- Page 2 and 3:
Rice Genetics II Proceedings of the
- Page 4 and 5:
Contents Foreword xiii K. J. Lampe
- Page 6 and 7:
Thermosensitive genetic male steril
- Page 8 and 9:
Intraspecific variation and genetic
- Page 10 and 11:
Restriction fragment length polymor
- Page 12 and 13:
Simple, rapid procedure for analyzi
- Page 14:
Foreword The First International Ri
- Page 17 and 18:
The First International Rice Geneti
- Page 20 and 21:
International cooperation in rice g
- Page 22 and 23:
Chromosomes were numbered in decrea
- Page 24 and 25:
Establishment of the Rice Genetics
- Page 26 and 27:
Abbreviations and acronyms A ABA AC
- Page 28:
S SCA = specific combining ability
- Page 32 and 33:
Association between Pox-1 variation
- Page 34 and 35:
Near-isogenic lines of Pox-1 Ten F
- Page 36 and 37:
Table 3. Association between charac
- Page 38 and 39:
2. Comparison of character measurem
- Page 40:
Fu P Y, Pai C (1979) Genetic studie
- Page 43 and 44:
protein, since a wx mutant (75-1) p
- Page 45 and 46:
2. Heterogeneity of Wx proteins (sp
- Page 47 and 48:
Altered gene expression As mentione
- Page 49 and 50:
Table 3. Distribution of nonwaxy al
- Page 51 and 52:
Notes Authors’ addresses: Y. Sano
- Page 53 and 54:
designation to “tropical japonica
- Page 55 and 56:
Table 2. Pollen and spikelet fertil
- Page 57 and 58:
Table 4. Mean similarity indices an
- Page 59 and 60:
Root thickness. The upland and aus
- Page 61 and 62:
The aus group is also closely relat
- Page 63 and 64:
Notes Authors’ addresses: T.T. Ch
- Page 65 and 66:
to identify the wide compatibility
- Page 67 and 68:
Hybrids between aus varieties Becau
- Page 69 and 70:
Genetic analysis of hybrid sterilit
- Page 71 and 72:
Table 5. Detection of wide compatib
- Page 73 and 74:
Discussion The system of F 1 hybrid
- Page 76 and 77:
How was rice differentiated into in
- Page 78 and 79:
1. Relations among phenol reaction,
- Page 80 and 81:
3. Association of 9 genes and chara
- Page 82 and 83:
The pattern of association among th
- Page 84:
Oka H I (1974) Analysis of genes co
- Page 87 and 88:
Table 1. Varieties used in crossing
- Page 89 and 90:
1. Hierarchical ascendant classific
- Page 91 and 92:
three restriction patterns, while a
- Page 93 and 94:
Table 4. Distorted F 2 segregations
- Page 95 and 96:
explain the correspondence between
- Page 98 and 99:
Traditional highland rices originat
- Page 100 and 101:
Morphophysiological variability The
- Page 102 and 103:
2. Distribution of varieties from M
- Page 104 and 105: Locus Allele Whole sample Indica gr
- Page 106 and 107: For the japonica group, Madagascar
- Page 108 and 109: Thus, rice introduction in Madagasc
- Page 110: Rabary E, Noyer J L, Benyayer P, Ar
- Page 113 and 114: The spontaneous hybrids were called
- Page 115 and 116: Table 1. Mean outcrossing rates (m)
- Page 117 and 118: Table 3. Comparison of 1) ratios of
- Page 119 and 120: fertility and to eliminate unfavora
- Page 122 and 123: Discussion Session 1: Varietal diff
- Page 124 and 125: Q— Chang : The earliest remains o
- Page 126: A— Morishima : Since most of the
- Page 130 and 131: Analysis of genes for stigma colora
- Page 132 and 133: Table 1. Varieties used as parents
- Page 134 and 135: Underlying genes in these different
- Page 136 and 137: (121:9, 2 homogeneous crosses poole
