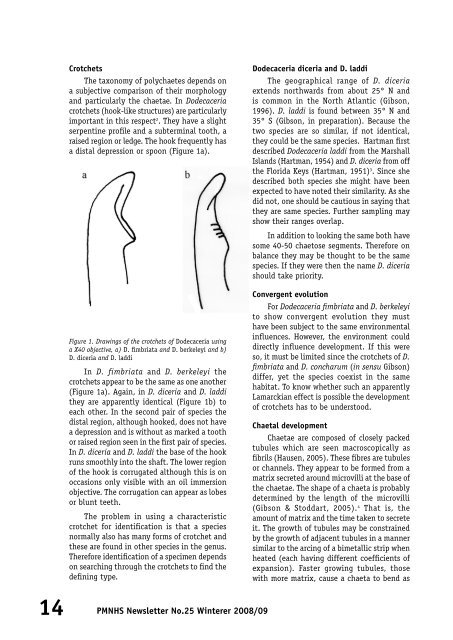
CrotchetsThe taxonomy of polychaetes depends ona subjective comparison of their morphologyand particularly the chaetae. In Dodecaceriacrotchets (hook-like structures) are particularlyimportant in this respect 2 . They have a slightserpentine profile and a subterminal tooth, araised region or ledge. The hook frequently hasa distal depression or spoon (Figure 1a).Dodecaceria diceria and D. laddiThe geographical range of D. diceriaextends northwards from about <strong>25</strong>° N andis common in the North Atlantic (Gibson,1996). D. laddi is found between 35° N and35° S (Gibson, in preparation). Because thetwo species are so similar, if not identical,they could be the same species. Hartman firstdescribed Dodecaceria laddi from the MarshallIslands (Hartman, 1954) and D. diceria from offthe Florida Keys (Hartman, 1951) 3 . Since shedescribed both species she might have beenexpected to have noted their similarity. As shedid not, one should be cautious in saying thatthey are same species. Further sampling mayshow their ranges overlap.In addition to looking the same both havesome 40-50 chaetose segments. Therefore onbalance they may be thought to be the samespecies. If they were then the name D. diceriashould take priority.Figure 1. Drawings of the crotchets of Dodecaceria usinga X40 objective, a) D. fimbriata and D. berkeleyi and b)D. diceria and D. laddiIn D. fimbriata and D. berkeleyi thecrotchets appear to be the same as one another(Figure 1a). Again, in D. diceria and D. laddithey are apparently identical (Figure 1b) toeach other. In the second pair of species thedistal region, although hooked, does not havea depression and is without as marked a toothor raised region seen in the first pair of species.In D. diceria and D. laddi the base of the hookruns smoothly into the shaft. The lower regionof the hook is corrugated although this is onoccasions only visible with an oil immersionobjective. The corrugation can appear as lobesor blunt teeth.The problem in using a characteristiccrotchet for identification is that a speciesnormally also has many forms of crotchet andthese are found in other species in the genus.Therefore identification of a specimen dependson searching through the crotchets to find thedefining type.Convergent evolutionFor Dodecaceria fimbriata and D. berkeleyito show convergent evolution they musthave been subject to the same environmentalinfluences. However, the environment coulddirectly influence development. If this wereso, it must be limited since the crotchets of D.fimbriata and D. concharum (in sensu Gibson)differ, yet the species coexist in the samehabitat. To know whether such an apparentlyLamarckian effect is possible the developmentof crotchets has to be understood.Chaetal developmentChaetae are composed of closely packedtubules which are seen macroscopically asfibrils (Hausen, 2005). These fibres are tubulesor channels. They appear to be formed from amatrix secreted around microvilli at the base ofthe chaetae. The shape of a chaeta is probablydetermined by the length of the microvilli(Gibson & Stoddart, 2005). 4 That is, theamount of matrix and the time taken to secreteit. The growth of tubules may be constrainedby the growth of adjacent tubules in a mannersimilar to the arcing of a bimetallic strip whenheated (each having different coefficients ofexpansion). Faster growing tubules, thosewith more matrix, cause a chaeta to bend as14PMNHS <strong>Newsletter</strong> No.<strong>25</strong> <strong>Winter</strong>er <strong>2008</strong>/09
