anatomia del neurone - CPRG
anatomia del neurone - CPRG
anatomia del neurone - CPRG

Create successful ePaper yourself
Turn your PDF publications into a flip-book with our unique Google optimized e-Paper software.
la costruzione<br />
<strong>del</strong> <strong>neurone</strong><br />
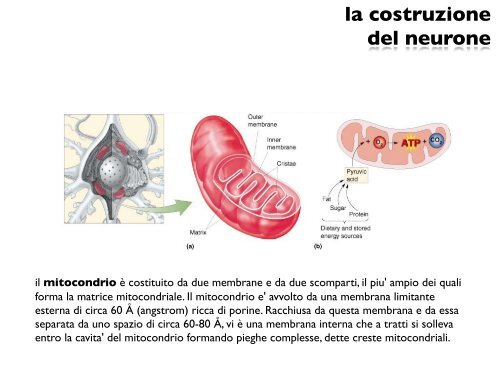
il mitocondrio è costituito da due membrane e da due scomparti, il piu' ampio dei quali<br />
forma la matrice mitocondriale. Il mitocondrio e' avvolto da una membrana limitante<br />
esterna di circa 60 Å (angstrom) ricca di porine. Racchiusa da questa membrana e da essa<br />
separata da uno spazio di circa 60-80 Å, vi è una membrana interna che a tratti si solleva<br />
entro la cavita' <strong>del</strong> mitocondrio formando pieghe complesse, dette creste mitocondriali.
la costruzione<br />
<strong>del</strong> <strong>neurone</strong><br />
La membrana interna divide il mitocondrio in due scomparti: la camera esterna compresa<br />
tra le due membrane e che occupa la parte centrale <strong>del</strong>le creste, e la camera interna<br />
limitata dalla membrana interna. La camera interna contiene un materiale detto matrice<br />
mitocondriale (ricca di ribosomi). Le creste mitocondriali formate dalle estroflessioni<br />
<strong>del</strong>la membrana interna, costituiscono <strong>del</strong>le pieghe che non interrompono la continuità<br />
<strong>del</strong>la camera interna.
la costruzione<br />
<strong>del</strong> <strong>neurone</strong><br />
il mitocondrio è la sede <strong>del</strong>la respirazione cellulare<br />
nell’organulo entrano grassi, zuccheri e proteine sotto forma di acido piruvico oltre ad<br />
ossigeno
la costruzione<br />
<strong>del</strong> <strong>neurone</strong><br />
l’acido piruvico è sottoposto ad una serie di reazioni biochimiche (ciclo di Krebs)<br />
si ha l’aggiunta di un gruppo fosfato all’adenindifosfato (ADP) dando vita<br />
all’adenintrifosfato (ATP)<br />
per ogni molecola di acido piruvico vengono prodotte 17 molecole di ATP ovvero la<br />
benzina <strong>del</strong>la cellula nervosa
<strong>anatomia</strong> <strong>del</strong><br />
<strong>neurone</strong><br />
la membrana neuronale<br />
è una barriera che si frappone per contenere il citoplasma all’interno <strong>del</strong> <strong>neurone</strong>, esclude<br />
sostanze dannose <strong>del</strong> fluido extracellulare.<br />
ha spessore di circa 5nm ed è costellata di proteine le quali svolgono un’azione di<br />
pompaggio attivo di sostanze dentro e fuori la cellula. altre proteine invece sono selettive<br />
per il passaggio di determinate sostanze.<br />
le proteine di membrana <strong>del</strong> <strong>neurone</strong> sono specifiche per il distretto d’interesse: quelle<br />
<strong>del</strong> soma saranno diverse da quelle dei dendriti<br />
non è possibile comprendere il funzionamento dei neuroni prescindendo dalle proprietà<br />
morfologiche ed elettriche <strong>del</strong>le diverse membrane
<strong>anatomia</strong> <strong>del</strong><br />
<strong>neurone</strong><br />
membrana neuronale<br />
la struttura sulla quale poggia la membrana neuronale è il citoscheletro<br />
i componenti fondamentali <strong>del</strong> citoscheletro sono:<br />
microtubuli<br />
microfilamenti<br />
neurofilamenti
<strong>anatomia</strong> <strong>del</strong><br />
<strong>neurone</strong><br />
membrana neuronale
<strong>anatomia</strong> <strong>del</strong><br />
<strong>neurone</strong><br />
membrana neuronale<br />
Citoscheletro Statico (microtubuli e neurofilamenti)<br />
Citoscheletro Dinamico (microfilamenti e proteine contrattili associate)<br />
microtubuli<br />
Sono organuli citoplasmatici presenti in tutte le cellule. Appaiono al<br />