- Page 138 and 139: Table 6. Linkage relations estimate
- Page 140 and 141: Table 9. Recombination values obtai
- Page 142 and 143: In this study, inhibitors for stigm
- Page 144 and 145: Gene mapping of some morphological
- Page 146 and 147: • Liguleless-a. The auricle and l
- Page 148 and 149: Table 3. Linkage between sheathed p
- Page 150 and 151: Table 6. Linkage between short ligu
- Page 152 and 153: 3. Linkage relationships between ne
- Page 156 and 157: 1. Linkage map of 5 genes including
- Page 158: Notes Authors’ addresses: H.S. Su
- Page 161 and 162: Table 1. Rice isozyme genes and cor
- Page 163 and 164: 10 5 5 8 6 14 18 20 34 15 6.13 (0.5
- Page 165 and 166: 1. Linkage maps of 11 isozyme genes
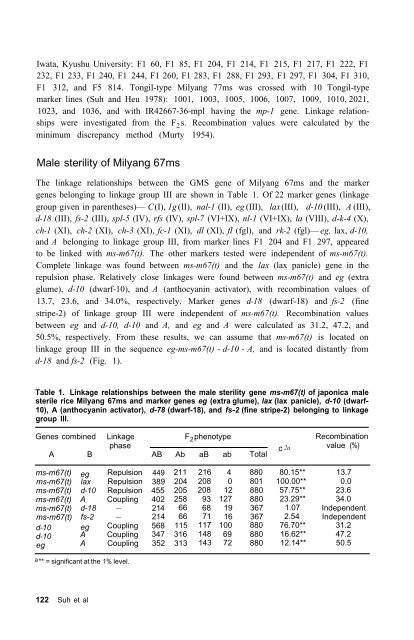
- Page 167 and 168: Table 5. Number of variant subcell
- Page 169 and 170: Ranjhan S, Glaszmann J C, Ramirez D
- Page 171 and 172: environment, have pleiotropic effec
- Page 173 and 174: Table 3. Segregation for Enp-1 in B
- Page 175 and 176: Linkage relationship between isozym
- Page 177 and 178: Table 8. Summary of linkage relatio
- Page 180 and 181: Production of monosomic alien addit
- Page 182 and 183: 1. Development of O. sativa MAALs h
- Page 184 and 185: MAAL H, which corresponded to the h
- Page 186 and 187: 4. a) PMC of MAAL N showing 12 II +
- Page 188: Jena K K, Khush G S (1989) Monosomi
- Page 192: SESSION 3 Genetics of Stress Tolera
- Page 195 and 196: Knowledge of the genetics of drough
- Page 197 and 198: esistant plants unrolled. The resis
- Page 199 and 200: Table 3. Leaf rolling and unrolling
- Page 201 and 202: IRRI—International Rice Research
- Page 203 and 204: Materials and methods Genetic analy
- Page 205 and 206:
Table 3. Estimates of genetic param
- Page 207 and 208:
1. Grain yield per plant. 1 = IR20,
- Page 210 and 211:
Inheritance of panicle exsertion in
- Page 212:
Notes Authors’ address: D.M. Mahi
- Page 215 and 216:
parallel to normal soil. Do you thi
- Page 218 and 219:
Inheritance of grain size and its i
- Page 220 and 221:
size may occur. Takeda (1983) found
- Page 222 and 223:
genes allelic to Lk-f. These result
- Page 224 and 225:
Grain size in relation to yield To
- Page 226:
Takeda K, Saito K (1980) Major gene
- Page 229 and 230:
inhibitors were recovered, plants r
- Page 231 and 232:
corresponding amides. Cysteine was
- Page 233 and 234:
4. Relationships among seed weight,
- Page 235 and 236:
tate was significantly higher in th
- Page 238 and 239:
Mutants for rice storage proteins T
- Page 240 and 241:
1. SDS-PAGE analysis of salt-insolu
- Page 242 and 243:
Table 1. Content of proteins extrac
- Page 244 and 245:
Table 3. Segregation of normal and
- Page 246:
Kumamaru T, Satoh H, Iwata N, Omura
- Page 249 and 250:
Table 1. Lines carrying late-headin
- Page 251 and 252:
Crosses between late-heading lines
- Page 254 and 255:
Photoperiod-conditioned male steril
- Page 256 and 257:
Table 1. Daily changes in pollen fe
- Page 258 and 259:
Plants (%) 1. Distribution of NK58s
- Page 260 and 261:
generative nucleus degenerates, wit
- Page 262 and 263:
Table 6. Segregation for spikelet f
- Page 264 and 265:
Thermosensitive genetic male steril
- Page 266 and 267:
Table 3. Seed set percentage and fl
- Page 268 and 269:
Table 8. Fertility of H89-1 grown u
- Page 270 and 271:
