

Witte, F., Rutjes, C., Wanink, J. & van den Thillart, G. (2005)
Witte, F., Rutjes, C., Wanink, J. & van den Thillart, G. (2005)
Witte, F., Rutjes, C., Wanink, J. & van den Thillart, G. (2005)

Create successful ePaper yourself
Turn your PDF publications into a flip-book with our unique Google optimized e-Paper software.
EEN ADEMBENEMEND LEVEN:<br />
CICHLIDEN EN HUN AANPASSINGEN AAN LAGE<br />
ZUURSTOFCONCENTRATIES<br />
Tekst en foto’s: Frans <strong>Witte</strong>, Carlo <strong>Rutjes</strong>, Jan <strong>Wanink</strong> & Guido <strong>van</strong> <strong>den</strong> <strong>Thillart</strong><br />
Sectie Integratieve Zoologie, Instituut Biologie Lei<strong>den</strong><br />
Inleiding<br />
De cichli<strong>den</strong> uit de Oost-Afrikaanse meren zijn<br />
bekend om hun soortenrijkdom, snelle evolutie<br />
en grote verschei<strong>den</strong>heid <strong>van</strong> aanpassingen,<br />
waar<strong>van</strong> vooral die aan verschillende voedseltypen<br />
opvalt. Er zijn echter ook andere milieufactoren<br />
waaraan cichli<strong>den</strong> een scala <strong>van</strong> aanpassingen<br />
vertonen, bijvoorbeeld aan lichtomstandighe<strong>den</strong><br />
en aan zuurstofconcentraties.<br />
Licht en ogen zijn in een vorig nummer <strong>van</strong> dit<br />
tijdschrift aan de orde gekomen (<strong>Witte</strong> et al.,<br />
2003). In dit artikel bespreken we de eigenschappen<br />
<strong>van</strong> zuurstof in water, de bouw<br />
en werking <strong>van</strong> vissenkieuwen, en de<br />
aanpassingen bij cichli<strong>den</strong> die onder<br />
lage zuurstofconcentraties leven.<br />
Zuurstof in water<br />
In water zit ongeveer 30 maal minder<br />
zuurstof dan in lucht. De hoeveelheid<br />
wordt uitgedrukt in milligram<br />
per liter (mg/l) en is afhankelijk<br />
<strong>van</strong> de temperatuur <strong>van</strong> het<br />
water en <strong>van</strong> de hoogte boven zeeniveau.<br />
Bij 20 ºC op zeeniveau, bevat<br />
verzadigd water 9.1 mg zuurstof<br />
per liter. Bij een stijgende temperatuur<br />
daalt de concentratie; bij<br />
30 ºC is de zuurstofconcentratie 7.7<br />
mg/l. Net als in lucht neemt bij toenemende<br />
hoogte de zuurstofspanning,<br />
en daarmee ook de concentratie,<br />
in water af.<br />
De zuurstofmoleculen dringen het<br />
water binnen aan de oppervlakte,<br />
waarbij wind en golven zorgen<br />
voor menging. In het aquarium<br />
wordt hiervoor een bruiser gebruikt.<br />
Een tweede manier waarop<br />
142<br />
zuurstof in water komt is door afgifte door planten.<br />
Met behulp <strong>van</strong> energie uit licht produceren<br />
planten en algen koolhydraten waarbij<br />
zuurstof vrij komt. Dit proces heet fotosynthese.<br />
De fotosynthese stopt in het donker en dan verbruiken<br />
planten juist zuurstof, waardoor de<br />
zuurstofconcentratie in water met veel planten<br />
of algen ’s nachts zeer sterk kan dalen. Verder<br />
verdwijnt zuurstof uit het water doordat dit gas<br />
wordt verbruikt bij de ademhaling <strong>van</strong> dieren<br />
en voor een belangrijk deel door de afbraak <strong>van</strong><br />
dierlijk en plantaardig materiaal door micro-organismen.<br />
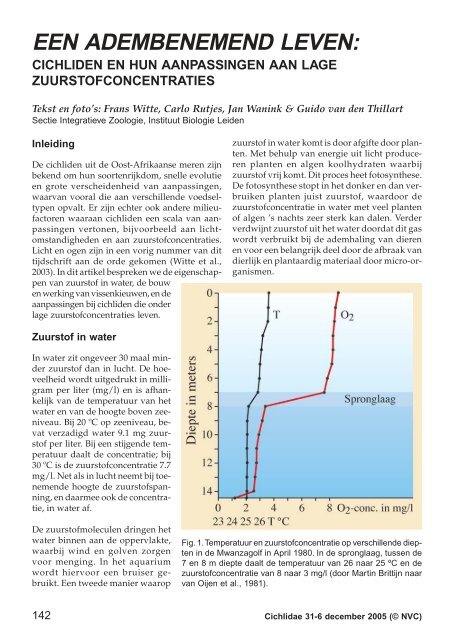
Fig. 1. Temperatuur en zuurstofconcentratie op verschillende diepten<br />
in de Mwanzagolf in April 1980. In de spronglaag, tussen de<br />
7 en 8 m diepte daalt de temperatuur <strong>van</strong> 26 naar 25 ºC en de<br />
