Les acides gras Structure des acides gras - Biologie
Les acides gras Structure des acides gras - Biologie
Les acides gras Structure des acides gras - Biologie

Create successful ePaper yourself
Turn your PDF publications into a flip-book with our unique Google optimized e-Paper software.
<strong>Les</strong> <strong>aci<strong>des</strong></strong> <strong>gras</strong><br />
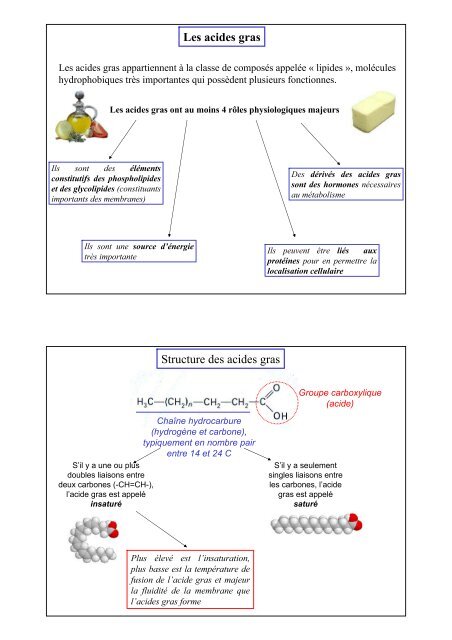
<strong>Les</strong> <strong>aci<strong>des</strong></strong> <strong>gras</strong> appartiennent à la classe de composés appelée « lipi<strong>des</strong> », molécules<br />
hydrophobiques très importantes qui possèdent plusieurs fonctionnes.<br />
<strong>Les</strong> <strong>aci<strong>des</strong></strong> <strong>gras</strong> ont au moins 4 rôles physiologiques majeurs<br />
Ils sont <strong>des</strong> éléments<br />
constitutifs <strong>des</strong> phospholipi<strong>des</strong><br />
et <strong>des</strong> glycolipi<strong>des</strong> (constituants<br />
importants <strong>des</strong> membranes)<br />
Des dérivés <strong>des</strong> <strong>aci<strong>des</strong></strong> <strong>gras</strong><br />
sont <strong>des</strong> hormones nécessaires<br />
au métabolisme<br />
Ils sont une source d’énergie<br />
très importante<br />
Ils peuvent être liés aux<br />
protéines pour en permettre la<br />
localisation cellulaire<br />
<strong>Structure</strong> <strong>des</strong> <strong>aci<strong>des</strong></strong> <strong>gras</strong><br />
Groupe carboxylique<br />
(acide)<br />
Chaîne hydrocarbure<br />
(hydrogène et carbone),<br />
typiquement en nombre pair<br />
entre 14 et 24 C<br />
S’il y a une ou plus<br />
doubles liaisons entre<br />
deux carbones (-CH=CH-),<br />
l’acide <strong>gras</strong> est appelé<br />
insaturé<br />
S’il y a seulement<br />
singles liaisons entre<br />
les carbones, l’acide<br />
<strong>gras</strong> est appelé<br />
saturé<br />
Plus élevé est l’insaturation,<br />
plus basse est la température de<br />
fusion de l’acide <strong>gras</strong> et majeur<br />
la fluidité de la membrane que<br />
l’<strong>aci<strong>des</strong></strong> <strong>gras</strong> forme
<strong>Les</strong> <strong>aci<strong>des</strong></strong> <strong>gras</strong> sont stockés surtout sous forme de triacylglycérol (esters non chargés<br />
du glycérol):<br />
Cellule adipose:<br />
-Cytoplasme<br />
-Noyau<br />
-Globule de<br />
stockage <strong>des</strong><br />
triacylglycérols<br />
<strong>Les</strong> carbones <strong>des</strong> <strong>aci<strong>des</strong></strong> <strong>gras</strong> sont très réduits, donc ils possèdent beaucoup d’énergie<br />
chimique. <strong>Les</strong> triacylglycérol sont donc <strong>des</strong> molécules de stockage de l’énergie très efficaces:<br />
leur oxydation donne environ 9 kcal/gramme, tandis que pour le sucres l’oxydation donne<br />
environ 4 kcal/gramme.<br />
Nomenclature <strong>des</strong> <strong>aci<strong>des</strong></strong> <strong>gras</strong><br />
Différents types de nomenclature:<br />
Numération <strong>des</strong> carbones à<br />
partir du groupe carboxylique<br />
<strong>Les</strong> carbones 2 et 3 e le dernier sont souvent<br />
appelés respectivement et (oméga)<br />
Exemple de nomenclature souvent utilisée:<br />
C18: 3 <br />