microscopio come strutture cilindriche cave, con un diametro di 25 nm ed<br />
uno interno di 15 nm.<br />
In sezione longitudinale i microtubuli appaiono come bastoncini di lunghezza<br />
variabile che può raggiungere 20-60 μm.<br />
La parete dei microtubuli è composta da una serie di unità sferoidali ordinate<br />
rigidamente di 4 nm. Ogni subunità corrisponde ad una molecola di<br />
tubulina
<strong>anatomia</strong> <strong>del</strong><br />
microtubuli<br />
La tubulina è un dimero (molecola formata da 2<br />
subunità) formato da subunità di sequenza<br />
amminoacidica simile, chiamate tubulina α e tubulina ß.<br />
<strong>neurone</strong><br />
membrana neuronale<br />
I dimeri di tubulina polimerizzano a formare lunghe<br />
catene chiamate protofilamenti<br />
Nella cellula i protofilamenti sono assemblati a gruppi<br />
di tredici in una struttura che nel complesso forma il<br />
microtubulo<br />
I protofilamenti si avvolgono a spirale e decorrono<br />
paralleli tra di loro intorno all'asse <strong>del</strong> microtubulo
<strong>anatomia</strong> <strong>del</strong><br />
microtubuli<br />
La vita media <strong>del</strong>la tubulina è di circa un giorno.<br />
La vita media di un microtubulo è di soli 10 minuti<br />
<strong>neurone</strong><br />
membrana neuronale<br />
Sono in continuo stato di assemblaggio e<br />
disassemblaggio. Questa caratteristica è detta<br />
“instabilità dinamica”<br />
La crescita dei microtubuli è ovviamente influenzata da<br />
molti fattori quali ad esempio la divisione cellulare e il<br />
movimento.<br />
Il movimento <strong>del</strong>le vescicole o degli organuli cellulari<br />
all’interno <strong>del</strong>la cellula dipende dai microtubuli e dalle<br />
proteine ad essi associate (MAP)
<strong>anatomia</strong> <strong>del</strong><br />
microtubuli<br />
Sono proteine definite come MAP (microtubules associated<br />
proteins) o proteine associate ai microtubuli<br />
<strong>neurone</strong><br />
membrana neuronale<br />
Le MAP arrivano a costituire il 20 % <strong>del</strong>la massa totale<br />
Le MAP appartengono a due classi di proteine:<br />
le MAP motrici, e le MAP non motrici<br />
Le MAP motrici comprendono la chinesina e la dineina<br />
le MAP non motrici sono in grado di coordinare<br />
l’organizzazione dei microtubuli nel citoplasma
<strong>anatomia</strong> <strong>del</strong><br />
<strong>neurone</strong><br />
membrana neuronale
<strong>anatomia</strong> <strong>del</strong><br />
<strong>neurone</strong><br />
membrana neuronale<br />
microfilamenti<br />
formati dall’assemblaggio di molte unità <strong>del</strong>la<br />
proteina actina<br />
Sono strutture in grado di assemblarsi e<br />
disassemblarsi<br />
Sebbene presenti in tutta la cellula, si<br />
concentrano nella zona sottostante alla<br />
membrana plasmatica<br />
connettono complessi proteici ed organelli in<br />
regioni diverse <strong>del</strong>la cellula<br />
hanno la funzione di binari per lo spostamento<br />
degli organuli ad essi ancorati
<strong>anatomia</strong> <strong>del</strong><br />
<strong>neurone</strong><br />
membrana neuronale<br />
neurofilamenti<br />
dimensioni intermedie (Ø 8-12 nm) rispetto a<br />
microfilamenti e microtubuli<br />
Sono costituiti da vari tipi di proteine, tra cui la<br />
cheratina<br />
Formano una solida rete nel citoplasma<br />
Si ancorano alla membrana plasmatica al livello<br />
<strong>del</strong>le giunzioni tra cellule adiacenti<br />
Sono le strutture più stabili <strong>del</strong> citoscheletro
<strong>anatomia</strong> <strong>del</strong><br />
<strong>neurone</strong><br />
membrana neuronale<br />
ricapitolando:<br />
Neurofilamenti<br />
(cheratine)<br />
Microtubuli<br />
(α e β<br />
tubulina)<br />
Microfilamenti<br />
(actina)
