

The utility of song and morphological characters in delineating ...
The utility of song and morphological characters in delineating ...
The utility of song and morphological characters in delineating ...

Create successful ePaper yourself
Turn your PDF publications into a flip-book with our unique Google optimized e-Paper software.
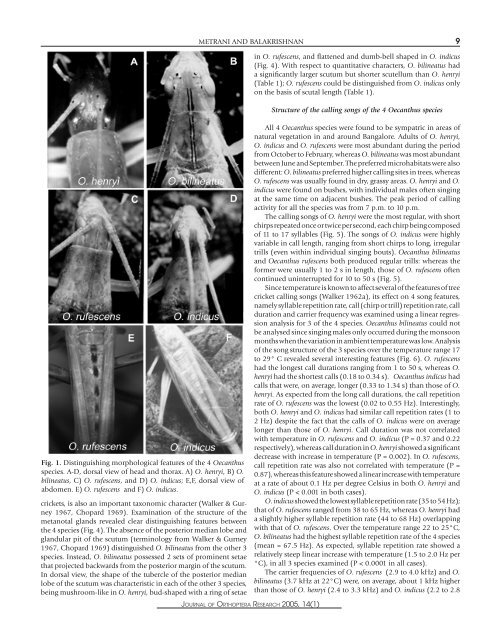
Fig. 1. Dist<strong>in</strong>guish<strong>in</strong>g <strong>morphological</strong> features <strong>of</strong> the 4 Oecanthusspecies. A-D, dorsal view <strong>of</strong> head <strong>and</strong> thorax. A) O. henryi, B) O.bil<strong>in</strong>eatus, C) O. rufescens, <strong>and</strong> D) O. <strong>in</strong>dicus; E,F, dorsal view <strong>of</strong>abdomen. E) O. rufescens <strong>and</strong> F) O. <strong>in</strong>dicus.crickets, is also an important taxonomic character (Walker & Gurney1967, Chopard 1969). Exam<strong>in</strong>ation <strong>of</strong> the structure <strong>of</strong> themetanotal gl<strong>and</strong>s revealed clear dist<strong>in</strong>guish<strong>in</strong>g features betweenthe 4 species (Fig. 4). <strong>The</strong> absence <strong>of</strong> the posterior median lobe <strong>and</strong>gl<strong>and</strong>ular pit <strong>of</strong> the scutum (term<strong>in</strong>ology from Walker & Gurney1967, Chopard 1969) dist<strong>in</strong>guished O. bil<strong>in</strong>eatus from the other 3species. Instead, O. bil<strong>in</strong>eatus possessed 2 sets <strong>of</strong> prom<strong>in</strong>ent setaethat projected backwards from the posterior marg<strong>in</strong> <strong>of</strong> the scutum.In dorsal view, the shape <strong>of</strong> the tubercle <strong>of</strong> the posterior medianlobe <strong>of</strong> the scutum was characteristic <strong>in</strong> each <strong>of</strong> the other 3 species,be<strong>in</strong>g mushroom-like <strong>in</strong> O. henryi, bud-shaped with a r<strong>in</strong>g <strong>of</strong> setaeMETRANI AND BALAKRISHNAN 9JOURNAL OF ORTHOPTERA RESEARCH 2005, 14(1)<strong>in</strong> O. rufescens, <strong>and</strong> flattened <strong>and</strong> dumb-bell shaped <strong>in</strong> O. <strong>in</strong>dicus(Fig. 4). With respect to quantitative <strong>characters</strong>, O. bil<strong>in</strong>eatus hada significantly larger scutum but shorter scutellum than O. henryi(Table 1); O. rufescens could be dist<strong>in</strong>guished from O. <strong>in</strong>dicus onlyon the basis <strong>of</strong> scutal length (Table 1).Structure <strong>of</strong> the call<strong>in</strong>g <strong>song</strong>s <strong>of</strong> the 4 Oecanthus speciesAll 4 Oecanthus species were found to be sympatric <strong>in</strong> areas <strong>of</strong>natural vegetation <strong>in</strong> <strong>and</strong> around Bangalore. Adults <strong>of</strong> O. henryi,O. <strong>in</strong>dicus <strong>and</strong> O. rufescens were most abundant dur<strong>in</strong>g the periodfrom October to February, whereas O. bil<strong>in</strong>eatus was most abundantbetween June <strong>and</strong> September. <strong>The</strong> preferred microhabitats were alsodifferent: O. bil<strong>in</strong>eatus preferred higher call<strong>in</strong>g sites <strong>in</strong> trees, whereasO. rufescens was usually found <strong>in</strong> dry, grassy areas. O. henryi <strong>and</strong> O.<strong>in</strong>dicus were found on bushes, with <strong>in</strong>dividual males <strong>of</strong>ten s<strong>in</strong>g<strong>in</strong>gat the same time on adjacent bushes. <strong>The</strong> peak period <strong>of</strong> call<strong>in</strong>gactivity for all the species was from 7 p.m. to 10 p.m.