kalt integriner. Integriner er<br />
bundet <strong>til</strong> ECM (collagen eller fibronektin). Se figur<br />
Hovedforskjellen mellom fokalpunkt <strong>og</strong> hemidesomosom er:<br />
- at i fokalpunktet er aktinfilamentet forankret <strong>til</strong> ECM, men i hemidesmosomer er intermediært<br />
filament forankret <strong>til</strong> ECM via integriner <strong>og</strong> intermediært filament er bundet <strong>til</strong> integrinet via plakk<br />
av proteiner.<br />
- Hemidesmosmer forbinder epitelceller <strong>til</strong> basalmembranen via laminin-reseptor som bindes <strong>til</strong><br />
proteinet laminin i basal membranen.
8<br />
c) Oppbyggingen av GAG <strong>og</strong> prote<strong>og</strong>lykaner<br />
Glykosamin<strong>og</strong>lykaner består av lange uforgreinete<br />
repeterende disakkarider. En av sukkerenhetene alltid er<br />
N-acetylglukosamin eller N-acetylgalakosamin. Den<br />
andre er ofte galaktose eller glukuronat<br />
De tre vanligst typene er keratan sulfate, kondroitin<br />
sulfate <strong>og</strong> hyalurononate. Hylauronoate skiller seg ut<br />
ved at det ikke inneholder sulfatgrupper.<br />
Prote<strong>og</strong>lykaner er GAG forbundet <strong>til</strong> et<br />
protein via en linker tetrasakkarid.<br />
Funksjoner:<br />
• Danner gel som fiberproteiner ”flyter” i.<br />
• Negative ladninger <strong>til</strong>trekker kationer <strong>og</strong> vann slik at gelen har et stort væsketrykk som motstår<br />
F ytre trykk krefter.<br />
• Regulerer trafikken/aktiviteten av celler, næringsstoffer/avfallsstoffer <strong>og</strong> molekyler (kjemiske<br />
signalmolekyl/enzymer) ved å begrense transport gjennom ekstracellulær matrix <strong>og</strong> binde<br />
signalmolekyler/proteiner<br />
Fibronektin er et adhesivt protein bestående av mange domener med bindingssteder <strong>til</strong> ulike<br />
bestandeler i ECM. I <strong>til</strong>legg inneholder det peptidsekvensen RGD som bindes <strong>til</strong> integrinet<br />
fibronektin reseptor.<br />
Oppgave 5: Immunol<strong>og</strong>i<br />
Klonal seleksjonsteorien for B-celler.<br />
1. Hver B-celle lages for å syntetisere et antistoff<br />
B-celler lages av stamceller <strong>og</strong> hver celle<br />
gjennomgår DNA rearrangering som gir opphave <strong>til</strong><br />
genet for antistoffet som hver B-celle kan produsere.<br />
Dette gir opphav <strong>til</strong> et enormt antall forskjellige<br />
antistoffer<br />
2. B-cellene produserer antistoff uten at de<br />
stimuleres med antigen.<br />
B-cellene produserer antistoffet <strong>og</strong> det<br />
plasseres på overflaten, med det<br />
antigengjenkjennende delen ut. De fleste B-cellene<br />
trengs ikke, men det store repertoaret gjør at ”alle”<br />
antigen kan detekteres.<br />
3. B-celler selekteres av antigenbinding <strong>og</strong> B-cellen aktiveres noe som gir massiv<br />
antistoffproduksjon.
B-cellene aktiveres av antigen. Enten av bare antigen eller som oftest <strong>og</strong>så sammen med T-<br />
hjelpeceller (CD4-T-celler). Aktiveringen fører <strong>til</strong> proliferasjon av den ene B-cellen, som gir en<br />
økning i antallet B-celler (danner en klon) som lager det riktige antistoffet. De aktiverte B-cellene<br />
utvikles <strong>til</strong> to celletyper.<br />
Den ene celletypen kalles plasmaceller <strong>og</strong> produserer <strong>og</strong> sekreterer store mengder antistoff. For å få<br />
<strong>til</strong> denne prosessen har plasma-cellen store mengder ru ER for å sikre stor proteinproduksjon g<br />
effektiv sekresjon. Plasmaceller lever kun kort tid etter at antigenet er forsvunnet.<br />
Vaksine<br />
Den andre typen B-celler er hukommelsesceller. Disse lager ikke antistoff, men aktiveres av<br />
antigenet om det møter det igjen. Denne aktiveringen er raskere enn den initiale aktiveringen, <strong>og</strong><br />
ved aktiveringen vil en del av hukommelses B-cellene proliferer <strong>og</strong> bli plasmaceller som kan<br />
produsere antistoff. Den sekundært immunresponsen er mye raskere <strong>og</strong> vil kun ta timer, men den<br />
initiale responsen tar flere dager. Hukommelses B-cellene er bakgrunnen for at vi kan lage vaksiner<br />
for en del bakterier <strong>og</strong> virus. Vi injiseres da med et antigen vi ønsker å beskytte oss mot.<br />
