170 Harfsame (Dowling et al., 1992). Endocytosis and intracellular multiplication of L.pneumophila in amoeba and in a monocyte-like cell line (U937) was investigated byusing cytocholasin D, and methylamine (King et al., 1991). Cytocholasin D is aninhibitor of actin filament polymerization involved in plasma membrane invaginationduring phagocytosis. Methylamine is an inhibitor of transglutaminase, a plasmamembrane enzyme involved in aggregation of ligand-receptor complexes required forreceptor-mediated pinocytosis in fibroblasts and macrophages. Methylamine inhibitedreplication of L. pneumophila in amoeba. Intra amoebal multiplication of L.pneumophila was not inhibited by cytocholasin D. Thus it is microfilamentindependent,in contrast to macrophages, and according to Fields (1993) a newterminology is needed to describe bacterial uptake by a microfilament-independentprocess.The efficiency of consumption is different, depending on protozoan type andgrowth state, temperature, bacterial abundance, and size and nature of bacterial prey(Gonzalez et al., 1990). Many of the bacteria are digested, especially enterobacteriawhich are used as nutriment. Gram positive bacteria seem to be digested slower,owing to their thick cell wall.Escaping digestionSome bacteria, as shown for Gram negatives by Cavalier-Smith and Lee (1985),may avoid digestion by liberation out of the phagosome before lysosomal fusion orresist the cellular mechanisms of digestion in phagosomes and escape into the hostcell cytoplasm where they multiply. (Hall and Voelz, 1985; Reisser et al., 1985).According to Choi et al., 1992; and Jeon, 1993, non-fusible vesicles such assymbiosomes include symbiont derived lipopolysaccharides which prevent lysosomesfrom fusing with them. Fields (1993), investigating a model of L. pneumophilaendocytosis by Hartmannella vermiformis, described a fusion of the endosómecontaining the internalized Legionella with the host cell's endoplasmic reticulum. Theendoplasmic reticulum becomes the site of bacterial multiplication as it is rich innutrients and would prevent exposure to lysosomes.Escaping digestion can evolve in different types of interactions between amoebaand bacteria :Endosymbionts are permanent harboured bacteria and cannot be cultured so far.Some are described as toxic killer symbionts (Görtz, 1993). An obligateintracellular bacterial parasite, decribed as Sarcobium lyticum, causing fatalinfection in Acanthamoeba castellana, shows a close relationship to anintracellular Legionella (Drozanski 1991). According to Fritsche et al, (1993),
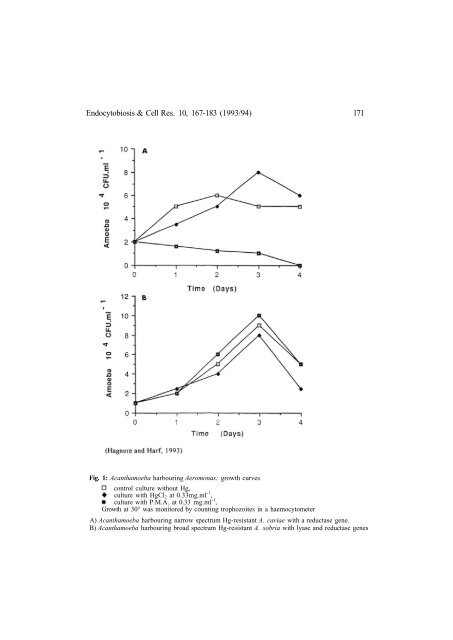
Endocytobiosis & Cell Res. 10, 167-183 (1993/94) 171Fig. 1: Acanthamoeba harbouring Aeromonas: growth curvescontrol culture without Hg,culture with HgCl 2 at 0.33mg.ml -1 ,culture with P.M.A. at 0.33 mg.ml -1 ,Growth at 30° was monitored by counting trophozoites in a haemocytometerA) Acanthamoeba harbouring narrow spectrum Hg-resistant A. caviae with a reductase gene.B) Acanthamoeba harbouring broad spectrum Hg-resistant A. sobria with lyase and reductase genes