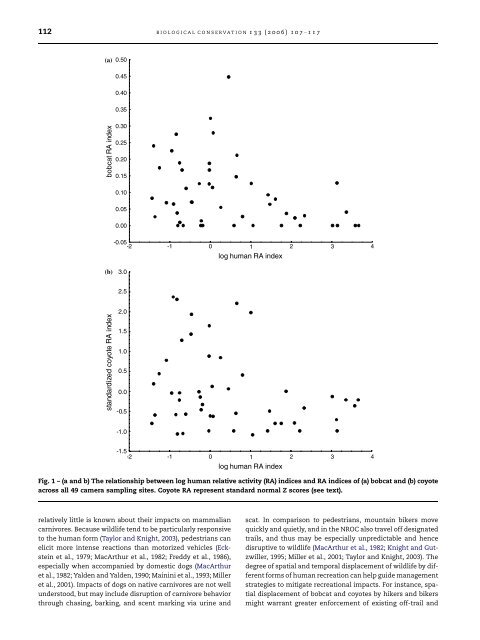
112 BIOLOGICAL CONSERVATION 133 (2006) 107– 117(a) 0.500.450.400.35bobcat RA <strong>in</strong>dex0.300.250.200.150.100.050.00-0.05-2 -1 0 1 2 34log hum<strong>an</strong> RA <strong>in</strong>dex(b)3.02.5st<strong><strong>an</strong>d</strong>ardized coyote RA <strong>in</strong>dex2.01.51.00.50.0-0.5-1.0-1.5-2 -1 0 1 2 34log hum<strong>an</strong> RA <strong>in</strong>dexFig. 1 – (a <strong><strong>an</strong>d</strong> b) The relationship between log hum<strong>an</strong> relative <strong>activity</strong> (RA) <strong>in</strong>dices <strong><strong>an</strong>d</strong> RA <strong>in</strong>dices of (a) bobcat <strong><strong>an</strong>d</strong> (b) coyoteacross all 49 camera sampl<strong>in</strong>g sites. Coyote RA represent st<strong><strong>an</strong>d</strong>ard normal Z scores (see text).relatively little is known about their impacts on <strong>mammal</strong>i<strong>an</strong>carnivores. Because wildlife tend to be particularly responsiveto the hum<strong>an</strong> form (Taylor <strong><strong>an</strong>d</strong> Knight, 2003), pedestri<strong>an</strong>s c<strong>an</strong>elicit more <strong>in</strong>tense reactions th<strong>an</strong> motorized vehicles (Eckste<strong>in</strong>et al., 1979; MacArthur et al., 1982; Freddy et al., 1986),especially when accomp<strong>an</strong>ied by domestic dogs (MacArthuret al., 1982; Yalden <strong><strong>an</strong>d</strong> Yalden, 1990; Ma<strong>in</strong><strong>in</strong>i et al., 1993; Milleret al., 2001). Impacts of dogs on native carnivores are not wellunderstood, but may <strong>in</strong>clude disruption of carnivore behaviorthrough chas<strong>in</strong>g, bark<strong>in</strong>g, <strong><strong>an</strong>d</strong> scent mark<strong>in</strong>g via ur<strong>in</strong>e <strong><strong>an</strong>d</strong>scat. In comparison to pedestri<strong>an</strong>s, mounta<strong>in</strong> bikers movequickly <strong><strong>an</strong>d</strong> quietly, <strong><strong>an</strong>d</strong> <strong>in</strong> the NROC also travel off designatedtrails, <strong><strong>an</strong>d</strong> thus may be especially unpredictable <strong><strong>an</strong>d</strong> hencedisruptive to wildlife (MacArthur et al., 1982; Knight <strong><strong>an</strong>d</strong> Gutzwiller,1995; Miller et al., 2001; Taylor <strong><strong>an</strong>d</strong> Knight, 2003). Thedegree of spatial <strong><strong>an</strong>d</strong> temporal displacement of wildlife by differentforms of hum<strong>an</strong> recreation c<strong>an</strong> help guide m<strong>an</strong>agementstrategies to mitigate recreational impacts. For <strong>in</strong>st<strong>an</strong>ce, spatialdisplacement of bobcat <strong><strong>an</strong>d</strong> coyotes by hikers <strong><strong>an</strong>d</strong> bikersmight warr<strong>an</strong>t greater enforcement of exist<strong>in</strong>g off-trail <strong><strong>an</strong>d</strong>