<strong>anatomia</strong> <strong>del</strong><br />
ricapitolando:<br />
Mictrotubuli (Ø 20 nm)<br />
Motilità cellulare<br />
Forma <strong>del</strong>la cellula<br />
Movimento dei cromosomi e degli organuli<br />
Trasporto assonale<br />
<strong>neurone</strong><br />
membrana neuronale<br />
Microfilamenti (Ø 5 nm)<br />
Scorrimento <strong>del</strong> citoplasma<br />
Movimento ameboide<br />
Divisione cellulare<br />
Forma <strong>del</strong>la cellula<br />
Neurofilamenti (Ø 8-12 nm)<br />
(altrove filamenti intermedi)<br />
Supporto strutturale<br />
Forma <strong>del</strong>la cellula
<strong>neurone</strong><br />
membrana neuronale<br />
ricapitolando:<br />
il citoscheletro da forma e resistenza meccanica alla cellula<br />
raccordandosi con la membrana plasmatica e il sistema <strong>del</strong>le membrane<br />
Mantiene nella giusta posizione gli organuli citoplasmatici<br />
Alcuni elementi <strong>del</strong> citoscheletro fungono da vie per il trasporto di<br />
molecole e organuli<br />
È implicato nel movimento <strong>del</strong> citoplasma e di ciglia e flagelli<br />
Svolge un ruolo chiave nella divisione cellulare<br />
inoltre:<br />
In alcuni processi degenerativi, come ad esempio nella demenza di<br />
Alzheimer, gli elementi <strong>del</strong> citoscheletro proliferano fino a far morire la<br />
cellula e poi rimangono a lungo dopo la morte cellulare (la cosiddetta<br />
placca di neurofibrilla)<br />
<strong>anatomia</strong> <strong>del</strong>
<strong>anatomia</strong> <strong>del</strong><br />
<strong>neurone</strong><br />
membrana neuronale<br />
un’ultima funzione importante:<br />
Nel <strong>neurone</strong> porzioni diverse <strong>del</strong>la membrana plasmatica<br />
(ad es quella dei dendriti) hanno proprietà molto diverse<br />
e svolgono funzioni molto specializzate (determinate dalla<br />
diversa composizione nelle proteine di membrana)<br />
In assenza di altri fattori la composizione <strong>del</strong>le membrane<br />
tenderebbe ad uniformarsi<br />
Il citoscheletro impedisce che questo avvenga ancorando<br />
le proteine di membrana in una certa parte <strong>del</strong>la cellula
<strong>anatomia</strong> <strong>del</strong><br />
l’assone è una caratteristica esclusiva<br />
dei neuroni<br />
<strong>neurone</strong><br />
assone<br />
il punto d’inizio <strong>del</strong>l’assone è il cono di<br />
integrazione<br />
gli assoni sono solitamente privi di RER<br />
la composizione <strong>del</strong>la membrana<br />
assonica è diversa da quella <strong>del</strong> soma<br />
non c’è sintesi proteica a livello<br />
<strong>del</strong>l’assone quindi le proteine devono<br />
essere sintetizzate nel soma<br />
le ramificazioni degli assoni sono dette<br />
assoni collaterali e quando tornano<br />
sull’assone principale vengono detti<br />
ricorrenti<br />
grande variabilità in lunghezza e<br />
diametro (da 1micron a 1mm): effetti<br />
sulla velocità di trasmissione
<strong>anatomia</strong> <strong>del</strong><br />
il punto d’inizio <strong>del</strong>l’assone è il cono di integrazione<br />
mentre la parte finale è detta terminale assonico o<br />
bottone terminale<br />
<strong>neurone</strong><br />
assone
<strong>anatomia</strong> <strong>del</strong><br />
a livello <strong>del</strong> terminale assonico avviene la sinapsi<br />
<strong>neurone</strong><br />
assone<br />
a volte il terminale assonico è molto ramificato, un<br />
processo denominato arborizzazione dendritica<br />
ciascuna ramificazione fa sinapsi con dentriti o corpi<br />
cellulari adiacenti<br />
i bottoni sinaptici<br />
sono rigonfiamenti assonici<br />
sui quali avviene una<br />
sinapsi. questo particolare<br />
tipo di comunicazione è<br />
detta innervazione
