Plasticité synaptique à long terme
Plasticité synaptique à long terme
Plasticité synaptique à long terme

Create successful ePaper yourself
Turn your PDF publications into a flip-book with our unique Google optimized e-Paper software.
Enregistrement<br />
électrophysiologique<br />
Stimulation électrique<br />
de l’axone<br />
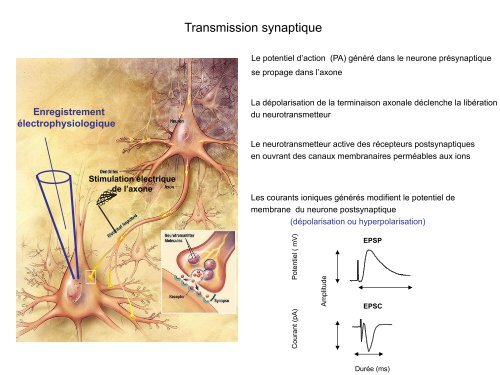
Transmission <strong>synaptique</strong><br />
Le potentiel d’action (PA) généré dans le neurone pré<strong>synaptique</strong><br />
se propage dans l’axone<br />
La dépolarisation de la terminaison axonale déclenche la libération<br />
du neurotransmetteur<br />
Le neurotransmetteur active des récepteurs post<strong>synaptique</strong>s<br />
en ouvrant des canaux membranaires perméables aux ions<br />
Les courants ioniques générés modifient le potentiel de<br />
membrane du neurone post<strong>synaptique</strong><br />
(dépolarisation ou hyperpolarisation)<br />
Potentiel ( mV)<br />
Courant (pA)<br />
Amplitude<br />
EPSP<br />
EPSC<br />
Durée (ms)
mV<br />
pA<br />
Dépression<br />
<strong>Plasticité</strong> <strong>synaptique</strong>:<br />
Modification de l’efficacité de la transmission <strong>synaptique</strong><br />
EPSP<br />
EPSC<br />
Potentialisation
<strong>Plasticité</strong> <strong>à</strong> court <strong>terme</strong> <strong>Plasticité</strong> <strong>à</strong> <strong>long</strong> <strong>terme</strong><br />
milli secondes - minutes minutes - jours<br />
Paired pulse facilitation<br />
(PPF)<br />
Paired pulse depression<br />
(PPD)<br />
milli secondes<br />
<strong>Plasticité</strong> <strong>synaptique</strong>:<br />
Modification de l’efficacité de la transmission <strong>synaptique</strong><br />
Post tetanic potentiation<br />
(PTP)<br />
Post tetanic depression<br />
(PTD)<br />
secondes<br />
Long term potentiation<br />
(LTP)<br />
Long term depression<br />
(LTD)
Facilitation<br />
<strong>Plasticité</strong> <strong>synaptique</strong> <strong>à</strong> court <strong>terme</strong><br />
EPSP<br />
Potentiels d’action pré<strong>synaptique</strong>s
Dépression<br />
Stimulation<br />
pré<strong>synaptique</strong><br />
<strong>Plasticité</strong> <strong>synaptique</strong> <strong>à</strong> court <strong>terme</strong>
Post tetanic potentiation<br />
Amplitude<br />
<strong>Plasticité</strong> <strong>synaptique</strong> <strong>à</strong> court <strong>terme</strong><br />
pré<strong>synaptique</strong>
<strong>Plasticité</strong> <strong>synaptique</strong> <strong>à</strong> court <strong>terme</strong><br />
Mécanismes<br />
Mécanismes<br />
Pré<strong>synaptique</strong>s<br />
(facilitation et dépression)<br />
Mécanismes<br />
Post<strong>synaptique</strong>s<br />
(dépression)
Le Ca 2+ qui entre dans la terminaison<br />
pré<strong>synaptique</strong> par les canaux calcique<br />
dépendent du voltage (VDCC) est<br />
responsable de la libération du neurotransmetteur<br />
<strong>Plasticité</strong> <strong>synaptique</strong> <strong>à</strong> court <strong>terme</strong><br />
Mécanismes pré<strong>synaptique</strong>s (facilitation)<br />
VDCC<br />
PA<br />
La probabilité de libération du neurotransmetteur,<br />
est proportionnelle <strong>à</strong> la quantité de Ca 2+ libre dans<br />
la terminaison pré<strong>synaptique</strong><br />
Ca 2+
Le Ca 2+ qui entre dans la terminaison<br />
pré<strong>synaptique</strong> par les canaux calcique<br />
dépendent du voltage (VDCC) est<br />
responsable de la libération du neurotransmetteur<br />
<strong>Plasticité</strong> <strong>synaptique</strong> <strong>à</strong> court <strong>terme</strong><br />
Mécanismes pré<strong>synaptique</strong>s (facilitation)<br />
VDCC<br />
EPSP<br />
Ca 2+<br />
PA<br />
PA<br />
La probabilité de libération du neurotransmetteur,<br />
est proportionnelle <strong>à</strong> la quantité de Ca 2+ libre dans<br />
la terminaison pré<strong>synaptique</strong><br />
Ca 2+<br />
Le Ca 2+ libre augmente pendante une<br />