BIOLOGICAL CONSERVATION 133 (2006) 107– 117 113Table 3 – Relationship between wildlife relative <strong>activity</strong> (RA) <strong><strong>an</strong>d</strong> percent daytime <strong>activity</strong> (PDA) <strong><strong>an</strong>d</strong> hum<strong>an</strong> recreationalcategoriesBobcat Coyote Mule deerCoefficient r P Coefficient r P Coefficient r PRA *Overall hum<strong>an</strong> 0.023 0.33 0.021 0.193 0.29 0.047 0.007 0.04 0.775Hiker 0.038 0.34 0.016 0.371 0.34 0.017 0.012 0.05 0.756Biker 0.045 0.36 0.010 0.307 0.26 0.076 0.051 0.18 0.229Vehicles 0.039 0.11 0.433 0.555 0.17 0.250 0.008 0.01 0.947Equestri<strong>an</strong> 0.040 0.07 0.629 0.240 0.04 0.764 0.221 0.17 0.252Dog 0.028 0.07 0.633 0.318 0.08 0.578 0.128 0.14 0.352PDA **Overall hum<strong>an</strong> 0.369 0.50 0.013 0.127 0.22 0.193 0.042 0.13 0.480Hiker 0.618 0.54 0.006 0.243 0.27 0.108 0.108 0.18 0.329Biker 2.367 0.45 0.028 0.229 0.22 0.193 0.021 0.04 0.819Vehicles 0.100 0.03 0.882 0.407 0.16 0.366 0.087 0.05 0.806Equestri<strong>an</strong> 0.485 0.10 0.654 0.354 0.07 0.671 0.749 0.25 0.173Dog 1.574 0.51 0.012 1.078 0.29 0.081 0.129 0.04 0.849*All n = 49.**All n = 24 for bobcats; all n = 36 for coyotes; all n = 32 for mule deer.Table 4 – AIC results for wildlife percent daytime <strong>activity</strong> (PDA)Species Model log(Lhood) K D i AIC c W iBobcat a Log Hum<strong>an</strong> 10.168 3 0 0.642(Log Hum<strong>an</strong>) 2 8.633 3 3.070 0.138Log Hum<strong>an</strong> + (Log Hum<strong>an</strong>) 2 10.370 4 3.406 0.117Null 6.715 2 3.650 0.103Coyote a (Log Hum<strong>an</strong>) 2 9.812 3 0 0.423Null 8.067 2 0.714 0.296Log Hum<strong>an</strong> 8.979 3 1.667 0.184Log Hum<strong>an</strong> + (Log Hum<strong>an</strong>) 2 9.885 4 2.935 0.097Mule deer Null 24.958 2 0 0.614Log Hum<strong>an</strong> 25.228 3 2.347 0.190(Log Hum<strong>an</strong>) 2 25.026 3 2.750 0.155Log Hum<strong>an</strong> + (Log Hum<strong>an</strong>) 2 25.306 4 5.439 0.040Four models were exam<strong>in</strong>ed: (1) Null, <strong>in</strong> which hum<strong>an</strong>s were excluded from <strong>an</strong>alysis; (2) Log Hum<strong>an</strong>, represent<strong>in</strong>g a l<strong>in</strong>ear relationshipbetween hum<strong>an</strong> relative <strong>activity</strong> <strong>in</strong>dices <strong><strong>an</strong>d</strong> wildlife percent daytime <strong>activity</strong>; (3) (Log Hum<strong>an</strong>) 2 , which represents a non-l<strong>in</strong>ear relationshipbetween hum<strong>an</strong> <strong>in</strong>dices <strong><strong>an</strong>d</strong> wildlife percent daytime <strong>activity</strong>; <strong><strong>an</strong>d</strong> (4) Log Hum<strong>an</strong> + (Log Hum<strong>an</strong>) 2 , a non-l<strong>in</strong>ear 2nd order polynomial model.Sites with five images or more were <strong>in</strong>cluded for species PDA <strong>an</strong>alyses (see text).a St<strong><strong>an</strong>d</strong>ard normal (Z) scores.trespass<strong>in</strong>g regulations, or even sett<strong>in</strong>g