

Mécano-sensibilité cellulaire : adaptation physique à la rigidité
Mécano-sensibilité cellulaire : adaptation physique à la rigidité
Mécano-sensibilité cellulaire : adaptation physique à la rigidité

You also want an ePaper? Increase the reach of your titles
YUMPU automatically turns print PDFs into web optimized ePapers that Google loves.
<strong>Mécano</strong>-<strong>sensibilité</strong> <strong>cellu<strong>la</strong>ire</strong> : <strong>adaptation</strong> <strong>physique</strong> <strong>à</strong> <strong>la</strong> <strong>rigidité</strong><br />
0,02<br />
0,5<br />
1,2<br />
0,1<br />
Vitesse de contraction (µm/s)<br />
0,015<br />
0,01<br />
0,005<br />
0,4<br />
0,3<br />
0,2<br />
0,1<br />
Puissance mécanique (f W)<br />
V/V max<br />
1<br />
0,8<br />
0,6<br />
0,4<br />
0,2<br />
0,08<br />
0,06<br />
0,04<br />
0,02<br />
P/F max V max<br />
0<br />
0<br />
0<br />
50 100 150 200<br />
0<br />
0<br />
Charge (nN)<br />
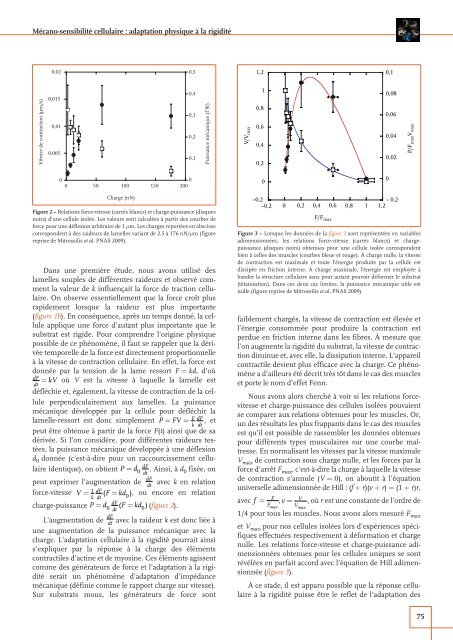
Figure 2 – Re<strong>la</strong>tions force-vitesse (carrés b<strong>la</strong>ncs) et charge-puissance (disques<br />
noirs) d’une cellule isolée. Les valeurs sont calculées <strong>à</strong> partir des courbes de<br />
force pour une déflexion arbitraire de 1 μm. Les charges reportées en abscisse<br />
correspondent <strong>à</strong> des raideurs de <strong>la</strong>melles variant de 2,5 <strong>à</strong> 176 nN/μm (figure<br />
reprise de Mitrossilis et al. PNAS 2009).<br />
Dans une première étude, nous avons utilisé des<br />
<strong>la</strong>melles souples de différentes raideurs et observé comment<br />
<strong>la</strong> valeur de k influençait <strong>la</strong> force de traction <strong>cellu<strong>la</strong>ire</strong>.<br />
On observe essentiellement que <strong>la</strong> force croît plus<br />
rapidement lorsque <strong>la</strong> raideur est plus importante<br />
(figure 1b). En conséquence, après un temps donné, <strong>la</strong> cellule<br />
applique une force d’autant plus importante que le<br />
substrat est rigide. Pour comprendre l’origine <strong>physique</strong><br />
possible de ce phénomène, il faut se rappeler que <strong>la</strong> dérivée<br />
temporelle de <strong>la</strong> force est directement proportionnelle<br />
<strong>à</strong> <strong>la</strong> vitesse de contraction <strong>cellu<strong>la</strong>ire</strong>. En effet, <strong>la</strong> force est<br />
donnée par <strong>la</strong> tension de <strong>la</strong> <strong>la</strong>me ressort F = kd, d’où<br />
dF<br />
= kV où V est <strong>la</strong> vitesse <strong>à</strong> <strong>la</strong>quelle <strong>la</strong> <strong>la</strong>melle est<br />
dt<br />
défléchie et, également, <strong>la</strong> vitesse de contraction de <strong>la</strong> cellule<br />
perpendicu<strong>la</strong>irement aux <strong>la</strong>melles. La puissance<br />
mécanique développée par <strong>la</strong> cellule pour défléchir <strong>la</strong><br />
<strong>la</strong>melle-ressort est donc simplement P= FV =<br />
F dF<br />
, et<br />
k dt<br />
peut être obtenue <strong>à</strong> partir de <strong>la</strong> force F(t) ainsi que de sa<br />
dérivée. Si l’on considère, pour différentes raideurs testées,<br />
<strong>la</strong> puissance mécanique développée <strong>à</strong> une déflexion<br />
d 0 donnée (c’est-<strong>à</strong>-dire pour un raccourcissement <strong>cellu<strong>la</strong>ire</strong><br />
identique), on obtient P= 0 . Ainsi, <strong>à</strong> d 0 fixée, on<br />
peut exprimer l’augmentation de dF avec k en re<strong>la</strong>tion<br />
dt<br />
force-vitesse V =<br />
1 dF<br />
( F=<br />
kd<br />
k dt 0), ou encore en re<strong>la</strong>tion<br />
charge-puissance P= d dF ( F=<br />
kd )(figure 2).<br />
d dF<br />
dt<br />
0 dt 0<br />
L’augmentation de dF<br />
dt<br />
avec <strong>la</strong> raideur k est donc liée <strong>à</strong><br />
une augmentation de <strong>la</strong> puissance mécanique avec <strong>la</strong><br />