Nombre de carbones<br />
Nombre de double<br />
liaison<br />
Position de la dernière<br />
double liaison par rapport<br />
au carbone <br />
On peut trouver aussi l’indication, par exemple:<br />
cis- 9 ça signifie une double liaison avec configuration cis entre le carbone 9 et 10
Hydrolyses <strong>des</strong> triacylglycérols<br />
<strong>Les</strong> triacylglycérols sont la forme principale de stockage <strong>des</strong> <strong>aci<strong>des</strong></strong> <strong>gras</strong> (chez l’homme ils<br />
stockent environ 150 fois plus d’énergie que le glycogène), et pour les utiliser il faut hydrolyser<br />
les triacylglycérols accumulés dans les adipocytes ou assimilés.<br />
Des enzymes appelés lipase catalysent<br />
l’hydrolyse de la liaison ester entre<br />
<strong>aci<strong>des</strong></strong> <strong>gras</strong> et glycérol.<br />
Le glycérol peut être<br />
converti en DHAP et<br />
former glucose par la<br />
gluconéogenèse<br />
<strong>Les</strong> lipases <strong>des</strong> cellules adipeuses<br />
sont activées par phosphorylation<br />
induit par <strong>des</strong> hormones telles que le<br />
glucagon et l’adrénaline (et<br />
l’insuline a l’effet opposé)
Dégradation <strong>des</strong> <strong>aci<strong>des</strong></strong> <strong>gras</strong>: activation<br />
<strong>Les</strong> <strong>aci<strong>des</strong></strong> <strong>gras</strong> sont dégradés par oxydation au niveau du carbone ( oxydation).<br />
La première étape est l’activation <strong>des</strong> <strong>aci<strong>des</strong></strong> <strong>gras</strong> par liaison avec le Coenzyme A (en utilisant<br />
l’énergie de l’ATP):<br />
Acyl-CoA synthétase<br />
Acide <strong>gras</strong><br />
Coenzyme A<br />
Acyl-CoA<br />
pyrophosphate<br />
La réaction est rendue irréversible par l’hydrolyse rapide du pyrophosphate à 2 molécules de P i (déjà vu<br />
par l’enzyme UDP-glucose pyrophosphorylase pour la synthèse du glycogène)<br />
L’activation est effectuée dans la membrane mitochondriale externe, la dégradation (oxydation)<br />
dans la matrice mitochondriale:<br />
Activation<br />
Oxydation<br />
Le transport <strong>des</strong> acyl-CoA dans la<br />
matrice mitochondriale est facilité<br />
par une petite molécule appelée<br />
carnitine<br />
Dégradation <strong>des</strong> <strong>aci<strong>des</strong></strong> <strong>gras</strong>: oxydation<br />
Un acyl-CoA saturé est dégradé par une séquence récurrente de 4<br />
réactions:<br />
1) Oxydation par le FAD<br />
Résultat pour<br />
chaque cycle<br />
2) Hydratation<br />
3) Oxydation par le NAD +<br />
4) Thiolyse par le CoA<br />
-Formation d’un acyl-CoA plus court de 2 carbone.<br />
-Formation du FADH 2<br />
, NADH et acétyl-CoA<br />
(1)<br />
(2)<br />
Acyl-CoA déshydrogénase<br />
1) Acyl-CoA + E-FAD trans- 2 -énoyl-CoA + E-FADH 2<br />
énoyl-CoA hydratase<br />
2) trans- 2 -énoyl-CoA + H 2 O L-3-hydroxyacyl-CoA<br />
(3)<br />
hydroxyacyl-CoA déshydrogénase<br />
3) L-3-hydroxyacyl-CoA + NAD + 3-cétoacyl-CoA + NADH + H +<br />
-cétothiolase<br />
4) 3-cétoacyl-CoA (n carbones) + HS-CoA acétyl-CoA + acyl-CoA (n-2 carbones)<br />
(4)<br />
Remarquer que les étapes 1, 2 et 3 sont très similaires à celles-là du<br />
cycle de Krebs du succinate à l’oxaloacétate (étapes 6, 7 et 8).