it grows 4 . The shape of chaetae will thereforedepend on the characteristics of the microvilli.A wave of secretory activity appears to passacross the microvilli. The nature of thewave, its mathematical characteristic, wouldmodulate growth.The polychaete crotchet and uncinusappear to have the same basic shape. Onecan be graphically converted to the otherusing a D’Arcy Thompson transformation(Gibson, 2002). That is, one type can be“morphed”, evolutionarily speaking, into theother by differential growth. This principleprobably applies to all types of chaetae. Theshape of chaetae, then, might, as mentionedabove, conform to a mathematic function(Gibson, Robson & Armitage, 1999). Thisfunction may to some degree be influencedby the environment as in developmentalcanalization, first postulated by Waddington(Calow, 1983).Chaetae grow sequentially and their shapevaries with their age. The form of the crotchetsof Dodecaceria varies within the fascicle of aparapoda, between neuropodia and notopodiaand along the body (Caullery & Mesnil, 1898).This, as noted, is the same for other species.In Arenicola, for example, the rostral teeth ofthe crotches become smaller and can disappear(Ashworth, 1912). Interaction betweenenvironment and gene control may thereforechange with age.Polychaete evolutionPolychaetes are best understood as anad hoc group. That is, their evolution is notclearly monophyletic with one group givingrise to another 5 . The structure of their chaetae,for example, is best seen in terms of functionrather than phylogeny. Dales based hisclassification on feeding structures (1963).Rouse & Pleijel (2001) were equivocal overmorphological evidence for a monophyleticorigin. Polychaetes could have polyphyleticorigin if they had a Precambrian planktonicorigin (Gibson, in preparation). Speciationcould have occurred within the planktonand species dispersed globally in surfacewaters. This might account for the apparentbipolar distributions of D. fimbriata and D.berkeleyi 6 . Planktonic dispersal obviates theneed to postulate continual drift. However,this origin supposes evolutionary stasis (orthat exactly the same changes have occurredin both species).NomenclatureThe distribution and naming of Dodecaceriafimbriata/D. berkeleyi and D. diceria/ D. laddihighlights a weakness in classical taxonomy.The use of priority in naming species has ledto confusion (Gibson & Heppell, 1995; Heppell& Gibson, 1995). The problem is not solved bycompelling authors to follow prescribed codesformulated by bodies such as the InternationalCommission on Zoological Nomenclature. Theseprocedures fail because they stultify taxonomy(Heppell, 1991). Systems must be flexibleenough to allow for changes in approach.Because morphological taxonomy dependson judgement it lacks rigor. This is particularlytrue for Dodecaceria where the species aremorphologically so similar. Computer imageanalysis of the shape of chaetae, a moreobjective method, was used in an attempt toseparate species in this genus (Gibson, Robson& Armitage, 1999; Gibson & Stoddart, 2005).Because the reproduction of the species ofDodecaceria is so varied another approach tothe taxonomy was to use differences in theirreproductive biology (Gibson, 1978). Othermethods that might help in separating speciescould be the use of ontology and ecology. Barcoding (Heppell, 1991) and DNA finger printingmight be effectively used in naming species.SummaryDodecaceria fimbriata and D. berkeleyi havea bipolar distribution even though they appearto be the same species. Continental drift canbe used to account for their distribution. Analternative explanation is that the ancestorsto polychaetes evolved in the Precambrianplankton and were distributed within surfacewaters. There is also the possibility ofconvergent evolution due to similarities inhabitats. However, the environment mayalso have a direct effect on chaetal structurethrough developmental canalization.Dodecaceria diceria and D. laddi arealso morphologically very similar. Theirdistributions although different may on furtherPMNHS <strong>Newsletter</strong> No.<strong>25</strong> <strong>Winter</strong> <strong>2008</strong>/0915