

Dispersal and transient dynamics in metapopulations - IEB
Dispersal and transient dynamics in metapopulations - IEB
Dispersal and transient dynamics in metapopulations - IEB

You also want an ePaper? Increase the reach of your titles
YUMPU automatically turns print PDFs into web optimized ePapers that Google loves.
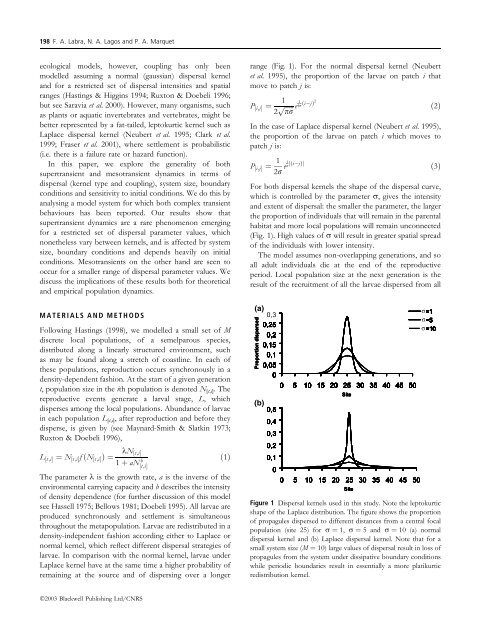
198 F. A. Labra, N. A. Lagos <strong>and</strong> P. A. Marquetecological models, however, coupl<strong>in</strong>g has only beenmodelled assum<strong>in</strong>g a normal (gaussian) dispersal kernel<strong>and</strong> for a restricted set of dispersal <strong>in</strong>tensities <strong>and</strong> spatialranges (Hast<strong>in</strong>gs & Higg<strong>in</strong>s 1994; Ruxton & Doebeli 1996;but see Saravia et al. 2000). However, many organisms, suchas plants or aquatic <strong>in</strong>vertebrates <strong>and</strong> vertebrates, might bebetter represented by a fat-tailed, leptokurtic kernel such asLaplace dispersal kernel (Neubert et al. 1995; Clark et al.1999; Fraser et al. 2001), where settlement is probabilistic(i.e. there is a failure rate or hazard function).In this paper, we explore the generality of bothsuper<strong>transient</strong> <strong>and</strong> meso<strong>transient</strong> <strong>dynamics</strong> <strong>in</strong> terms ofdispersal (kernel type <strong>and</strong> coupl<strong>in</strong>g), system size, boundaryconditions <strong>and</strong> sensitivity to <strong>in</strong>itial conditions. We do this byanalys<strong>in</strong>g a model system for which both complex <strong>transient</strong>behaviours has been reported. Our results show thatsuper<strong>transient</strong> <strong>dynamics</strong> are a rare phenomenon emerg<strong>in</strong>gfor a restricted set of dispersal parameter values, whichnonetheless vary between kernels, <strong>and</strong> is affected by systemsize, boundary conditions <strong>and</strong> depends heavily on <strong>in</strong>itialconditions. Meso<strong>transient</strong>s on the other h<strong>and</strong> are seen tooccur for a smaller range of dispersal parameter values. Wediscuss the implications of these results both for theoretical<strong>and</strong> empirical population <strong>dynamics</strong>.MATERIALS AND METHODSFollow<strong>in</strong>g Hast<strong>in</strong>gs (1998), we modelled a small set of Mdiscrete local populations, of a semelparous species,distributed along a l<strong>in</strong>early structured environment, suchas may be found along a stretch of coastl<strong>in</strong>e. In each ofthese populations, reproduction occurs synchronously <strong>in</strong> adensity-dependent fashion. At the start of a given generationt, population size <strong>in</strong> the ith population is denoted N [t,i] . Thereproductive events generate a larval stage, L, whichdisperses among the local populations. Abundance of larvae<strong>in</strong> each population L [t,i] , after reproduction <strong>and</strong> before theydisperse, is given by (see Maynard-Smith & Slatk<strong>in</strong> 1973;Ruxton & Doebeli 1996),L ½t;iŠ ¼ N ½t;iŠ f ðN ½t;iŠ Þ¼kN ½t;iŠ1 þ aN½t;iŠbð1ÞThe parameter k is the growth rate, a is the <strong>in</strong>verse of theenvironmental carry<strong>in</strong>g capacity <strong>and</strong> b describes the <strong>in</strong>tensityof density dependence (for further discussion of this modelsee Hassell 1975; Bellows 1981; Doebeli 1995). All larvae areproduced synchronously <strong>and</strong> settlement is simultaneousthroughout the metapopulation. Larvae are redistributed <strong>in</strong> adensity-<strong>in</strong>dependent fashion accord<strong>in</strong>g either to Laplace ornormal kernel, which reflect different dispersal strategies oflarvae. In comparison with the normal kernel, larvae underLaplace kernel have at the same time a higher probability ofrema<strong>in</strong><strong>in</strong>g at the source <strong>and</strong> of dispers<strong>in</strong>g over a longerrange (Fig. 1). For the normal dispersal kernel (Neubertet al. 1995), the proportion of the larvae on patch i thatmove to patch j is:P ½i;jŠ ¼ p 12 ffiffiffiffiffi e4r 1 ði jÞ2 ð2ÞprIn the case of Laplace dispersal kernel (Neubert et al. 1995),the proportion of the larvae on patch i which moves topatch j is:P ½i;jŠ ¼ 12r e1 r jði jÞj ð3ÞFor both dispersal kernels the shape of the dispersal curve,which is controlled by the parameter r, gives the <strong>in</strong>tensity<strong>and</strong> extent of dispersal: the smaller the parameter, the largerthe proportion of <strong>in</strong>dividuals that will rema<strong>in</strong> <strong>in</strong> the parentalhabitat <strong>and</strong> more local populations will rema<strong>in</strong> unconnected(Fig. 1). High values of r will result <strong>in</strong> greater spatial spreadof the <strong>in</strong>dividuals with lower <strong>in</strong>tensity.The model assumes non-overlapp<strong>in</strong>g generations, <strong>and</strong> soall adult <strong>in</strong>dividuals die at the end of the reproductiveperiod. Local population size at the next generation is theresult of the recruitment of all the larvae dispersed from allFigure 1 <strong>Dispersal</strong> kernels used <strong>in</strong> this study. Note the leptokurticshape of the Laplace distribution. The figure shows the proportionof propagules dispersed to different distances from a central focalpopulation (site 25) for r ¼ 1, r ¼ 5 <strong>and</strong> r ¼ 10 (a) normaldispersal kernel <strong>and</strong> (b) Laplace dispersal kernel. Note that for asmall system size (M ¼ 10) large values of dispersal result <strong>in</strong> loss ofpropagules from the system under dissipative boundary conditionswhile periodic boundaries result <strong>in</strong> essentially a more platikurticredistribution kernel.σσσÓ2003 Blackwell Publish<strong>in</strong>g Ltd/CNRS