<strong>Les</strong> acétyl-CoA produits par<br />
l’oxydation <strong>des</strong> <strong>aci<strong>des</strong></strong> <strong>gras</strong><br />
entrent dans le cycle de Krebs<br />
<strong>Les</strong> FADH 2 et les NADH produits sont oxydés par la chaîne de transport électronique<br />
mitochondriale pour permettre à la production d’ATP
Dégradation <strong>des</strong> <strong>aci<strong>des</strong></strong> <strong>gras</strong>: oxydation sur <strong>aci<strong>des</strong></strong> <strong>gras</strong> insaturés<br />
Pour la dégradation <strong>des</strong> <strong>aci<strong>des</strong></strong> <strong>gras</strong> insaturés (avec <strong>des</strong> double liaisons), deux enzymes<br />
supplémentaires sont nécessaires: une réductase et une isomérase.<br />
Cette intermédiaire ne peut pas être utilise pour les<br />
réactions suivantes d’oxydation<br />
La réductase (NADPH dépendent) réduit une double<br />
liaison et déplace l’autre en position 3.<br />
La double liaison (cis- 3 ) entre C3 et C4 empêche<br />
la formation de la double liaison entre C2 e C3<br />
(premier intermédiaire de la -oxydation)<br />
L’isomérase convertit la double liaison cis- 3 en trans- 2<br />
pour obtenir l’intermédiaire normal de la -oxydation<br />
Règle: les double liaison impair sont traitées par l’isomérase,<br />
tandis que les double liaison pair par la réductase e l’isomérase<br />
Synthèse <strong>des</strong> <strong>aci<strong>des</strong></strong> <strong>gras</strong><br />
La synthèse <strong>des</strong> <strong>aci<strong>des</strong></strong> <strong>gras</strong> est faite de 3 phases à partir de l’acétyl-CoA:<br />
1) Activation<br />
2) Élongation<br />
3) Terminaison<br />
Phase 1: Activation<br />
La synthèse <strong>des</strong> <strong>aci<strong>des</strong></strong> <strong>gras</strong> commence par la carboxylation de l’acétyl-CoA en malonyl-CoA<br />
Acétyl-CoA<br />
carboxylase<br />
Cette réaction irréversible est l’étape qui engage la synthèse <strong>des</strong> <strong>aci<strong>des</strong></strong> <strong>gras</strong>.<br />
Le cofacteur de la carboxylase est la biotine.
Acétyl-CoA<br />
carboxylase<br />
gluconéogenèse<br />
Pyruvate<br />
carboxylase<br />
Pyruvate<br />
Oxaloacétate<br />
Le cofacteur <strong>des</strong> deux carboxylases est la biotine<br />
Synthèse <strong>des</strong> <strong>aci<strong>des</strong></strong> <strong>gras</strong>: le complexe de l’acide <strong>gras</strong> synthase<br />
Chez les eucaryotes, un seul complexe multifonctionnel cytosolique catalyse toutes les<br />
réactions de synthèse de l’acide <strong>gras</strong><br />
Phase 2: Élongation<br />
La chaîne <strong>des</strong> acide <strong>gras</strong> en croissance est allongée par addition séquentielle d’unités à 2 carbones<br />
Cette phase commence par la formation d’acétyl-ACP et de malonyl-ACP.<br />
L’ACP (acyl carrier protein) a la fonction de transporter les unités acétyl; le groupe qui lie<br />
l’acétyl a la même structure que celui du Coenzyme A.<br />
Acétyl-CoA + ACP Acétyl-ACP + CoA<br />
Malonyl-CoA + ACP Malonyl-ACP + CoA
L’acide <strong>gras</strong> est allongé par une séquence récurrente de 4 réactions:<br />
1) Condensation<br />
2) Réduction par le NADPH<br />
3) Déshydratation<br />
4) Réduction par le NADPH<br />
Enzyme condensant<br />
acyl-malonyl-ACP<br />
1) Acétyl-ACP + malonyl-ACP acétoacétyl-ACP + ACP + CO 2<br />
La décarboxylation est nécessaire pour rendre exergonique la réaction<br />
de condensation.<br />
Le deuxième cycle et les suivants permettent l’allongement de<br />
la chaîne par la condensation du butyryl-ACP (au lieu de<br />
l’acétyl-ACP) avec une molécule de malonyl-ACP<br />
Phase 3: terminaison<br />
<strong>Les</strong> réactions d’allongement sont termines quand la chaîne est<br />
composée de 16 carbones. L’intermédiaire lié à l’enzyme est hydrolysé<br />
pour donner un acide <strong>gras</strong> C:16 (palmitate)<br />
Stœchiométrie total pour la synthèse du palmitate:<br />
8 acétyl-CoA + 7 ATP + 14 NADPH + 6 H + palmitate + 14 NADP + + 8 CoA + 6 H2O + 7 ADP + 7 P i<br />