activité pré<strong>synaptique</strong> <strong>à</strong> haute fréquence
<strong>Plasticité</strong> <strong>synaptique</strong> <strong>à</strong> court <strong>terme</strong><br />
Mécanismes pré<strong>synaptique</strong>s (facilitation)<br />
Mécanisme 1: un élargissement des PAs se produit pendant une activité <strong>à</strong> haute fréquence<br />
ce qui détermine un augmentation de Ca 2+ dans la terminaison <strong>synaptique</strong><br />
Pré<strong>synaptique</strong><br />
Post<strong>synaptique</strong><br />
Debanne 2004<br />
L’élargissement des PAs est due <strong>à</strong><br />
l’inactivation des canaux potassiques<br />
pendant l’activité <strong>à</strong> haute fréquence<br />
(Canaux K-type A)
Na +<br />
Le PA est dû <strong>à</strong> l’ouverture séquentielle de canaux Na + et K +<br />
Un stimulus dépolarise la membrane<br />
Na +<br />
K +<br />
K +<br />
ext<br />
int<br />
Vm en mV<br />
+ 40<br />
0<br />
- 40<br />
- 80<br />
Temps (ms)
Na +<br />
Le PA est dû <strong>à</strong> l’ouverture séquentielle de canaux Na + et K +<br />
Na +<br />
Na + Na +<br />
K +<br />
K +<br />
K +<br />
Si la dépolarisation atteint le seuil de - 40 mV, les<br />
canaux Na + s’ouvrent ⇒ entrée d’ions Na + positifs<br />
⇒ inversion de Vm<br />
ext<br />
int<br />
Vm en mV<br />
+ 40<br />
0<br />
- 40<br />
- 80<br />
seuil<br />
Temps (ms)
Na +<br />
Le PA est dû <strong>à</strong> l’ouverture séquentielle de canaux Na + et K +<br />
Na +<br />
Na + Na + Na + Na +<br />
K +<br />
+ K+<br />
K<br />
K +<br />
ext<br />
int<br />
Vm en mV<br />
+ 40<br />
0<br />
- 40<br />
- 80<br />
repolarisation<br />
Temps (ms)<br />
Les canaux Na+ s’inactivent très<br />
rapidement.<br />
Les canaux K + s’ouvrent <strong>à</strong> leur tour<br />
sous l’effet de la dépolarisation ⇒<br />
sortie d’ions K + . Comme l’intérieur<br />
de l’axone perd des charges +, il<br />
redevient négatif par rapport <strong>à</strong><br />
l’extérieur (repolarisation).
Na +<br />
Le PA est dû <strong>à</strong> l’ouverture séquentielle de canaux Na + et K +<br />
Na +<br />
Na + Na + Na + Na +<br />
K +<br />
+ K+<br />
K<br />
K +<br />
ext<br />
int<br />
Vm en mV<br />
+ 40<br />
0<br />
- 40<br />
- 80<br />
hyperpolarisation<br />
Temps (ms)<br />
Avant que les canaux K + ne se<br />
ferment sous l’effet de la<br />
repolarisation, la sortie des ions K +<br />
continue ⇒ hyperpolarisation de la<br />
membrane.<br />
Na + Na +<br />
K +
Na +<br />
Le PA est dû <strong>à</strong> l’ouverture séquentielle de canaux Na + et K +<br />
Na +<br />
Na + Na + Na + Na +<br />
K +<br />
K + K+<br />
K +<br />
ext<br />
int<br />
+ 40<br />
0<br />
- 40<br />
- 80<br />
Activation des canaux K +<br />
Vm en mV<br />
Retour au potentiel de repos.<br />
Inactivation des canaux K +<br />
Temps (ms)<br />
Na + Na +<br />
K +
<strong>Plasticité</strong> <strong>synaptique</strong> <strong>à</strong> court <strong>terme</strong><br />
Mécanismes pré<strong>synaptique</strong>s (facilitation)<br />
Mécanisme 2: Le Ca 2+ produit par les premiers PAs est tamponné par des protéines du cytoplasme<br />
(Calcium binding protein ; ex. Calbinding-D28K; parvalbumine)<br />
Calcium binding protein (CBP)<br />
1 er potentiel d’action<br />
1 er EPSP<br />
PA<br />
Une partie du Ca 2+ est séquestré par les CBP<br />
2 em potentiel d’action<br />
2 em EPSP<br />
PA<br />
Les CBP sont saturés. Tout le Ca 2+ participe <strong>à</strong> la<br />
libération du neurotransmetteur
<strong>Plasticité</strong> <strong>synaptique</strong> <strong>à</strong> court <strong>terme</strong><br />
Mécanismes pré<strong>synaptique</strong>s (facilitation)<br />
Mécanisme 3: Une partie du Ca 2+ n’arrive pas <strong>à</strong> être extrudé avant l’arrivée des PAs suivants<br />
(Ca 2+ résiduelle)<br />
Na +<br />
Ca 2+<br />
1er potentiel d’action<br />
+ 2+<br />
Na -Ca excanger<br />
ER<br />
PA<br />
Mitocondre<br />
2+<br />
Ca ATPase<br />
Le Ca 2+ entré par les VDCC est<br />
