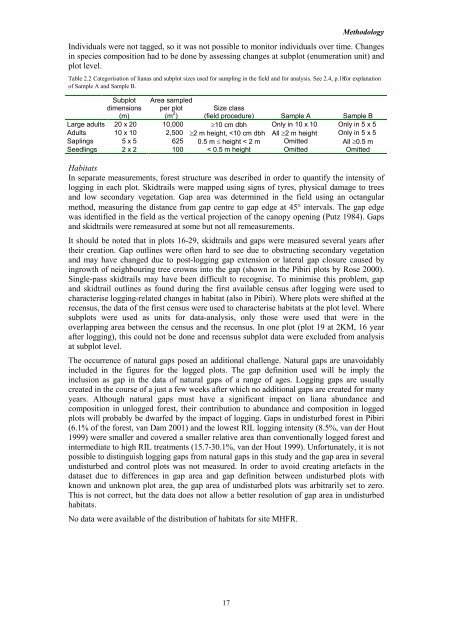
Methodology Individuals were not tagged, so it was not possible to m<strong>on</strong>itor <strong>in</strong>dividuals over time. Changes <strong>in</strong> species compositi<strong>on</strong> had to be d<strong>on</strong>e by assess<strong>in</strong>g changes at subplot (enumerati<strong>on</strong> unit) <strong>and</strong> plot level. Table 2.2 Categorisati<strong>on</strong> of <strong>liana</strong>s <strong>and</strong> subplot sizes used for sampl<strong>in</strong>g <strong>in</strong> the field <strong>and</strong> for analysis. See 2.4, p.18for explanati<strong>on</strong> of Sample A <strong>and</strong> Sample B. Subplot dimensi<strong>on</strong>s (m) Area sampled per plot (m 2 ) Size class (field procedure) Sample A Sample B Large adults 20 x 20 10,000 ≥10 cm dbh Only <strong>in</strong> 10 x 10 Only <strong>in</strong> 5 x 5 Adults 10 x 10 2,500 ≥2 m height,
<str<strong>on</strong>g>Logg<strong>in</strong>g</str<strong>on</strong>g> <str<strong>on</strong>g>effects</str<strong>on</strong>g> <strong>on</strong> <strong>liana</strong> <strong>diversity</strong> <strong>and</strong> <strong>abundance</strong> <strong>in</strong> <strong>Central</strong> <strong>Guyana</strong> 2.4 ANALYSIS 2.4.1 Data preparati<strong>on</strong> Data storage The data were <strong>in</strong>itially stored <strong>in</strong> separate databases for plots (plot database, c<strong>on</strong>ta<strong>in</strong><strong>in</strong>g the <strong>in</strong>formati<strong>on</strong> from Table 2.1), species (species database, c<strong>on</strong>ta<strong>in</strong><strong>in</strong>g <strong>in</strong>formati<strong>on</strong> about names), <strong>in</strong>dividuals (<strong>in</strong>dividuals database, c<strong>on</strong>ta<strong>in</strong><strong>in</strong>g the <strong>in</strong>formati<strong>on</strong> obta<strong>in</strong>ed from the field measurements, see “measurements”) <strong>and</strong> habitats (habitat database, c<strong>on</strong>ta<strong>in</strong><strong>in</strong>g identifiers <strong>and</strong> coord<strong>in</strong>ates of the polyg<strong>on</strong>s describ<strong>in</strong>g skidtrails <strong>and</strong> gaps). Subplots were used as identifiers to relate censuses from different years <strong>in</strong> the Pibiri plots, while this was not possible <strong>in</strong> the rema<strong>in</strong><strong>in</strong>g plots due to shifts <strong>in</strong> plot locati<strong>on</strong>s. Informati<strong>on</strong> <strong>on</strong> <strong>in</strong>dividuals measured <strong>in</strong> each census could not be related to each other, as they were not <strong>in</strong>dividually tagged. The <strong>in</strong>dividuals database <strong>and</strong> the habitat database were spatially l<strong>in</strong>ked us<strong>in</strong>g the PCraster GIS package. Before this was d<strong>on</strong>e, habitats were mapped <strong>and</strong> 5 m border z<strong>on</strong>es were def<strong>in</strong>ed <strong>in</strong>side <strong>and</strong> outside gaps. This was d<strong>on</strong>e for two reas<strong>on</strong>s: the methodological uncerta<strong>in</strong>ty associated with determ<strong>in</strong><strong>in</strong>g gap edges <strong>in</strong> the field, <strong>and</strong> to cater for possible ecological edge <str<strong>on</strong>g>effects</str<strong>on</strong>g> of gaps <strong>in</strong>to forest surround<strong>in</strong>g gaps <strong>and</strong> of surround<strong>in</strong>g forest <strong>in</strong>to gaps. Possible ecological edge <str<strong>on</strong>g>effects</str<strong>on</strong>g> <strong>in</strong>clude the micro-climatological <str<strong>on</strong>g>effects</str<strong>on</strong>g> of shad<strong>in</strong>g or <strong>in</strong>solati<strong>on</strong> <strong>in</strong>to the gap <strong>and</strong> forest, respectively (van Dam 2001), the