Remarquer que la synthèse <strong>des</strong> <strong>aci<strong>des</strong></strong> <strong>gras</strong> est faite par une série de réactions de type contraire à<br />
celle de l’oxydation (mais évidemment les enzyme et les cofacteurs sont différents)<br />
-Oxydation<br />
1) Oxydation par le FAD<br />
2) Hydratation<br />
3) Oxydation par le NAD +<br />
4) Thiolyse par le CoA<br />
Oxydation<br />
•Mitochondrie<br />
•NADH, FADH 2<br />
•CoA<br />
•No CO 2<br />
•Enzymes séparés<br />
Différences<br />
Synthèse<br />
Synthèse<br />
1) Condensation<br />
2) Réduction par le NADPH<br />
3) Déshydratation<br />
4) Réduction par le NADPH<br />
•Cytosol<br />
•NADPH<br />
•Acyl carrier protéine (ACP)<br />
•CO 2 activation<br />
•Un seul complexe
Modification du palmitate<br />
Le palmitate, l’acide <strong>gras</strong> synthétisé par l’acide <strong>gras</strong> synthase, est un acide <strong>gras</strong> saturé à 16<br />
atomes de carbone, mais la cellule possède nombreux types d’acide <strong>gras</strong> .<br />
Des systèmes enzymatique accessoires catalysent l’élongation et la désaturation du palmitate<br />
pour former différents types <strong>des</strong> <strong>aci<strong>des</strong></strong> <strong>gras</strong>. Plusieurs enzymes qui catalysent ces réactions<br />
sont associés à la membrane cytosolique du réticulum endoplasmique (dans les plantes il y en a<br />
aussi dans le chloroplaste)<br />
IMPORTANT:<br />
<strong>Les</strong> mammifères n’ont pas la capacité<br />
d’introduire <strong>des</strong> double liaison entre<br />
atomes de carbone au-delà de C-9<br />
dans une chaîne d’<strong>aci<strong>des</strong></strong> <strong>gras</strong>.<br />
Il faut donc assimiler certains <strong>aci<strong>des</strong></strong><br />
<strong>gras</strong>, qui sont donc appelés essentiels<br />
(par ex. ceux-là connus comme 3 et<br />
6)<br />
Source du NADPH<br />
Le NADPH utilisé par le synthèse <strong>des</strong> <strong>aci<strong>des</strong></strong> <strong>gras</strong> (voie anabolique) provient de deux sources<br />
principales:<br />
-NADPH produit par la voie <strong>des</strong> pentoses phosphates<br />
-NADPH produit par l’enzyme malique<br />
Matrice mitochondriale<br />
ATP + CoA<br />
Cytosol<br />
citrate<br />
CoA<br />
Acétyl-CoA<br />
oxaloacétate<br />
ADP + P i + 2 H +<br />
citrate<br />
ATP-citrate<br />
lyase<br />
Malate<br />
déshydrogénase<br />
cytosolique<br />
Enzyme<br />
malique<br />
Acétyl-CoA + ADP + P i<br />
oxaloacétate<br />
malate<br />
NADH + H +<br />
NAD +<br />
NADP +<br />
ATP + H 2 O<br />
Pyruvate + CO 2<br />
Pyruvate + CO 2<br />
NADPH<br />
Le transport de l’acétyl-CoA de la matrice mitochondriale au cytosol est réalisé par formation du citrate;<br />
dans le cytosol le citrate est puis retransformé en oxaloacétate et acétyl-CoA; la malate déshydrogénase<br />
cytosolique et l’enzyme malique permettent de obtenir du NADPH à partir du NADH
Régulation de la synthèse <strong>des</strong> <strong>aci<strong>des</strong></strong> <strong>gras</strong><br />
Le synthèse <strong>des</strong> <strong>aci<strong>des</strong></strong> <strong>gras</strong> est faite lorsque les gluci<strong>des</strong> et l’énergie sont abondants, et<br />
lorsque les <strong>aci<strong>des</strong></strong> <strong>gras</strong> sont rares.<br />
L’acétyl-CoA carboxylase est l’enzyme clé pour le control de la voie (sous contrôle<br />
allostérique et par modification covalente):<br />
L’acétyl-CoA carboxylase est inhibée par phosphorylation par<br />
une kinase AMP dépendante; la phosphatase (2A), par contre, est<br />
sous contrôle hormonale (désactivée par le glucagon)<br />
<strong>Les</strong> phospholipi<strong>des</strong> sont <strong>des</strong> constituants de membrane qui contiennent <strong>des</strong> <strong>aci<strong>des</strong></strong> <strong>gras</strong><br />
Tête polaire<br />
queue apolaire<br />
La présence <strong>des</strong> <strong>aci<strong>des</strong></strong> <strong>gras</strong> insaturés<br />
rompt l’empilement <strong>des</strong> chaînes<br />
hydrophobiques et rend la membrane<br />
plus fluide.