extrudé par les transporteurs du Ca 2+<br />
Ca 2+ ATPase<br />
Na + -Ca 2+ exchanger ( 3 Na + par 1 Ca 2+ )<br />
EPSP
<strong>Plasticité</strong> <strong>synaptique</strong> <strong>à</strong> court <strong>terme</strong><br />
Mécanismes pré<strong>synaptique</strong>s (facilitation)<br />
Mécanisme 3: Une partie du Ca 2+ n’arrive pas <strong>à</strong> être extrudé avant l’arrivé des PAs suivants<br />
(Ca 2+ résiduelle)<br />
Na +<br />
Ca 2+<br />
+ 2+<br />
Na -Ca excanger<br />
ER<br />
PA<br />
Mitocondre<br />
En fonction de la vitesse d’extrusions, du<br />
Ca 2+ peut rester dans la terminaison au<br />
moment de l’arrivée du PA suivant<br />
2+<br />
Ca ATPase
<strong>Plasticité</strong> <strong>synaptique</strong> <strong>à</strong> court <strong>terme</strong><br />
Mécanismes pré<strong>synaptique</strong>s (facilitation)<br />
Mécanisme 3: Une partie du Ca 2+ n’arrive pas <strong>à</strong> être extrudé avant l’arrivé des PAs suivants<br />
(Ca 2+ résiduel)<br />
Na +<br />
Ca 2+<br />
2nd potentiel d’action<br />
+ 2+<br />
Na -Ca excanger<br />
ER<br />
PA<br />
Mitocondre<br />
L’entré de Ca 2+ déclenché par le PA suivant<br />
s’additionne au Ca 2+ résiduel. Cela augmente<br />
la quantité de neurotransmetteur libéré<br />
(par rapport au PA précédent).<br />
2+<br />
Ca ATPase<br />
EPSP
Na +<br />
Ca 2+<br />
Canal Na<br />
+<br />
Canal Ca 2+<br />
Le PA produit l’entrée de Na +<br />
par les canaux Na<br />
Espace<br />
extracellulaire<br />
Na+-Ca 2+ exchanger<br />
+ 2+<br />
Na -Ca excanger<br />
Espace<br />
intracellulaire<br />
<strong>Plasticité</strong> <strong>synaptique</strong> <strong>à</strong> court <strong>terme</strong><br />
Mécanismes pré<strong>synaptique</strong>s (PTP)<br />
PA<br />
Avec une faible [Na + ] intracellulaire l’échangeur Na + -Ca 2+<br />
fait sortir le Ca 2+ et entrer le Na +
Na +<br />
Ca 2+<br />
Canal Na<br />
+<br />
Canal Ca 2+<br />
Le « tetanus » produit une accumulation<br />
des ions Na +<br />
Espace<br />
extracellulaire<br />
+ 2+<br />
Na -Ca exchanger<br />
Espace<br />
intracellulaire<br />
<strong>Plasticité</strong> <strong>synaptique</strong> <strong>à</strong> court <strong>terme</strong><br />
Mécanismes pré<strong>synaptique</strong>s (PTP)<br />
PA<br />
Avec une [Na + ] intracellulaire élevé l’échangeur Na + -Ca 2+<br />
inverse son activité
Activité <strong>à</strong> haute fréquence<br />
pro<strong>long</strong>ée (Tetanus)<br />
Accumulation de ions Na +<br />
Inactivation/inversion<br />
échangeur Na + -Ca 2+<br />
Accumulation de Ca2+<br />
<strong>Plasticité</strong> <strong>synaptique</strong> <strong>à</strong> court <strong>terme</strong><br />
Mécanismes pré<strong>synaptique</strong>s (PTP)<br />
Post tetanic potentiation
<strong>Plasticité</strong> <strong>synaptique</strong> <strong>à</strong> court <strong>terme</strong><br />
Mécanismes pré<strong>synaptique</strong>s (dépression)<br />
Mécanisme1: L’inactivation de canaux Na + réduit les PAs pré<strong>synaptique</strong>s<br />
Réduction des PAs<br />
pré<strong>synaptique</strong>s<br />
Dépression de la<br />
transmission post<strong>synaptique</strong>
Mécanisme2: Déplétion du neurotransmetteur<br />
Readily Releasable Pool<br />
Ensemble de vésicules de<br />
neurotransmetteur ancré <strong>à</strong> la<br />
membrane prêtent <strong>à</strong> être libéré<br />
<strong>Plasticité</strong> <strong>synaptique</strong> <strong>à</strong> court <strong>terme</strong><br />
Mécanismes pré<strong>synaptique</strong>s (dépression)
Mécanisme2: Déplétion du neurotransmetteur<br />
Readily Releasable Pool avant le 1 er PA<br />
<strong>Plasticité</strong> <strong>synaptique</strong> <strong>à</strong> court <strong>terme</strong><br />
Mécanismes pré<strong>synaptique</strong>s (dépression)<br />
Reserve pool<br />
1 er PA<br />
EPSP<br />
PA<br />
La libération du neurotransmetteur<br />
est proportionnel au Readily Releasable Pool
Mécanisme2: Déplétion du neurotransmetteur<br />