different probability of propagule arrival close to edges as compared to gap centres <strong>and</strong> other differences such as litterfall (van Dam 2001). In the data analysis, when gap size is described this was based <strong>on</strong> the gap def<strong>in</strong>iti<strong>on</strong> as used <strong>in</strong> the field. However, when subplots were assigned to habitat-categories (gap/n<strong>on</strong>-gap), the 5 m z<strong>on</strong>e <strong>in</strong>to the forest was <strong>in</strong>cluded <strong>in</strong> the gap def<strong>in</strong>iti<strong>on</strong>, for the reas<strong>on</strong>s described above. In the database, <strong>in</strong>dividuals can be l<strong>in</strong>ked to habitats <strong>in</strong> two ways. Either the coord<strong>in</strong>ates of each <strong>in</strong>dividual <strong>and</strong> the coord<strong>in</strong>ates of the various habitat categories can be l<strong>in</strong>ked to give <strong>in</strong>dividual habitat assignments, or subplots can be assigned a code <strong>on</strong> the basis of the most extensive habitat. There were two habitat variables cod<strong>in</strong>g for ground-level disturbance (<strong>on</strong> or off a skidtrail) <strong>and</strong> for canopy-level disturbance (locati<strong>on</strong> <strong>in</strong> forest, forest edge, gap edge <strong>and</strong> gap <strong>in</strong>terior; both edge z<strong>on</strong>es are 5 m wide). Habitat categories were merged for analysis if necessary (e.g. gaps were def<strong>in</strong>ed as forest edge + gap edge + gap <strong>in</strong>terior for some analysis). For analysis, given the uncerta<strong>in</strong>ty associated with determ<strong>in</strong><strong>in</strong>g coord<strong>in</strong>ates <strong>in</strong> the field, preference was given to the sec<strong>on</strong>d method of assign<strong>in</strong>g habitat codes to <strong>in</strong>dividuals, i.e. by means of uniform codes at the subplot level. All subplots were classified <strong>in</strong> <strong>on</strong>e of four habitat categories: skidded gap (canopy <strong>and</strong> soil disturbance), skidtrail (soil disturbance), gap (canopy disturbance) or forest <strong>in</strong>terior (no disturbance). Selecti<strong>on</strong> of data – Sample A <strong>and</strong> Sample B In the <strong>in</strong>dividuals database, <strong>in</strong>dividuals were selected for analysis based <strong>on</strong> a number of criteria: • Seedl<strong>in</strong>gs as enumerated <strong>in</strong> the 2 x 2 m subplots were reta<strong>in</strong>ed for descriptive purposes of populati<strong>on</strong> size distributi<strong>on</strong> <strong>on</strong>ly. They were not <strong>in</strong>cluded <strong>in</strong> the analysis of compositi<strong>on</strong>al change <strong>and</strong> species resp<strong>on</strong>ses to logg<strong>in</strong>g. • The size criteri<strong>on</strong> for large adult <strong>liana</strong>s (dbh>10 cm) made that few <strong>in</strong>dividuals were measured at the level of the 20 x 20 m subplot (Table 2.2). Before the logg<strong>in</strong>g, <strong>on</strong>ly 1.9% of the total measured populati<strong>on</strong> <strong>in</strong> Pibiri c<strong>on</strong>sisted of <strong>in</strong>dividuals of this size (average 0.8 per subplot or 21 per ha, range 7-34). Therefore, this spatial level was <strong>on</strong>ly used for descriptive purposes but not most analysis. Instead, two samples were created at different spatial levels of analysis (Table 2.2). Sample A <strong>in</strong>cludes all <strong>liana</strong>s > 2m sampled with<strong>in</strong> the 10 x 10 m subplots. Sample B <strong>in</strong>cludes all <strong>liana</strong>s ≥ 0.5 m height with<strong>in</strong> the 5 x 5 m subplots. All <strong>liana</strong>s satisfy<strong>in</strong>g the criteria for <strong>in</strong>clusi<strong>on</strong> <strong>in</strong> 18