zuurstofconcentratie <strong>van</strong> 8 naar 3 mg/l (door Martin Brittijn naar<br />
<strong>van</strong> Oijen et al., 1981).<br />
Cichlidae 31-6 december <strong>2005</strong> (© NVC)
De zuurstofconcentratie in water wordt lager<br />
naarmate men dieper komt, omdat zuurstof verbruikt<br />
wordt over de hele waterkolom, terwijl<br />
het in het water komt door diffusie aan de oppervlakte<br />
en door invloed <strong>van</strong> licht op planten<br />
die vooral in ondiep water voorkomen. In situaties<br />
met weinig wind en relatief hoge temperaturen<br />
aan de oppervlakte kan er zelfs een scherpe<br />
scheiding ontstaan tussen zuurstofrijk water in<br />
de bovenste laag en zuurstofarm water daaronder.<br />
Dit komt omdat het warme water lichter is<br />
en alleen kan mengen met de onderste koelere<br />
lagen bij voldoende wind en stroming. In deze<br />
gevallen noemt men het overgangsgebied,<br />
waarin de temperatuur snel afneemt, de spronglaag.<br />
In de Mwanzagolf <strong>van</strong> het Victoriameer kan<br />
een dergelijke spronglaag al op acht meter diepte<br />
voorkomen (Fig. 1; <strong>van</strong> Oijen et al., 1981).<br />
In heel diepe tropische meren zoals het Tanganjikameer<br />
(1470m diep) en het Malawimeer (700<br />
m diep) komt bene<strong>den</strong> de 200 m praktisch geen<br />
zuurstof meer voor (Fryer & Iles, 1972). Doordat<br />
de temperatuur afneemt met de diepte<br />
blijft het water permanent gelaagd. Op<br />
deze grote diepte heeft de wind geen invloed<br />
meer. In de gematigde en koude streken<br />
vindt tweemaal per jaar een volledige<br />
circulatie plaats. Water heeft zijn grootste<br />
dichtheid bij 4ºC. Zodra in de herfst de<br />
zuurstofrijke bovenlaag <strong>van</strong> het water 4ºC<br />
wordt, zakt deze naar omlaag. In de lente<br />
gebeurt hetzelfde nog een keer, waardoor<br />
in die streken wel zuurstof voorkomt op<br />
de bodem <strong>van</strong> diepe meren, zoals in het 1620 m<br />
diepe Baikalmeer (Kozhova & Izmest’eva, 1998).<br />
Ademhaling bij vissen<br />
Water is een minder geschikt medium voor ademhaling<br />
dan lucht. Behalve dat een bepaald volume<br />
water 30 maal minder zuurstof bevat dan een<br />
zelfde volume lucht, is de dichtheid <strong>van</strong> water<br />
1000 maal zo hoog als die <strong>van</strong> lucht. Door de hogere<br />
dichtheid <strong>van</strong> water is er meer energie nodig<br />
om het over een ademhalingsorgaan te pompen<br />
dan bij lucht het geval is. Toch zijn vissen goed<br />
aangepast aan ademhaling in het water. Ze kunnen<br />
tot 80% <strong>van</strong> de opgeloste hoeveelheid zuurstof<br />
uit het water halen. Ter vergelijking: de mens<br />
haalt maar 25% <strong>van</strong> de zuurstof uit de ingeademde<br />
lucht. Vissen danken hun efficiëntie aan<br />
de bouw <strong>van</strong> de kieuwen en de bloedcirculatie<br />
daarin, en aan het feit dat ze een continue waterstroom<br />
weten te genereren. In tegenstelling tot<br />
luchtademhaling met longen, waarbij de lucht<br />
heen en weer gepompt wordt, stroomt bij vissen<br />
het water <strong>van</strong> de mond, door de mondholte, over<br />
de kieuwen naar de kieuwopeningen. Deze<br />
stroom in één richting is <strong>van</strong> belang, want door<br />
de hoge dichtheid en viscositeit (= stroperigheid)<br />
<strong>van</strong> water zou omkeer <strong>van</strong> de stroomrichting veel<br />
energie kosten.<br />