Reserve pool<br />
<strong>Plasticité</strong> <strong>synaptique</strong> <strong>à</strong> court <strong>terme</strong><br />
Mécanismes pré<strong>synaptique</strong>s (dépression)<br />
Le Readily Releasable Pool est réduite après<br />
la libération<br />
2 em PA<br />
EPSP<br />
PA<br />
La libération du neurotransmetteur<br />
est réduite
<strong>Plasticité</strong> <strong>synaptique</strong> <strong>à</strong> court <strong>terme</strong><br />
Mécanismes post<strong>synaptique</strong>s (dépression)<br />
Mécanisme3: Activation des auto récepteurs métabotropiques<br />
L’activation des récepteurs GABA B , qui est activé par le GABA libéré pendant le 1 er PA,<br />
réduit la probabilité de libération de GABA pendant le 2nd PA<br />
2nd 1st Stimulus<br />
GABA B<br />
GABA A<br />
Cl -<br />
1st stimulus 2nd stimulus<br />
50 pA<br />
100 ms<br />
L’activation des GABA B active une voie de signalisation intracellulaire qui abouti<br />
<strong>à</strong> l’inactivation de canaux calciques pré<strong>synaptique</strong>s<br />
Crt<br />
CGP<br />
35348
<strong>Plasticité</strong> <strong>synaptique</strong> <strong>à</strong> court <strong>terme</strong><br />
Mécanismes post<strong>synaptique</strong>s (dépression)<br />
écanisme1 : Désensibilisation des récepteurs post<strong>synaptique</strong>s<br />
les récepteurs activés par les neurotransmetteurs<br />
libérés pendant le 1 er PA sont désensibilisés<br />
Moins des récepteurs sont activés<br />
pendant le 2 nd PA
<strong>Plasticité</strong> <strong>synaptique</strong> <strong>à</strong> court <strong>terme</strong><br />
Mécanismes post<strong>synaptique</strong>s (dépression)<br />
Mécanisme 2 : Saturation des récepteurs post<strong>synaptique</strong><br />
es neurotransmetteurs libérés pendant le 1 er PA<br />
restent liés aux récepteurs<br />
Moins des récepteurs sont activés<br />
pendant le 2 nd PA
<strong>Plasticité</strong> <strong>synaptique</strong> <strong>à</strong> court <strong>terme</strong><br />
Rôle fonctionnelle<br />
Plusieurs mécanisme de plasticité <strong>à</strong> cour <strong>terme</strong> peuvent<br />
coexister dans la même synapse<br />
La plasticité <strong>à</strong> court <strong>terme</strong> change en fonction<br />
des différentes conditions environnementales:<br />
Le type de synapse<br />
La fréquence d’activation<br />
Facteurs modulatoires<br />
La plasticité <strong>synaptique</strong> permet de modifier de<br />
façon dynamique les propriétés<br />
de la transmission <strong>synaptique</strong><br />
Climbing fiber (cervelet)<br />
Parallel fiber (cervelet)<br />
Shaffer collateral (hippocampe)
La plasticité <strong>à</strong> court <strong>terme</strong> permet<br />
de filtrer les signaux qui passent du<br />
neurone pré<strong>synaptique</strong> au neurone<br />
post<strong>synaptique</strong><br />
<strong>Plasticité</strong> <strong>synaptique</strong> <strong>à</strong> court <strong>terme</strong><br />
Fonction<br />
(Filtrage de l’information neuronale)<br />
PA pré<strong>synaptique</strong>s<br />
Facilitation (high pass filter)<br />
PA post<strong>synaptique</strong>s<br />
Depression (low pass filter)<br />
PA post<strong>synaptique</strong>s<br />
Seuil pour la gホnホration des PA<br />
Seuil pour la gホnホration des PA
Climbing fiber (cervelet)<br />
Parallel fiber (cervelet)<br />
Shaffer collateral (hippocampe)<br />
<strong>Plasticité</strong> <strong>synaptique</strong> <strong>à</strong> court <strong>terme</strong><br />
Rôle fonctioenelle<br />
(Filtrage de l’information neuronale)<br />
Low-pass filter (activité du neurone pré<strong>synaptique</strong> < 10 Hz)<br />
High-pass filter (activité du neurone pré<strong>synaptique</strong> > 20 Hz)<br />
Band-pass filter (2 Hz > activité du neurone pré<strong>synaptique</strong> < 30 Hz)
<strong>Plasticité</strong> <strong>synaptique</strong> <strong>à</strong> court <strong>terme</strong>