<strong>The</strong> call<strong>in</strong>g <strong>song</strong>s <strong>of</strong> O. henryi were the most regular, with shortchirps repeated once or twice per second, each chirp be<strong>in</strong>g composed<strong>of</strong> 11 to 17 syllables (Fig. 5). <strong>The</strong> <strong>song</strong>s <strong>of</strong> O. <strong>in</strong>dicus were highlyvariable <strong>in</strong> call length, rang<strong>in</strong>g from short chirps to long, irregulartrills (even with<strong>in</strong> <strong>in</strong>dividual s<strong>in</strong>g<strong>in</strong>g bouts). Oecanthus bil<strong>in</strong>eatus<strong>and</strong> Oecanthus rufescens both produced regular trills: whereas theformer were usually 1 to 2 s <strong>in</strong> length, those <strong>of</strong> O. rufescens <strong>of</strong>tencont<strong>in</strong>ued un<strong>in</strong>terrupted for 10 to 50 s (Fig. 5).S<strong>in</strong>ce temperature is known to affect several <strong>of</strong> the features <strong>of</strong> treecricket call<strong>in</strong>g <strong>song</strong>s (Walker 1962a), its effect on 4 <strong>song</strong> features,namely syllable repetition rate, call (chirp or trill) repetition rate, callduration <strong>and</strong> carrier frequency was exam<strong>in</strong>ed us<strong>in</strong>g a l<strong>in</strong>ear regressionanalysis for 3 <strong>of</strong> the 4 species. Oecanthus bil<strong>in</strong>eatus could notbe analysed s<strong>in</strong>ce s<strong>in</strong>g<strong>in</strong>g males only occurred dur<strong>in</strong>g the monsoonmonths when the variation <strong>in</strong> ambient temperature was low. Analysis<strong>of</strong> the <strong>song</strong> structure <strong>of</strong> the 3 species over the temperature range 17to 29° C revealed several <strong>in</strong>terest<strong>in</strong>g features (Fig. 6). O. rufescenshad the longest call durations rang<strong>in</strong>g from 1 to 50 s, whereas O.henryi had the shortest calls (0.18 to 0.34 s). Oecanthus <strong>in</strong>dicus hadcalls that were, on average, longer (0.33 to 1.34 s) than those <strong>of</strong> O.henryi. As expected from the long call durations, the call repetitionrate <strong>of</strong> O. rufescens was the lowest (0.02 to 0.55 Hz). Interest<strong>in</strong>gly,both O. henryi <strong>and</strong> O. <strong>in</strong>dicus had similar call repetition rates (1 to2 Hz) despite the fact that the calls <strong>of</strong> O. <strong>in</strong>dicus were on averagelonger than those <strong>of</strong> O. henryi. Call duration was not correlatedwith temperature <strong>in</strong> O. rufescens <strong>and</strong> O. <strong>in</strong>dicus (P = 0.37 <strong>and</strong> 0.22respectively), whereas call duration <strong>in</strong> O. henryi showed a significantdecrease with <strong>in</strong>crease <strong>in</strong> temperature (P = 0.002). In O. rufescens,call repetition rate was also not correlated with temperature (P =0.87), whereas this feature showed a l<strong>in</strong>ear <strong>in</strong>crease with temperatureat a rate <strong>of</strong> about 0.1 Hz per degree Celsius <strong>in</strong> both O. henryi <strong>and</strong>O. <strong>in</strong>dicus (P < 0.001 <strong>in</strong> both cases).O. <strong>in</strong>dicus showed the lowest syllable repetition rate (35 to 54 Hz);that <strong>of</strong> O. rufescens ranged from 38 to 65 Hz, whereas O. henryi hada slightly higher syllable repetition rate (44 to 68 Hz) overlapp<strong>in</strong>gwith that <strong>of</strong> O. rufescens. Over the temperature range 22 to 25°C,O. bil<strong>in</strong>eatus had the highest syllable repetition rate <strong>of</strong> the 4 species(mean = 67.5 Hz). As expected, syllable repetition rate showed arelatively steep l<strong>in</strong>ear <strong>in</strong>crease with temperature (1.5 to 2.0 Hz per°C), <strong>in</strong> all 3 species exam<strong>in</strong>ed (P < 0.0001 <strong>in</strong> all cases).<strong>The</strong> carrier frequencies <strong>of</strong> O. rufescens (2.9 to 4.0 kHz) <strong>and</strong> O.bil<strong>in</strong>eatus (3.7 kHz at 22°C) were, on average, about 1 kHz higherthan those <strong>of</strong> O. henryi (2.4 to 3.3 kHz) <strong>and</strong> O. <strong>in</strong>dicus (2.2 to 2.8