Discussion Session 4: Genetics of m
- Page 272:
Q— Virmani: IRRI is thankful to J
- Page 276 and 277:
Transfer of blast and bacterial bli
- Page 278 and 279:
inocula in the International Rice B
- Page 280 and 281:
1. Blast lesions on a) Oryza sativa
- Page 282 and 283:
Table 4. Reaction of WHD IS 78-1 BC
- Page 284 and 285:
minuta Acc. 101141 has been shown t
- Page 286 and 287:
Inheritance of resistance to rice t
- Page 288 and 289:
203 is recessive. The F 2 populatio
- Page 290 and 291:
esults of Pankhari 203/TN1 showed a
- Page 292 and 293:
RTSV can spread as an independent v
- Page 294 and 295:
Genetics of rice resistance to brow
- Page 296 and 297:
Table 1. Reaction to BPH of F 1 s a
- Page 298 and 299:
esistant:8 segregating:1 susceptibl
- Page 300:
Data on the relative contribution o
- Page 303 and 304:
88 (using ADR52 and N’Diang Marie
- Page 305 and 306:
Crosses with ADR52 and N’Diang Ma
- Page 308:
Discussion Session 5: Genetics of d
- Page 312 and 313:
Application of somaclonal variation
- Page 314 and 315:
Unfavorable conditions induced some
- Page 316 and 317:
tion. Root and leaf growth was prog
- Page 318:
in vitro selection at the haploid l
- Page 321 and 322:
information on the inheritance of c
- Page 323 and 324:
espectively). Average frequencies o
- Page 325 and 326:
3. (Vr, Wr) graph for plant regener
- Page 327 and 328:
Table 6. Analysis of variance of 6
- Page 330 and 331:
Genetic analysis for salt tolerance
- Page 332 and 333:
Table 1. Regenerants from IR20 and
- Page 334 and 335:
Table 5. Fertile R 1 progeny lines
- Page 336 and 337:
Somaclonal male sterile mutants and
- Page 338 and 339:
sterile plants in the R 1 and R 2 w
- Page 340 and 341:
Table 2. Segregation patterns for s
- Page 342 and 343:
When Zhen shan 97 B and Er Jiu-ai B
- Page 344:
Oono K (1985) Putative homozygous m
- Page 347 and 348:
and exhibiting high regenerative ca
- Page 349 and 350:
Table 3. Detection of ß-glucuronid
- Page 351 and 352:
Lee L, Schroll R E, Grimes H D, Hod
- Page 353 and 354:
advantageous than wing somatic cell
- Page 355 and 356:
2. Division frequency of pollen at
- Page 357 and 358:
4. Effect of exposure time at diffe
- Page 359 and 360:
Table 4. Effect of quality and quan
- Page 362 and 363:
Cryopreservation: a method for main
- Page 364 and 365:
Table 1. Cryoprotectant and freezin
- Page 366 and 367:
treatments (Fig. 1b, c, d) was simi
- Page 368 and 369:
3. Post-freeze respiration of pretr
- Page 370 and 371:
after freezing. The membrane carrie
- Page 372 and 373:
References cited Abdullah R, Cockin
- Page 374 and 375:
Discussion Session 6: Tissue and ce
- Page 376 and 377:
Q— Zheng Kangle: I wonder if male
- Page 378:
SESSION 7 Molecular Genetics of Cyt
- Page 381 and 382:
1. Nucleotide sequence of 32-kDa qu
- Page 383 and 384:
2. Genetic and physical map of ctDN
- Page 385 and 386:
Kanno A, Hirai A (1989) The nucleot
- Page 387 and 388:
number of these genes have been obt
- Page 389 and 390:
346 André et al
- Page 391 and 392:
2. Annotated sequence of IR36 cox 3
- Page 393 and 394:
Table 1. Codon usage for 3 rice mit
- Page 395 and 396:
ase recognition enzyme and hence is
- Page 397 and 398:
Notes Authors’ address: C. Andé,
- Page 399 and 400:
pathogen Helminthosporium maydis, r
- Page 401 and 402:
cytoplasm of a naturally occurring