aside new areas thatrestrict recreation. Alternatively, temporal displacement bydomestic dogs may suggest limit<strong>in</strong>g the hours <strong>in</strong> which a <strong>reserve</strong>is open to dog walk<strong>in</strong>g.Our f<strong>in</strong>d<strong>in</strong>gs that bobcats appeared more responsive to hum<strong><strong>an</strong>d</strong>isturb<strong>an</strong>ces th<strong>an</strong> did coyotes are consistent with priorstudies <strong>in</strong> the region. For <strong>in</strong>st<strong>an</strong>ce, Crooks (2002) exam<strong>in</strong>edthe effects of habitat fragmentation on carnivores <strong>in</strong> coastalsouthern California <strong><strong>an</strong>d</strong> concluded that bobcats were moresensitive to l<strong><strong>an</strong>d</strong>scape variables such as fragment size <strong><strong>an</strong>d</strong>isolation th<strong>an</strong> coyotes. Tigas et al. (2002) studied the responsesof radio-collared bobcats <strong><strong>an</strong>d</strong> coyotes to fragmentation<strong><strong>an</strong>d</strong> corridors <strong>in</strong> <strong>an</strong> urb<strong>an</strong> area northwest of LosAngeles <strong><strong>an</strong>d</strong> suggested that although both species partiallyadjusted to habitat fragmentation through spatial <strong><strong>an</strong>d</strong> temporalavoid<strong>an</strong>ce, coyotes tended to be relatively more toler<strong>an</strong>t ofhum<strong>an</strong> development. In the same system, Riley et al. (2003)found that some bobcats <strong>in</strong>corporated partially developedl<strong><strong>an</strong>d</strong>s <strong>in</strong>to their home r<strong>an</strong>ges, but to a lesser extent th<strong>an</strong> coyotes.Consistent with our f<strong>in</strong>d<strong>in</strong>gs, Tigas et al. (2002) <strong><strong>an</strong>d</strong> Rileyet al. (2003) also found that bobcats <strong><strong>an</strong>d</strong> coyotes tended toshift towards nocturnal use of more fragmented, developedareas; studies <strong>in</strong> other systems have also suggested that coyotes<strong>in</strong> urb<strong>an</strong> areas shift their <strong>activity</strong> to times when hum<strong>an</strong>sare less active (Andelt <strong><strong>an</strong>d</strong> Mah<strong>an</strong>, 1980; Qu<strong>in</strong>n, 1997a; Gr<strong>in</strong>der<strong><strong>an</strong>d</strong> Krausm<strong>an</strong>, 2001; McClennen et al., 2001).Research on urb<strong>an</strong> deer typically has focused on topicssuch as space use (Kilpatrick <strong><strong>an</strong>d</strong> Spohr, 2000), movementpatterns (Grund et al., 2002), hum<strong>an</strong> conflicts (McCulloughet al., 1997; Wagner et al., 1997), <strong><strong>an</strong>d</strong> responses to hunt<strong>in</strong>gpressures (Kilpatrick <strong><strong>an</strong>d</strong> Lima, 1999), but fewer studies have<strong>in</strong>vestigated possible recreational impacts on urb<strong>an</strong> deer