

Resilience in aquatic ecosystems - hysteresis, homeostasis, and ...
Resilience in aquatic ecosystems - hysteresis, homeostasis, and ...
Resilience in aquatic ecosystems - hysteresis, homeostasis, and ...

You also want an ePaper? Increase the reach of your titles
YUMPU automatically turns print PDFs into web optimized ePapers that Google loves.
8<br />
Reynolds/ Aquatic Ecosystem Health <strong>and</strong> Management 5 (2002) 3–17<br />
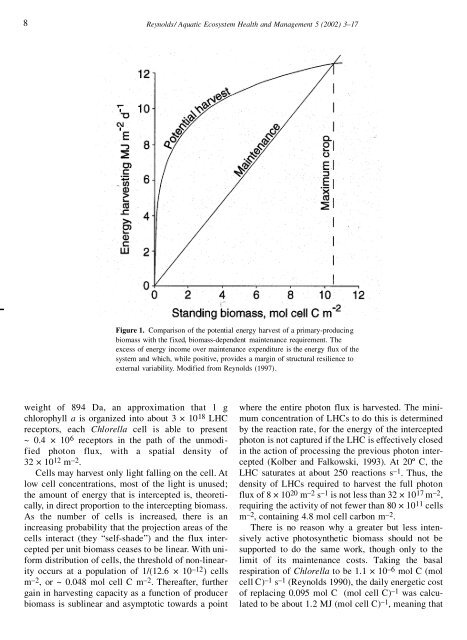
Figure 1. Comparison of the potential energy harvest of a primary-produc<strong>in</strong>g<br />
biomass with the fixed, biomass-dependent ma<strong>in</strong>tenance requirement. The<br />
excess of energy <strong>in</strong>come over ma<strong>in</strong>tenance expenditure is the energy flux of the<br />
system <strong>and</strong> which, while positive, provides a marg<strong>in</strong> of structural resilience to<br />
external variability. Modified from Reynolds (1997).<br />
weight of 894 Da, an approximation that 1 g<br />
chlorophyll a is organized <strong>in</strong>to about 3 × 10 18 LHC<br />
receptors, each Chlorella cell is able to present<br />
~ 0.4 × 10 6 receptors <strong>in</strong> the path of the unmodified<br />
photon flux, with a spatial density of<br />
32 × 10 12 m –2 .<br />
Cells may harvest only light fall<strong>in</strong>g on the cell. At<br />
low cell concentrations, most of the light is unused;<br />
the amount of energy that is <strong>in</strong>tercepted is, theoretically,<br />
<strong>in</strong> direct proportion to the <strong>in</strong>tercept<strong>in</strong>g biomass.<br />
As the number of cells is <strong>in</strong>creased, there is an<br />
<strong>in</strong>creas<strong>in</strong>g probability that the projection areas of the<br />
cells <strong>in</strong>teract (they “self-shade”) <strong>and</strong> the flux <strong>in</strong>tercepted<br />
per unit biomass ceases to be l<strong>in</strong>ear. With uniform<br />
distribution of cells, the threshold of non-l<strong>in</strong>earity<br />
occurs at a population of 1/(12.6 × 10 –12 ) cells<br />
m –2 , or ~ 0.048 mol cell C m –2 . Thereafter, further<br />
ga<strong>in</strong> <strong>in</strong> harvest<strong>in</strong>g capacity as a function of producer<br />
biomass is subl<strong>in</strong>ear <strong>and</strong> asymptotic towards a po<strong>in</strong>t<br />
where the entire photon flux is harvested. The m<strong>in</strong>imum<br />
concentration of LHCs to do this is determ<strong>in</strong>ed<br />
by the reaction rate, for the energy of the <strong>in</strong>tercepted<br />
photon is not captured if the LHC is effectively closed<br />
<strong>in</strong> the action of process<strong>in</strong>g the previous photon <strong>in</strong>tercepted<br />
(Kolber <strong>and</strong> Falkowski, 1993). At 20º C, the<br />
LHC saturates at about 250 reactions s –1 . Thus, the<br />
density of LHCs required to harvest the full photon<br />
flux of 8 × 10 20 m –2 s –1 is not less than 32 × 10 17 m –2 ,<br />
requir<strong>in</strong>g the activity of not fewer than 80 × 10 11 cells<br />
m –2 , conta<strong>in</strong><strong>in</strong>g 4.8 mol cell carbon m –2 .<br />
There is no reason why a greater but less <strong>in</strong>tensively<br />
active photosynthetic biomass should not be<br />
supported to do the same work, though only to the<br />
limit of its ma<strong>in</strong>tenance costs. Tak<strong>in</strong>g the basal<br />
respiration of Chlorella to be 1.1 × 10 –6 mol C (mol<br />
cell C) –1 s –1 (Reynolds 1990), the daily energetic cost<br />
of replac<strong>in</strong>g 0.095 mol C (mol cell C) –1 was calculated<br />
to be about 1.2 MJ (mol cell C) –1 , mean<strong>in</strong>g that