L’organisation <strong>des</strong> lipi<strong>des</strong> dans la<br />
membrane<br />
<strong>Les</strong> doubles liaisons cis dans les<br />
<strong>aci<strong>des</strong></strong> <strong>gras</strong> sont capable<br />
d’augmenter la fluidité <strong>des</strong><br />
membranes en diminuant la<br />
température de fusion.<br />
Utilisation <strong>des</strong> différentes sources d’énergie pendant les activités musculaires<br />
La créatine-phosphate est un<br />
réservoir de liaisons ~P qui<br />
permet la phosphorylation de<br />
l’ATP très rapidement<br />
-ATP 1 seconde<br />
- Créatine-P 5 seconde<br />
- Glycolyse anaérobie dés quelques secon<strong>des</strong> à quelques minutes<br />
- Glycolyse aérobie dés quelques secon<strong>des</strong> à plusieurs heures<br />
- Lipolyse (<strong>aci<strong>des</strong></strong> <strong>gras</strong>) après 15-30 minutes pour plusieurs heures
Stockage de l’énergie (homme de 70 kg):<br />
Glucose libre<br />
40 kcal<br />
Glycogène<br />
600 kcal<br />
(Protéines)<br />
25000 kcal<br />
Triacylglycérol (<strong>aci<strong>des</strong></strong> <strong>gras</strong>) 100000 kcal<br />
Sport différents utilisent <strong>des</strong> sources d’énergie différentes<br />
Exemples:<br />
Sprint sur 100 mètres 10 secon<strong>des</strong><br />
ATP<br />
(1 s)<br />
Créatine-P<br />
(4 s)<br />
glycolyse anaérobie du glucose<br />
(quelques secon<strong>des</strong>)<br />
L’ATP, la créatine et le glucose sont la principal source d’énergie pour <strong>des</strong> réactions anaérobies
Course 5 km 15 minutes<br />
glycolyse aérobie du glucose libre<br />
(peu de minutes)<br />
glycolyse aérobie du glycogène<br />
(plusieurs minutes)<br />
Le glycogène est la principal source d’énergie à travers la glycolyse aérobie<br />
Course longue (marathon, cyclisme) 2-6 heures<br />
Il faut utiliser les <strong>aci<strong>des</strong></strong> <strong>gras</strong> parce que l'énergie stockée sous forme de glycogène n’est pas<br />
suffisante pour <strong>des</strong> activités de plusieurs heures<br />
glycolyse aérobie du glycogène<br />
(heures)<br />
Lipolyse <strong>des</strong> triacylglycérols et<br />
oxydation <strong>aci<strong>des</strong></strong> <strong>gras</strong><br />
(plusieurs heures)<br />
L’organisme maintient le taux du glucose faible dans le sang rapport glucagon/insuline élevé<br />
activation lipolyse l’acétyl-CoA produit par oxydation <strong>des</strong> <strong>aci<strong>des</strong></strong> <strong>gras</strong> ralentie la pyruvate<br />
déshydrogénase et donc la glycolyse le glucose (glycogène) est économisé et utilisé pour<br />
plusieurs heures et les <strong>aci<strong>des</strong></strong> <strong>gras</strong> deviennent la principale source d’énergie<br />
<strong>Les</strong> corps cétoniques<br />
transporté<br />
Une série de réactions (qui se déroulent surtout dans le foie) permet la<br />
production de molécules appelées « corps cétoniques » à partir de<br />
l’acétyl-CoA.<br />
L’acétoacétate est une importantes source d’énergie, en condition<br />
normale surtout pour le muscle cardiaque, et en condition de jeûne<br />
prolongé aussi pour le cerveau (qui normalement utilise le glucose).<br />
L’acétoacétate produit dans le foie est transporté par le sang vers les<br />
tissus périphériques qui le retransforment en Acétyl-CoA qui sera<br />
oxydé par le cycle de Krebs.<br />
Au cycle de Krebs