- Page 403 and 404:
Generally, in vitro cultures genera
- Page 405 and 406:
Chowdhury M K U, Schaeffer G W, Smi
- Page 407 and 408:
Small I D, Earle E D, Escote-Carlso
- Page 409 and 410:
origin. Kadowaki et al (1988) exami
- Page 411 and 412:
Table 1. Presence of mitochondrial
- Page 413 and 414:
emarkable polymorphisms were observ
- Page 415 and 416:
Nakagahra M (1972) Genetic mechanis
- Page 417 and 418:
Q— Ranjekar: Are there any variat
- Page 420:
SESSION 8 Molecular Genetics of Nuc
- Page 423 and 424:
element has a very high G+C content
- Page 425 and 426:
3. Nucleotide sequence alignment of
- Page 427 and 428:
4c). However, no messenger RNA can
- Page 429 and 430:
6. Genomic blot analysis of IR36 (T
- Page 431 and 432:
This gives a ratio of 4.6, which is
- Page 434 and 435:
Genetic variation in tissue culture
- Page 436 and 437:
1. Genomic DNA was isolated from ri
- Page 438 and 439:
3. Genomic DNA from regenerated ric
- Page 440 and 441:
Table 2. RFLP analysis of rice plan
- Page 442 and 443:
5. Genomic DNA was isolated from ca
- Page 444 and 445:
References cited Baker A, Leaver C
- Page 446 and 447:
Characterization of repeated DNA se
- Page 448 and 449:
1. Hybridization with AA-specific 3
- Page 450 and 451:
were observed, suggesting that a te
- Page 452:
References cited Cordesse F, Second
- Page 455 and 456:
markers may facilitate selection du
- Page 457 and 458:
1. Genomic DNA of rice species dige
- Page 459 and 460:
4. Hybridization of pOm6 to Eco RI-
- Page 461 and 462:
Notes Authors’ addresses: H. Aswi
- Page 463 and 464:
still unclear how many isozymes are
- Page 465 and 466:
2. Restriction map of 5 rice chromo
- Page 467 and 468:
a Horizontal clones are abbreviated
- Page 469 and 470:
(Huttly et al 1988), the Amy2 subfa
- Page 471 and 472:
5. Summary of rice a -amylase genes
- Page 473 and 474:
intron is not always present. One p
- Page 476 and 477:
Discussion Session 8: Molecular gen
- Page 478:
Q— Ranjekar: a -amylase is a mult
- Page 482 and 483:
RFLP mapping of the rice genome S.D
- Page 484 and 485:
for plant species with comparable g
- Page 486 and 487:
on 113 backcross individuals, and t
- Page 488 and 489:
ice is the major grain crop. To fac
- Page 490 and 491:
Tagging genes for disease and insec
- Page 492 and 493:
Table 2. Summary of resistance char
- Page 494 and 495:
mapped. 2) We clone unique, or sing
- Page 496:
Kelemu S, Leach J E (1990) Cloning
- Page 499 and 500:
Recently developed restriction frag
- Page 501 and 502:
The majority of polymorphic clones,
- Page 503 and 504:
2. Map of main linkage group of ric
- Page 505 and 506:
Valent B, Farrall L, Chumley F G (1
- Page 507 and 508:
ecently, a linkage map of RFLP mark
- Page 509 and 510:
2. Segregation of some RFLP markers
- Page 511 and 512:
to the 12 chromosomes (Fig. 3). For
- Page 514 and 515:
Intraspecific variation and genetic
- Page 516 and 517:
library from Nipponbare, a Japanese
- Page 518 and 519:
Intraspecific differentiation in ri
- Page 520:
Oka H I (1953a) Phylogenetic differ
- Page 523 and 524:
ice (summarized in Second 1985b). T
- Page 525 and 526:
Ava I were studied in all plants, a
- Page 527 and 528:
longistaminata occurred (Second 198
- Page 529 and 530:
Mitochondrial DNA RFLP in section O
- Page 531 and 532:
SATIVA GROUP (Genome A) Ancestral s