<strong>anatomia</strong> <strong>del</strong><br />
anche il citoplasma <strong>del</strong> terminale assonico<br />
differisce da quello <strong>del</strong> corpo cellulare:<br />
<strong>neurone</strong><br />
assone<br />
-i microtubuli non vi arrivano <strong>del</strong><br />
terminale assonico avviene la sinapsi<br />
-è ricco di vescicole<br />
sinaptiche (50nm)<br />
-la parte interna <strong>del</strong>la<br />
membrana è ricca di<br />
proteine<br />
-grande concentrazione di<br />
mitocondri (grande richiesta<br />
energetica)
<strong>anatomia</strong> <strong>del</strong><br />
<strong>neurone</strong><br />
assone
<strong>anatomia</strong> <strong>del</strong><br />
<strong>neurone</strong><br />
assone<br />
non c’è sintesi proteica a livello <strong>del</strong>l’assone<br />
se separato dal resto <strong>del</strong>la cellula l’assone<br />
degenera (degenerazione walleriana) in<br />
quanto mancano i costituenti cellulari<br />
è necessario che il materiale arrivi dal soma<br />
fino alla parte terminale <strong>del</strong>l’assone<br />
attraverso un processo chiamato<br />
trasporto assoplasmatico
<strong>anatomia</strong> <strong>del</strong><br />
<strong>neurone</strong><br />
assone<br />
Anterogrado rapido (410 mm/giorno)<br />
Trasporta organuli e vescicole<br />
Microtubuli +<br />
chinesina<br />
Retrogrado rapido (200-300 mm/giorno)<br />
Trasporta organuli e vescicole<br />
Microtubuli +<br />
dineina<br />
Assonale lento – componente lenta (2,5 mm/giorno)<br />
– componente veloce (5 mm/giorno)<br />
Trasporta enzimi e altre macromoecole
<strong>anatomia</strong> <strong>del</strong><br />
<strong>neurone</strong><br />
assone<br />
Nel 1972 Ochs dimostrò che il trasporto<br />
assonale rapido avviene alla velocità di 410<br />
mm al giorno<br />
La sua verifica era basata sulla marcatura<br />
radiottiva <strong>del</strong>le proteine <strong>del</strong> soma di<br />
motoneuroni che appartenevano al nervo<br />
sciatico e sulla misura <strong>del</strong> fronte di<br />
avanzamento <strong>del</strong>la radioattività lungo gli<br />
assoni in tempi successivi
<strong>anatomia</strong> <strong>del</strong><br />
<strong>neurone</strong><br />
assone<br />
Il trasporto retrogrado rapido può essere utilizzato dai virus per<br />
infettare i neuroni e per penetrare la barriera emato-encefalica.<br />
Questo metodo viene ad esempio utilizzato dal virus <strong>del</strong>l’erpes e<br />
da quello <strong>del</strong>la rabbia<br />
Il trasporto retrogrado viene anche utilizzato dai ricercatori per<br />
tracciare le connessioni tra neuroni
<strong>anatomia</strong> <strong>del</strong><br />
<strong>neurone</strong><br />
assone<br />
Due MAP motrici sono la chinesina e la dineina, due proteine<br />
che fanno da ponte fra i microtubuli e le vescicole intracellulari.<br />
La chinesina e la dineina sono capaci di muoversi sui microtubuli<br />
che agiscono da binario in direzioni opposte, trasportando le<br />
vescicole intracellulari.
<strong>anatomia</strong> <strong>del</strong><br />
<strong>neurone</strong><br />
assone<br />
sono presenti <strong>del</strong>le teste che si<br />
legano ai microtubuli e all’ATP.<br />
Questo tipo di porzione di<br />
proteina detta testa si definisce<br />
anche “Motore ad ATPasi”<br />
poiché è il legame e l’idrolisi<br />
<strong>del</strong>l’ATP che permette il<br />
movimento.<br />
La zona <strong>del</strong>la coda si lega agli<br />
organuli cellulari o alle<br />
vescicole da spostare.
<strong>anatomia</strong> <strong>del</strong><br />
<strong>neurone</strong><br />
assone<br />
La chinesina si muove verso la porzione positiva (testa<br />
<strong>del</strong>l’assone) mentre la dineina verso quella negativa (coda<br />
<strong>del</strong>l’assone).