charge. L’<strong>adaptation</strong> <strong>cellu<strong>la</strong>ire</strong> <strong>à</strong> <strong>la</strong> <strong>rigidité</strong> pourrait ainsi<br />
s’expliquer par <strong>la</strong> réponse <strong>à</strong> <strong>la</strong> charge des éléments<br />
contractiles d’actine et de myosine. Ces éléments agissent<br />
comme des générateurs de force et l’<strong>adaptation</strong> <strong>à</strong> <strong>la</strong> <strong>rigidité</strong><br />
serait un phénomène d’<strong>adaptation</strong> d’impédance<br />
mécanique (définie comme le rapport charge sur vitesse).<br />
Sur substrats mous, les générateurs de force sont<br />
–0,2<br />
–0,2<br />
faiblement chargés, <strong>la</strong> vitesse de contraction est élevée et<br />
l’énergie consommée pour produire <strong>la</strong> contraction est<br />
perdue en friction interne dans les fibres. <strong>à</strong> mesure que<br />
l’on augmente <strong>la</strong> <strong>rigidité</strong> du substrat, <strong>la</strong> vitesse de contraction<br />
diminue et, avec elle, <strong>la</strong> dissipation interne. L’appareil<br />
contractile devient plus efficace avec <strong>la</strong> charge. Ce phénomène<br />
a d’ailleurs été décrit très tôt dans le cas des muscles<br />
et porte le nom d’effet Fenn.<br />
Nous avons alors cherché <strong>à</strong> voir si les re<strong>la</strong>tions forcevitesse<br />
et charge-puissance des cellules isolées pouvaient<br />
se comparer aux re<strong>la</strong>tions obtenues pour les muscles. Or,<br />
un des résultats les plus frappants dans le cas des muscles<br />
est qu’il est possible de rassembler les données obtenues<br />
pour différents types muscu<strong>la</strong>ires sur une courbe maîtresse.<br />
En normalisant les vitesses par <strong>la</strong> vitesse maximale<br />
V max de contraction sous charge nulle, et les forces par <strong>la</strong><br />
force d’arrêt F max , c’est-<strong>à</strong>-dire <strong>la</strong> charge <strong>à</strong> <strong>la</strong>quelle <strong>la</strong> vitesse<br />
de contraction s’annule (V = 0), on aboutit <strong>à</strong> l’équation<br />
universelle adimensionnée de Hill : (f + r)(v + r) = (1 + r)r,<br />
F<br />
F max<br />
0<br />
0,2<br />
0,4<br />
F/F max<br />
0,6<br />
– 0,2<br />
1,2<br />
Figure 3 – Lorsque les données de <strong>la</strong> figure 2 sont représentées en variables<br />
adimensionnées, les re<strong>la</strong>tions force-vitesse (carrés b<strong>la</strong>ncs) et chargepuissance<br />
(disques noirs) obtenues pour une cellule isolée correspondent<br />
bien <strong>à</strong> celles des muscles (courbes bleue et rouge). <strong>à</strong> charge nulle, <strong>la</strong> vitesse<br />
de contraction est maximale et toute l’énergie produite par <strong>la</strong> cellule est<br />
dissipée en friction interne. <strong>à</strong> charge maximale, l’énergie est employée <strong>à</strong><br />
bander <strong>la</strong> structure <strong>cellu<strong>la</strong>ire</strong> sans pour autant pouvoir déformer le substrat<br />
(tétanisation). Dans ces deux cas limites, <strong>la</strong> puissance mécanique utile est<br />
nulle (figure reprise de Mitrossilis et al. PNAS 2009).<br />
avec f = , v =<br />
V<br />
, où r est une constante de l’ordre de<br />
V max<br />
1/4 pour tous les muscles. Nous avons alors mesuré F max<br />
et V max pour nos cellules isolées lors d’expériences spécifiques<br />
effectuées respectivement <strong>à</strong> déformation et charge<br />
nulle. Les re<strong>la</strong>tions force-vitesse et charge-puissance adimensionnées<br />
obtenues pour les cellules uniques se sont<br />
révélées en parfait accord avec l’équation de Hill adimensionnée<br />
(figure 3).<br />
<strong>à</strong> ce stade, il est apparu possible que <strong>la</strong> réponse <strong>cellu<strong>la</strong>ire</strong><br />
<strong>à</strong> <strong>la</strong> <strong>rigidité</strong> puisse être le reflet de l’<strong>adaptation</strong> des<br />
0,8<br />
1<br />
75