- Page 533 and 534:
Dally AM, Second G (1989) Chloropla
- Page 535 and 536:
genetics of P. oryzae and P. grisea
- Page 537 and 538:
Valent et al 1986, Yaegashi and Asa
- Page 539 and 540:
[Gabriel 1989] and fungi [Leong and
- Page 541 and 542:
Table 2. Similarities between Pyric
- Page 543 and 544:
Kato H, Yamaguchi T (1980) Host ran
- Page 546 and 547:
Discussion Session 9: RFLP analysis
- Page 548:
all sequences are initially duplica
- Page 552 and 553:
Rice endosperm proteins and their a
- Page 554 and 555:
osmium tetraoxide staining, is obse
- Page 556 and 557:
3. Structures of signal sequences o
- Page 558 and 559:
endosperm cells. This figure is the
- Page 560:
Masumura T, Hibino T, Kidzu K, Mits
- Page 563 and 564:
Table 1. Composition of rice (Oryza
- Page 565 and 566:
1. Effect of ethanol concentration
- Page 567 and 568:
3. ELISA of rice albumin using vari
- Page 569 and 570:
against 70% ethanol. Using those im
- Page 571 and 572:
Table 1. Rice species used. Species
- Page 573 and 574:
Table 2. Rice DNA bands hybridized
- Page 575 and 576:
Takaiwa F, Kikuchi S, Oono K (1988)
- Page 578:
SESSION 11 Molecular Genetics of Di
- Page 581 and 582:
teins and enzymes contributing to r
- Page 583 and 584:
Chitinase gene transcripts began to
- Page 585 and 586:
phenotype for the chitinase gene. A
- Page 587 and 588:
oth a heterologous (tobacco) and in
- Page 590 and 591:
Molecular biology of rice tungro vi
- Page 592 and 593:
Rice tungro bacilliform virus The p
- Page 594 and 595:
Table 1. tRNAs that are (-)-strand
- Page 596 and 597:
Ge X, Gordon D T, Gingery R E, McMu
- Page 598:
Zhang H M, Yang H, Rech E L, Golds
- Page 601 and 602:
the magnitude of the problem, satis
- Page 603 and 604:
ecovered. Rice plants infected with
- Page 605 and 606:
Genetic map Through the work of sev
- Page 607 and 608:
Production of recombinant and trans
- Page 609 and 610:
Studies of host plant resistance ha
- Page 612:
Discussion Session 11: Molecular ge
- Page 616 and 617:
Co-transformation of indica rice vi
- Page 618 and 619:
1. Plasmid constructions used in tr
- Page 620 and 621:
2. Growth of IR54 protoplasts. a =
- Page 622 and 623:
antibiotic-resistant calli per 10 5
- Page 624 and 625:
6. Southern blot of DNA isolated fr
- Page 626 and 627:
another co-transformation study, To
- Page 628 and 629:
Efficient transformation of rice ce
- Page 630 and 631:
hygromycin (50 µg/ml). After furth
- Page 632 and 633:
2. Southern blot analysis of DNA ex
- Page 634 and 635:
Table 4. GUS activity of leaf extra
- Page 636:
Nagy F, Boutry M, Hsu M Y, Wong M,
- Page 639 and 640:
Gene constructs Regenerating rice p
- Page 641 and 642:
callus by histochemical staining us
- Page 643 and 644:
3. Southern blot analysis of DNA ex
- Page 645 and 646:
4. Tissue-specific and constitutive
- Page 647 and 648:
derived from the rice genes could b
- Page 650 and 651:
Evaluation of transformation techni
- Page 652 and 653:
4000 solution. After 30 min of incu
- Page 654 and 655:
Introduction of foreign DNA into dr
- Page 656 and 657:
Jefferson R A, Kavanagh T A, Bevan
- Page 658 and 659:
Transgenic rice plants produced by
- Page 660 and 661:
µg DNA/10 6 protoplasts, and the p
- Page 662 and 663:
3. a) Shoot and root differentiated
- Page 664:
one showed high GUS activity (Fig.