Oppgave 6 (Vekttall 1)<br />
I denne oppgaven får dere angitt 3 svar, hvorav ett er riktig. Sett kryss ved siden av det riktige<br />
svaret. Lever oppgavearket merket med studiepr<strong>og</strong>ram, studentnr, <strong>og</strong> sidetall.<br />
a) Dersom plasmamembranen ble permeabel for Na + <strong>og</strong> K + , hva ville Na + /K + pumpa gjøre:<br />
- Hemmes å slutte <strong>og</strong> pumpe<br />
- Pumpe Na + <strong>og</strong> K + i begge retninger<br />
- Fortsette å pumpe Na + ut av cellen <strong>og</strong> K + inn i cellen, <strong>og</strong> hydrolysere ATP, men energien<br />
ved hydrolyse vil være bortkastet idet varme produseres istedenfor ionegradient.<br />
b) Proteiner passerer membranene av mitokondrier:<br />
- i en foldet <strong>til</strong>stand<br />
- ufoldet ved kotranslasjon<br />
- ufoldet etter at translasjonen er avsluttet<br />
c) Histoner passerer gjennom kjerneporekomplekset ved:<br />
- Passiv diffusjon<br />
- Krever ATP<br />
- Krever G-bindende protein<br />
d) Kromatidtrådene trekkes <strong>til</strong> hver sin spindelpol i:<br />
- metafase<br />
- anafase<br />
- telofase<br />
e) Under celledeling brytes kjernekonvolutten opp i:<br />
- Interfase<br />
- Profase<br />
- Prometafase<br />
f) RNA syntetiseres gjennom cellesyklus i:<br />
- Hele interfasen<br />
- G1-fasen<br />
- S-fasen<br />
9
10<br />
g) rRNA syntese foregår:<br />
- i nukleolus<br />
- i ribosomer i cytosol<br />
- i endoplasmatisk reticulum<br />
h) Organellen som sannsynligvis stammer fra en annen organisme er:<br />
- Lysosomer<br />
- Golgi apparatet<br />
- Mitokondrier<br />
i) Sorteringssignalet bestemmer hvor i cellen et protein skal transporteres.<br />
Til hvor i cellen vil et protein uten sorteringssignal bli transportert?<br />
- Kjernen<br />
- Cytosol<br />
- Mitokondriene<br />
j) Du ønsker å detektere karbohydrater på plasmamembranen. Hvilket av de følgende molekylene<br />
kan da benyttes:<br />
- Lektiner<br />
- Cadheriner<br />
- Integriner<br />
k) v-SNARE deltar direkte i<br />
- dannelse av transportvesikler<br />
- transporten av vesikler<br />
- bindingen av vesikler <strong>til</strong> target organelle<br />
l) Kontraksjon av glatt muskulatur krever at intracellulær konsentrasjon av Ca 2+ øker. Dette skjer<br />
ved at Ca 2+ ionekanaler åpnes i:<br />
- Plasmamembranen<br />
- Mitochondria<br />
- Sarcoplasmatisk reticulum<br />
m) I genregulering er lac-operonet mye studert. Hva skjer om en muterer lac-repressoren slik at den<br />
ikke lenger kan binde laktose, men fortsatt kan binde seg <strong>til</strong> operatoren?<br />
- Ingen transkripsjon av lac-genene.<br />
- Full transkripsjon av genen.<br />
- Høy nedbryting av laktose<br />
n) Etter transkripsjon av DNA i en eukaryot celle modifiseres primærtranskriptet <strong>til</strong> et mRNAmolekyl.<br />
Hvilke av utsagnene under er rett beskrivelse av modifikasjonen av primærtranskriptet?<br />
- Primærtranskriptet kuttes <strong>og</strong> limes (ligeres) sammen på nytt uten introner<br />
- Primærtranskriptet kuttes <strong>og</strong> limes (ligeres) sammen på nytt men uten exoner.<br />
- Primærtranskriptet limes (ligeres) sammen med en polyAdenosin hale.<br />
o) Enzymer gjennomgår ofte post-translasjonelle endringer som en del av regulering av<br />
enzymaktiviteten. Hvilken av de følgende endringene vil være irreversible?<br />
- Aktivering ved fosforylering<br />
- Proteolytisk aktivering av proenzym<br />
- Poly-ubiquitinylering
11<br />
p)I hvilken <strong>til</strong>stand foreligger DNA hovedsakelig i cellekjernen?<br />
- Bundet <strong>til</strong> oktamerer av histon <strong>og</strong> flere typer ikke-histon proteiner<br />
- I fri <strong>til</strong>stand uten å være bundet <strong>til</strong> proteiner<br />
- Tett bundet rundt ribosomale proteiner<br />
q) Hva er en transkripsjonsfaktor?<br />
- DNA-bindende protein som regulerer genavlesingen<br />
- RNA-bindende protein som regulerer proteinsyntesen<br />
- Kompleksdannende protein som regulerer nedbryting av transfer-RNA<br />
r) Kroppens forsvar karakteriseres ved at aktivering av nødvendige celler ofte krever to signaler.<br />
Hva er den beste forklaringen på at dette er hensiktsmessig?<br />
- Det gir større sikkerhet mot feilaktig aktivering<br />
- Det gir bedre målstyring av responsene<br />
- Det gir bedre regulering av tempoet i aktiveringen