<strong>Plasticité</strong> <strong>synaptique</strong> <strong>à</strong> <strong>long</strong> <strong>terme</strong><br />
Changement persistant de l’efficacité de la transmission <strong>synaptique</strong><br />
entre deux ou plusieurs neurones<br />
Modification de la connectivité du réseau neuronale<br />
Base cellulaire de la mémoire
<strong>Plasticité</strong> <strong>synaptique</strong> <strong>à</strong> <strong>long</strong> <strong>terme</strong><br />
Base cellulaire de la mémoire<br />
(évidences expérimentales)<br />
-Les altérations de la plasticité <strong>synaptique</strong> sont associé <strong>à</strong> des altérations<br />
des processus mnésiques<br />
-Certaines formes de mémoires produisent de la plasticité <strong>synaptique</strong>
<strong>Plasticité</strong> <strong>synaptique</strong> dans les processus<br />
de mémoire implicite (procédurale)<br />
Acquisition et l'utilisation de compétences motrices<br />
(processus inconscients)<br />
Aplysie<br />
limace de mer<br />
<strong>Plasticité</strong> <strong>synaptique</strong> <strong>à</strong> <strong>long</strong> <strong>terme</strong><br />
<strong>Plasticité</strong> <strong>synaptique</strong> dans les processus<br />
de mémoire explicite (déclarative)<br />
Souvenir des expériences et informations acquises<br />
(processus conscients)<br />
Hippocampe
<strong>Plasticité</strong> <strong>synaptique</strong> <strong>à</strong> <strong>long</strong> <strong>terme</strong><br />
Manteau<br />
Sensibilisation de l’aplysie<br />
Branchie<br />
Stimulation<br />
tactile<br />
La stimulation tactile du siphon produit<br />
la rétraction du siphon lui-même<br />
et de la branchie<br />
Stimulation<br />
tactile<br />
Shock<br />
électrique<br />
<strong>à</strong> la queue<br />
Un shock électrique (stimulation douloureuse)<br />
de la queue augment la réponse de rétraction, produit<br />
par les stimulations tactiles suivantes.<br />
(Sensibilisation)<br />
La durée de la sensibilisation est fonction des nombre de chocs reçus par la limace:<br />
4 chocs électriques : sensibilisation <strong>à</strong> court <strong>terme</strong><br />
4 trains de chocs électriques: sensibilisation <strong>à</strong> <strong>long</strong> <strong>terme</strong><br />
D’âpres Kandel; Science 2001
Queue<br />
<strong>Plasticité</strong> <strong>synaptique</strong> <strong>à</strong> <strong>long</strong> <strong>terme</strong><br />
Sensibilisation de l’aplysie mécanismes neuronales<br />
Neurones sérotoninergiques activés<br />
par le choc électrique<br />
Branchie<br />
Neurones sensoriels activés<br />
par la stimulation du siphon<br />
Neurones moteurs activés<br />
par les neurones sensoriels<br />
Le phénomène de sensibilisation est due <strong>à</strong> l’augmentation de l’efficacité <strong>synaptique</strong><br />
au niveau des synapses entre les neurones sensoriels et moteurs<br />
Induite par l’action des neurones sérotoninergiques sur les neurones sensoriels
EPSP<br />
<strong>Plasticité</strong> <strong>synaptique</strong> <strong>à</strong> <strong>long</strong> <strong>terme</strong><br />
Sensibilisation de l’aplysie : mécanismes nerveux<br />
Sensibilisation <strong>à</strong> court <strong>terme</strong><br />
- La 5HT active adenylate cyclase qui<br />
produit l’AMP cyclique (cAMP)<br />
- Le cAMP active la protéine kinase A<br />
(PKA) qui phosphoryle et inactive les<br />
canaux K + (élargissement de PAs).<br />
La PKA agit aussi sur la<br />
libération du neurotransmetteur
<strong>Plasticité</strong> <strong>synaptique</strong> <strong>à</strong> <strong>long</strong> <strong>terme</strong><br />
Sensibilisation de l’aplysie : mécanismes nerveux<br />
Sensibilisation <strong>à</strong> <strong>long</strong> <strong>terme</strong><br />
-Avec la stimulation répétée des<br />
récepteurs 5HT, la PKA phosphoryle<br />
la protéine MAPK qui entre dan le noyau<br />
ou phosphoryle et active le facteur<br />
de transcription CREB qui favorise la<br />
transcriptions des gènes sous le promoteur<br />