- Page 667 and 668:
Materials and methods Experiments w
- Page 669 and 670:
Progeny test for resistance to hygr
- Page 671 and 672:
3. Southern blot analysis of transg
- Page 673 and 674:
Southern blot data are presented fo
- Page 675 and 676:
Feinberg A P, Vogelstein B (1983) A
- Page 678 and 679:
Polygenic transformation of rice us
- Page 680 and 681:
1. Restriction maps of the plasmids
- Page 682 and 683:
2. Transient GUS expression in root
- Page 684 and 685:
Several somatic embryos germinated
- Page 686 and 687:
Botti C, Vasil I K (1984) Ontogeny
- Page 688:
Yang H, Zhang M, Davey M R, Mulliga
- Page 691 and 692:
integrates into the host genome. Th
- Page 693 and 694:
I am therefore skeptical that bioli
- Page 695 and 696:
has still to be shown. This method,
- Page 697 and 698:
cells and, therefore, all problems
- Page 699 and 700:
Potrykus I, Paszkowski J, Saul M W,
- Page 701 and 702:
A— Datta: Sometimes efficiency is
- Page 704 and 705:
Characterization of a repetitive hy
- Page 706 and 707:
Comparative studies of the structur
- Page 708 and 709:
Evolution of the intergenic spacer
- Page 710 and 711:
Cytoplasmic and nuclear DNA differe
- Page 712 and 713:
Table 2. Correspondence among 6 enz
- Page 714 and 715:
correlations between centimorgans (
- Page 716 and 717:
Table 1. Effect of digestion time o
- Page 718 and 719:
Introduction of exogenous DNA direc
- Page 720 and 721:
Isolation of rice phytochrome genes
- Page 722 and 723:
Restriction fragment length polymor
- Page 724 and 725:
1. Southern blot analysis of size o
- Page 726 and 727:
Accumulation signals of rice endosp
- Page 728 and 729:
Assessment of protoclonal variation
- Page 730 and 731:
Tissue culture and somatic hybridiz
- Page 732 and 733:
References cited Abdullah R, Cockin
- Page 734 and 735:
The isolation of large numbers of s
- Page 736 and 737:
Table 1. Survival rate of callus pr
- Page 738 and 739:
1. Silver-stained 2-dimensional pol
- Page 740 and 741:
Cytohistological observations on pl
- Page 742 and 743:
Production of aneuhaploids (2n=13)
- Page 744 and 745:
Assessment of gametic selection in
- Page 746 and 747:
1. Representation of linkages betwe
- Page 748 and 749:
Salt tolerance in rice callus cultu
- Page 750 and 751:
Regeneration of plants from culture
- Page 752 and 753:
Chromosome studies and multiplicati
- Page 754 and 755:
Current linkage maps in rice T. Kin
- Page 756 and 757:
Table continued. Gene Locus Gene Lo
- Page 758 and 759:
Characterization of rice chromosome
- Page 760 and 761:
Table 1. Numerical data on somatic
- Page 762 and 763:
Table 1. Mutant lines that exhibite
- Page 764 and 765:
1. Segregation for heading time and
- Page 766 and 767:
Table 1. Classification of 19 dwarf
- Page 768 and 769:
Table 1. Top and root characters of
- Page 770 and 771:
The phenotypic diversity of these 2
- Page 772 and 773:
References cited Futsuhara Y, Yamag
- Page 774 and 775:
1. Frequency distribution of degree
- Page 776 and 777:
1. Changes in plant height, root le
- Page 778 and 779:
1. Variation in components among ri
- Page 780 and 781:
These analyses reveal the predomina
- Page 782 and 783:
Table 1. Genomic and continental re
- Page 784 and 785:
Reexamination of genetic control of
- Page 786 and 787:
Relatedness of annual and perennial
- Page 788 and 789:
among prolamin gene copies could be
- Page 790 and 791:
1. Changes in chlorophyll fluoresce
- Page 792 and 793:
Genetic control of chilling injury
- Page 794 and 795:
Table 4. Relationship between genot
- Page 796 and 797:
1. Polypeptide patterns of some M 1
- Page 798 and 799:
Table 1. Near-isogenic lines for BB
- Page 800 and 801:
Table 1. lntrapopulational variatio
- Page 802 and 803:
1. Correlation between reactions to
- Page 804 and 805:
Induced mutations for rice improvem
- Page 806 and 807:
References cited Arasu, N T, Gaul H
- Page 808 and 809:
Table 1. Breeding behavior of selec
- Page 810 and 811:
Mutagenic effects of accelerated ar
- Page 812 and 813:
References cited Cox R, Thacker J,
- Page 814 and 815:
1. Hypothesis for segregatim of sem
- Page 816 and 817:
Combining ability and heterosis for
- Page 818 and 819:
1. Circle of correlations. WGP = we
- Page 820 and 821:
Table 1. Estimates of genetic param
- Page 822 and 823:
Table 1. Components of variation an
- Page 824 and 825:
Table 1. Pollen and spikelet fertil
- Page 826 and 827:
Does hybrid sterility isolate indic
- Page 828 and 829:
Effects of genotype x environment i
- Page 830 and 831:
Variation in chlorophyll proteins b
- Page 832 and 833:
Notes Authors’addresses: N. Watan
- Page 834 and 835:
Table 1. Response of young panicle
- Page 836 and 837:
Differentiation between populations
- Page 838 and 839:
Rice aroma: methods of evaluation a
- Page 840 and 841:
Application of computer image analy
- Page 842 and 843:
Genetic analysis of embryo length i
- Page 844 and 845:
1. Frequency distribution of headin
- Page 846 and 847:
Reliable, simple method for produci
- Page 848 and 849:
2. Rice pollen plants by 1-step cul
- Page 850 and 851:
1. Internode elongation system in w
- Page 852 and 853:
Inheritance of high-density rice gr
- Page 854 and 855:
Table 2. Estimates of gene effects
- Page 856 and 857:
1. Banding patterns of RFLP marker
- Page 858 and 859:
1. Ancient rice seed (AD 800) from
- Page 860:
Hiratsuka J, Shimada H, Whittier R,
- Page 864 and 865:
Overview of the Second Internationa
- Page 866 and 867:
Closing remarks F.A. Bernardo I am
- Page 868:
• The members of the organizing c
- Page 871 and 872:
Report of the meeting on gene symbo
- Page 873 and 874:
Report of the meeting to discuss th
- Page 875 and 876:
References cited Glaszmann J C, de
- Page 877 and 878:
Participants AUSTRALIA P.M. Chay-Pr
- Page 879 and 880:
Xiangmin Wang Research Laboratory o
- Page 881 and 882:
G. Madhava Reddy Department of Gene
- Page 883 and 884:
Mitsugu Horita Hokkaido Green-Bio I
- Page 885 and 886:
K. Nonomura Faculty of Agriculture
- Page 887 and 888:
KOREA, REPUBLIC OF Moon Soo Cho Tae
- Page 889 and 890:
Chairerg Maneephong Faculty of Agri
- Page 891 and 892:
R.L. Rodriguez Department of Geneti
- Page 893 and 894:
Index of varieties and lines 1 001
- Page 895 and 896:
C Calmochi 101 352, 374 Caloro 522
- Page 897 and 898:
IR28211-43-1-1-2 797, 798 IR29692-6
- Page 899 and 900:
P P1231129 [PI 231 129] 444 Pachcha
- Page 901:
X X82 758 Xiang-Dao 80-66 784 Xin-h