The phylogenetic distribution of resupinate forms ... - Clark University
The phylogenetic distribution of resupinate forms ... - Clark University
The phylogenetic distribution of resupinate forms ... - Clark University

Create successful ePaper yourself
Turn your PDF publications into a flip-book with our unique Google optimized e-Paper software.
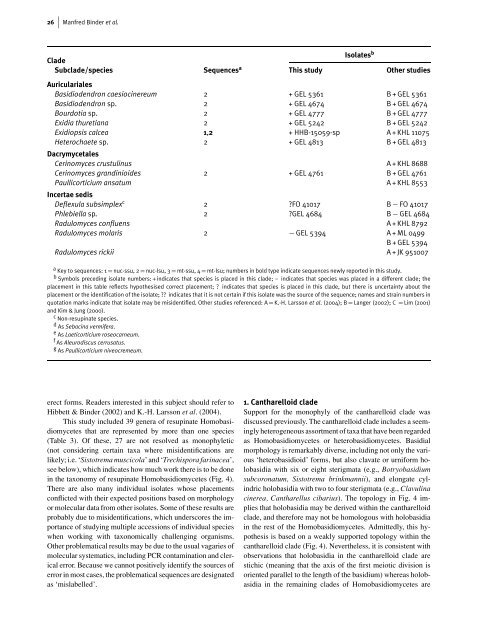
26 Manfred Binder et al.<br />
Isolates b<br />
Clade<br />
Subclade/species Sequences a This study Other studies<br />
Auriculariales<br />
Basidiodendron caesiocinereum 2 + GEL 5361 B + GEL 5361<br />
Basidiodendron sp. 2 + GEL 4674 B + GEL 4674<br />
Bourdotia sp. 2 + GEL 4777 B + GEL 4777<br />
Exidia thuretiana 2 + GEL 5242 B + GEL 5242<br />
Exidiopsis calcea 1,2 + HHB-15059-sp A + KHL 11075<br />
Heterochaete sp.<br />
Dacrymycetales<br />
2 + GEL 4813 B + GEL 4813<br />
Cerinomyces crustulinus A + KHL 8688<br />
Cerinomyces grandinioides 2 + GEL 4761 B + GEL 4761<br />
Paullicorticium ansatum<br />
Incertae sedis<br />
A + KHL 8553<br />
Deflexula subsimplexc 2 ?FO 41017 B − FO 41017<br />
Phlebiella sp. 2 ?GEL 4684 B − GEL 4684<br />
Radulomyces confluens A + KHL 8792<br />
Radulomyces molaris 2 − GEL 5394 A + ML 0499<br />
B + GEL 5394<br />
Radulomyces rickii A + JK 951007<br />
a Key to sequences: 1 = nuc-ssu, 2 = nuc-lsu, 3 = mt-ssu, 4 = mt-lsu; numbers in bold type indicate sequences newly reported in this study.<br />
b Symbols preceding isolate numbers: + indicates that species is placed in this clade; – indicates that species was placed in a different clade; the<br />
placement in this table reflects hypothesised correct placement; ? indicates that species is placed in this clade, but there is uncertainty about the<br />
placement or the identification <strong>of</strong> the isolate; ?? indicates that it is not certain if this isolate was the source <strong>of</strong> the sequence; names and strain numbers in<br />
quotation marks indicate that isolate may be misidentified. Other studies referenced: A = K.-H. Larsson et al. (2004); B = Langer (2002); C = Lim (2001)<br />
and Kim & Jung (2000).<br />
c Non-<strong>resupinate</strong> species.<br />
d As Sebacina vermifera.<br />
e As Laeticorticium roseocarneum.<br />
f As Aleurodiscus cerrusatus.<br />
g As Paullicorticium niveocremeum.<br />
erect <strong>forms</strong>. Readers interested in this subject should refer to<br />
Hibbett & Binder (2002) and K.-H. Larsson et al. (2004).<br />
This study included 39 genera <strong>of</strong> <strong>resupinate</strong> Homobasidiomycetes<br />
that are represented by more than one species<br />
(Table 3). Of these, 27 are not resolved as monophyletic<br />
(not considering certain taxa where misidentifications are<br />
likely; i.e. ‘Sistotrema muscicola’and‘Trechispora farinacea’,<br />
see below), which indicates how much work there is to be done<br />
in the taxonomy <strong>of</strong> <strong>resupinate</strong> Homobasidiomycetes (Fig. 4).<br />
<strong>The</strong>re are also many individual isolates whose placements<br />
conflicted with their expected positions based on morphology<br />
or molecular data from other isolates. Some <strong>of</strong> these results are<br />
probably due to misidentifications, which underscores the importance<br />
<strong>of</strong> studying multiple accessions <strong>of</strong> individual species<br />
when working with taxonomically challenging organisms.<br />
Other problematical results may be due to the usual vagaries <strong>of</strong><br />
molecular systematics, including PCR contamination and clerical<br />
error. Because we cannot positively identify the sources <strong>of</strong><br />
error in most cases, the problematical sequences are designated<br />
as ‘mislabelled’.<br />
1. Cantharelloid clade<br />
Support for the monophyly <strong>of</strong> the cantharelloid clade was<br />
discussed previously. <strong>The</strong> cantharelloid clade includes a seemingly<br />
heterogeneous assortment <strong>of</strong> taxa that have been regarded<br />
as Homobasidiomycetes or heterobasidiomycetes. Basidial<br />
morphology is remarkably diverse, including not only the various<br />
‘heterobasidioid’ <strong>forms</strong>, but also clavate or urniform holobasidia<br />
with six or eight sterigmata (e.g., Botryobasidium<br />
subcoronatum, Sistotrema brinkmannii), and elongate cylindric<br />
holobasidia with two to four sterigmata (e.g., Clavulina<br />
cinerea, Cantharellus cibarius). <strong>The</strong> topology in Fig. 4 implies<br />
that holobasidia may be derived within the cantharelloid<br />
clade, and therefore may not be homologous with holobasidia<br />
in the rest <strong>of</strong> the Homobasidiomycetes. Admittedly, this hypothesis<br />
is based on a weakly supported topology within the<br />
cantharelloid clade (Fig. 4). Nevertheless, it is consistent with<br />
observations that holobasidia in the cantharelloid clade are<br />
stichic (meaning that the axis <strong>of</strong> the first meiotic division is<br />
oriented parallel to the length <strong>of</strong> the basidium) whereas holobasidia<br />
in the remaining clades <strong>of</strong> Homobasidiomycetes are