CRE ( cAMP Réponses Element)<br />
-L’expression du gène précoce<br />
pur l’Ubiquitaire Hydrolase amène <strong>à</strong> la<br />
dégradation de la sous-unité régulatrice de<br />
la PKA( PKA activé de manière constitutive)<br />
-L’expression des gènes tardifs<br />
entraine des modifications structurales<br />
qui augmentent l’efficacité <strong>synaptique</strong><br />
(augmentation des varicosités <strong>synaptique</strong>s)<br />
La sensibilisation <strong>à</strong> <strong>long</strong> <strong>terme</strong> nécessite la synthèse de nouvelles protéines
<strong>Plasticité</strong> <strong>synaptique</strong> <strong>à</strong> <strong>long</strong> <strong>terme</strong><br />
Long Term Potentiation (LTP) dan l’hippocampe; CA1<br />
1- La réponse <strong>synaptique</strong>, générée par la stimulation électrique<br />
des collatérales de Schaffer :<br />
l’amplitude des EPSPS est stable au cours du temps<br />
2- Une bref train de stimulation <strong>à</strong> haute fréquence « Tetanus »<br />
est produit sur les collatérales de Schaffer<br />
3- La réponse <strong>synaptique</strong> est augmentée après le « Tetanus »<br />
D’âpres Hammond; Cellular and Molecular Neurophysiology
<strong>Plasticité</strong> <strong>synaptique</strong> <strong>à</strong> <strong>long</strong> <strong>terme</strong><br />
Long Term Potentiation (LTP) dans l’hippocampe; CA1<br />
La duré de la LTP dépend de l’intensité du Tetanus (nombre de trains)<br />
Un train induit une LTP de courte durée (quelques heures), LTP précoce<br />
Quatre trains induisent une LTP de <strong>long</strong>ue durée (plusieurs jours); LTP tardive<br />
La phase tardive de LTP dépende de la PKA, du facteur CREB et de la synthèse de nouvelle protéines<br />
D’âpres Kandel; Science 2001
<strong>Plasticité</strong> <strong>synaptique</strong> <strong>à</strong> <strong>long</strong> <strong>terme</strong><br />
Long Term Potentiation (LTP) dans l’hippocampe; CA1<br />
LTP est spécifique : seule la synapse qui reçoit le tétanus est potentialisée<br />
D’âpres Hammond; Cellular and Molecular Neurophysiology
<strong>Plasticité</strong> <strong>synaptique</strong> <strong>à</strong> <strong>long</strong> <strong>terme</strong><br />
Background théorique<br />
Postulat de Hebb dans la formation des processus de mémoire « stabilisation des ensembles neuronaux »<br />
(Donald Hebb;1949):<br />
Quand l’activité du neurone A contribue de manière répétée <strong>à</strong> la génération des PAs dans le neurone B<br />
la connectivité fonctionnelle entre A et B est renforcée<br />
A B<br />
Pré Tetanus<br />
Post Tetanus<br />
Tetanus<br />
L’activité du neurone A fa décharger le neurone B
<strong>Plasticité</strong> <strong>synaptique</strong> <strong>à</strong> <strong>long</strong> <strong>terme</strong><br />
Long Term Potentiation (LTP) dan l’hippocampe; CA1<br />
La plasticité <strong>synaptique</strong> dans la région CA1 est de type Hebbian<br />
Seule la synapse qui fait décharger le neurone post<strong>synaptique</strong> est potentialisée
Ion Mg +<br />
Glutamate<br />
Ion Ca 2+<br />
<strong>Plasticité</strong> <strong>synaptique</strong> <strong>à</strong> <strong>long</strong> <strong>terme</strong><br />
Long Term Potentiation (LTP) dan l’hippocampe; CA1<br />
Les récepteurs N-methyl D-aspartate (NMDA) sont des détecteurs de la coïncidence<br />
de l’activation pre et post <strong>synaptique</strong><br />
- - - - - - - - - - - - - - -<br />
++++++<br />
Dépolarisation<br />
post<strong>synaptique</strong><br />
++++++<br />
++++++<br />
- - - - -<br />
++++++<br />
<strong>Plasticité</strong> <strong>synaptique</strong>
<strong>Plasticité</strong> <strong>synaptique</strong> <strong>à</strong> <strong>long</strong> <strong>terme</strong><br />
Long Term Potentiation (LTP) dan l’hippocampe; CA1<br />
L’antagoniste du récepteur NMDA D-2-Amino-5-phosphonovalerate (D-APV)<br />
empêche l’induction de la LTP dans la région CA1 de l’hippocampe
<strong>Plasticité</strong> <strong>synaptique</strong> <strong>à</strong> <strong>long</strong> <strong>terme</strong><br />
Long Term Potentiation (LTP) dans l’hippocampe; CA1<br />
Dans des souris transgéniques dont les courants NMDA sont plus grands la LTP est augmentée<br />
Curant NMDA control<br />
Curant NMDA transgénique ( sous-unité NR2B augmenté)<br />
La LTP est plus grande dans les souris transgéniques Une stimulation <strong>à</strong> faible fréquence induit la LTP dans les souris<br />
Souris control Souris transgéniques<br />
transgéniques<br />
D’âpres Tang; Nature 1999
<strong>Plasticité</strong> <strong>synaptique</strong> <strong>à</strong> <strong>long</strong> <strong>terme</strong><br />
Long Term Potentiation (LTP) dans l’hippocampe; CA1<br />
Les souris transgénique pour le récepteur NMDA ont une plus grande rétention de mémoire de l’information<br />
Son<br />
(Stimulus conditionnel)<br />
Immobilisation conditionné ( %)<br />
choc électrique<br />
( Stimulus inconditionnel)<br />
Immobilisation par la peur<br />
(Réponse conditionné)<br />
D’âpres Tang; Nature 1999
<strong>Plasticité</strong> <strong>synaptique</strong> <strong>à</strong> <strong>long</strong> <strong>terme</strong><br />
Long Term Depression (LTD) dans l’hippocampe; CA1<br />
Une LTD peut être induite par une stimulation répétée <strong>à</strong> faible fréquence (1 Hz)<br />
D’âpres Dudek ; PNAS 1992
<strong>Plasticité</strong> <strong>synaptique</strong> <strong>à</strong> <strong>long</strong> <strong>terme</strong><br />
Long Term Depression (LTD) dans l’hippocampe; CA1<br />
La LTD dépend elle aussi de l’activation des récepteurs NMDA<br />
D’âpres Dudek ; PNAS 1992
<strong>Plasticité</strong> <strong>synaptique</strong> <strong>à</strong> <strong>long</strong> <strong>terme</strong><br />
LTP Vs LTD dans l’hippocampe; CA1<br />
La forme de plasticité (LTP/LTD) dépend de la concentration de Ca 2+ produit par la stimulation<br />
Stimulation <strong>à</strong> faible fréquence<br />
LTD<br />
Stimulation <strong>à</strong> haute fréquence<br />
LTP
Modification de la probabilité de libération du<br />
neurotransmetteur<br />
Modifications structurales<br />
Changement du nombre de connexions<br />
<strong>synaptique</strong>s<br />
<strong>Plasticité</strong> <strong>synaptique</strong> <strong>à</strong> <strong>long</strong> <strong>terme</strong><br />
Site d’expression<br />
Modification du nombre ou des propriétés des récepteurs<br />
actives par le neurotransmetteur ( ex. perméabilité affecté par la phosphorylation)<br />
Expression pré<strong>synaptique</strong><br />
Expression post<strong>synaptique</strong>
<strong>Plasticité</strong> <strong>synaptique</strong> <strong>à</strong> <strong>long</strong> <strong>terme</strong><br />
Messagers rétrogrades impliqués dans l’expression pré<strong>synaptique</strong><br />
Augmentation de Ca 2+<br />
Induction post<strong>synaptique</strong><br />
de la plasticité<br />
Expression pré<strong>synaptique</strong><br />
de la plasticité<br />
Brain Derived Neurotropic Factor<br />
( BDNF)
<strong>Plasticité</strong> <strong>synaptique</strong> <strong>à</strong> <strong>long</strong> <strong>terme</strong><br />
Spike Timing Dependent Plasticity ( STDP)<br />
Postulat de Hebb: quand la décharge du neurone B suit la décharge du neurone A<br />
la connexion entre A et B est renforcée<br />
Stimulation électrique<br />
du neurone A<br />
Enregistrement et<br />
stimulation électrique du neurone B<br />
EPSP<br />
Markram et al 1997; Science<br />
Be & Poo 1998, J Neuroscience
<strong>Plasticité</strong> <strong>synaptique</strong> <strong>à</strong> <strong>long</strong> <strong>terme</strong><br />
Spike Timing Dependent Plasticity ( STDP)<br />
Postulat de Hebb: quand la décharge du neurone B suit la décharge du neurone A<br />
la connexion entre A et B est renforcée.<br />
Quand la décharge du neurone B précède celle du neurone A<br />
la connexion entre A et B est affaiblie.
Modification de la probabilité de libération du<br />
neurotransmetteur<br />
Modifications structurales<br />
Changement du nombre de connexion<br />
<strong>synaptique</strong>s<br />
<strong>Plasticité</strong> <strong>synaptique</strong> <strong>à</strong> <strong>long</strong> <strong>terme</strong><br />
Site d’expression<br />
Modification du nombre ou des propriétés des récepteurs<br />
activés par le neurotransmetteur ( ex. perméabilité affectée par phosphorylation)<br />
Expression pré<strong>synaptique</strong><br />
Expression post<strong>synaptique</strong>
<strong>Plasticité</strong> <strong>synaptique</strong> <strong>à</strong> <strong>long</strong> <strong>terme</strong><br />
Messagers rétrogrades impliqués dans l’expression pré<strong>synaptique</strong><br />
Augmentation de Ca 2+<br />
Induction post<strong>synaptique</strong><br />
de la plasticité<br />
Expression pré<strong>synaptique</strong><br />
de la plasticité<br />
Oxyde nitrique (NO)<br />
Brain Derived Neurotropic Factor<br />
( BDNF)<br />
Endocannabinoïdes
<strong>Plasticité</strong> <strong>synaptique</strong> <strong>à</strong> <strong>long</strong> <strong>terme</strong><br />
Rôle du BDNF<br />
Activité neuronale<br />
Libération de<br />
BDNF<br />
Le BDNF est une protéine (≈ 29 kDa) qui fait partie de la famille des neurotrophines
<strong>Plasticité</strong> <strong>synaptique</strong> <strong>à</strong> <strong>long</strong> <strong>terme</strong><br />
Rôle du BDNF<br />
Le BDNF est nécessaire pour l’expression de la LTP dans l’hippocampe<br />
Pang et al Science 2004
<strong>Plasticité</strong> <strong>synaptique</strong> <strong>à</strong> <strong>long</strong> <strong>terme</strong><br />
Rôle du BDNF<br />
L’infusion de BDNF induit une LTP in vivo<br />
Canula d’infusion<br />
D’âpres Bramham 2005
<strong>Plasticité</strong> <strong>synaptique</strong> <strong>à</strong> <strong>long</strong> <strong>terme</strong><br />
Rôle du BDNF<br />
Un déficit d’apprentissage est associé <strong>à</strong> une réduction de la libération de BDNF
2<br />
Potentiels d’action rétro-propagés<br />
3<br />
Activation des récepteurs<br />
métabotropiques ( mGluR, GABA B )<br />
<strong>Plasticité</strong> <strong>synaptique</strong> <strong>à</strong> <strong>long</strong> <strong>terme</strong><br />
Rôle du BDNF<br />
GABA B -R<br />
AMPA-R<br />
1<br />
Activation des récepteurs<br />
Glutamatergique<br />
( AMP, NMDA)
<strong>Plasticité</strong> <strong>synaptique</strong> <strong>à</strong> <strong>long</strong> <strong>terme</strong><br />
Rôle du BDNF<br />
Activité neuronale<br />
<strong>Plasticité</strong> <strong>synaptique</strong><br />
Libération de<br />
BDNF
Uncaging du glutamate sur une épine<br />
dendritique<br />
<strong>Plasticité</strong> <strong>synaptique</strong> <strong>à</strong> <strong>long</strong> <strong>terme</strong><br />
Rôle du BDNF<br />
L’activation concomitante d’une épine dendritique<br />
avec la génération de PA dans le neurone post<strong>synaptique</strong><br />
produit une augmentation du volume de l’épine<br />
( plasticité structurale)<br />
D’âpres Tanaka et al Science 2008
Uncaging du glutamate sur une épine<br />
dendritique<br />
<strong>Plasticité</strong> <strong>synaptique</strong> <strong>à</strong> <strong>long</strong> <strong>terme</strong><br />
Rôle du BDNF<br />
La plasticité structurale des épines est<br />
dépendante du BDNF<br />
K252a inhibiteur du récepteur du BDNF TrKB<br />
TrkB-FC immunoglobuline qui piège le BDNF<br />
extracellulaire<br />
D’âpres Tanaka et al Science 2008
BDNF<br />
Uncaging du glutamate sur une épine<br />
dendritique<br />
<strong>Plasticité</strong> <strong>synaptique</strong> <strong>à</strong> <strong>long</strong> <strong>terme</strong><br />
Rôle du BDNF<br />
La plasticité peut être obtenue en absence des<br />
PAs si du BDNF exogène est ajouté<br />
Asinomycin = inhibiteur de la synthèse protéique<br />
D’âpres Tanaka et al Science 2008
<strong>Plasticité</strong> <strong>synaptique</strong> <strong>à</strong> <strong>long</strong> <strong>terme</strong><br />
Rôle du BDNF<br />
Les potentiels d’action rétro-propagés déclenchent une libération de BDNF dendritique<br />
BDNF-GFP<br />
Time lapse fluorescence Imaging<br />
BDNF-GFP<br />
Control Post stimulation<br />
Neurone expriment le BDNF conjugué <strong>à</strong> une<br />
protéine fluorescente
<strong>Plasticité</strong> <strong>synaptique</strong> <strong>à</strong> <strong>long</strong> <strong>terme</strong><br />
Rôle du BDNF<br />
Le BDNF libéré par les PAs qui rétro- propagent dans les dendrites agit comme indicateur de coïncidence<br />
entre l’activité pré et post <strong>synaptique</strong>