Bij vissen zitten de kieuwen aan bogen die inwendig<br />
door skeletelementen gesteund wor<strong>den</strong>.<br />
Net als bij longen, is de bouw <strong>van</strong> kieuwen er<br />
op gericht het oppervlak, waarover de gasuitwisseling<br />
plaatsvindt, zo groot mogelijk te<br />
Kieuw <strong>van</strong> een cichlide. In de bovenste foto is het<br />
deel binnen het witte kader uitvergroot (ca. 60 x);<br />
de witte pijl geeft de richting aan <strong>van</strong> het aanzicht<br />
in de onderste foto (ca. 250 x). De richting <strong>van</strong> de<br />
water- (blauwe pijl) en bloedstroom (rode pijl) is<br />
aangegeven (foto’s Monique Welten; samenstelling<br />
Martin Brittijn).<br />
Cichlidae 31-6 december <strong>2005</strong> (© NVC) 143
Fig. 2. Het verschil tussen (a) het tegenstroom- en het (b) meestroomprincipe. Bij het een tegengestelde<br />
stroomrichting <strong>van</strong> bloed en water kan er over de hele lengte <strong>van</strong> de secundaire lamel zuurstof wor<strong>den</strong><br />
opgenomen. Daardoor wordt de zuurstofconcentratie in het bloed hoger dan bij een gelijke stroomrichting.<br />
Open pijlen geven de richting <strong>van</strong> de zuurstofoverdracht aan (door Martin Brittijn).<br />
maken. Beenvissen hebben gewoonlijk aan elke<br />
zijde <strong>van</strong> de kop vier kieuwbogen die kieuwen<br />
dragen, haaien en roggen hebben er 5 tot 7 aan elke<br />
zijde. Elke kieuwboog bevat twee rijen kieuwfilamenten.<br />
Een rij filamenten wordt ook wel een<br />
hemibranch (= halve kieuw) genoemd. Op de<br />
filamenten, die ook wel primaire lamellen genoemd<br />
wor<strong>den</strong>, staan secundaire lamellen. Dit zijn<br />
grotendeels holle ruimten, waardoor het bloed<br />
stroomt en die opengehou<strong>den</strong> wor<strong>den</strong> door pilaarcellen.<br />
Zuurstofopname vindt plaats via het oppervlak<br />
<strong>van</strong> de secundaire lamellen. Bij haplochromine<br />
cichli<strong>den</strong> varieert het totale aantal filamenten op<br />
de acht kieuwbogen, <strong>van</strong> linker en rechterzijde samen,<br />
afhankelijk <strong>van</strong> de soort en de grootte <strong>van</strong> de<br />
vis, <strong>van</strong> 800 tot 8000. De gemiddelde lengte <strong>van</strong> de<br />
filamenten bedraagt 1 tot 5 mm (Galis en Barel,<br />
1980). De lengte <strong>van</strong> alle kieuwfilamenten samen<br />
is dus al gauw enkele meters. Het aantal secundaire<br />
lamellen per mm filament bedraagt 40 tot 80<br />
(aan twee zij<strong>den</strong>) en het totale aantal kan oplopen<br />
tot enkele honderdduizen<strong>den</strong>.<br />
De waterstroom door de mond- en kieuwholte<br />
wordt vooral opgewekt door het op en neer bewegen<br />
<strong>van</strong> de mondbodem en het naar buiten<br />
en binnen draaien <strong>van</strong> de wangplaten. Sommige<br />
snelle zwemmers zoals makrelen en tonijnen<br />
hebben zoveel zuurstof nodig dat de pompbeweging<br />
met de mond niet voldoende zou opleveren.<br />
Deze soorten laten het water over de<br />
kieuwen stromen door te zwemmen met een<br />
geopende bek. Dit wordt met een Engelse term<br />
“ram” ventilatie genoemd.<br />
144<br />
Vissen hebben in hun kieuwen een zogenaamd<br />
tegenstroomprincipe. Dat wil zeggen dat bij de<br />
secundaire lamellen bloed en water in tegengestelde<br />
richting stromen. Hierdoor komt het<br />
zuurstofarme bloed dat de lamellen instroomt<br />
in aanraking met water dat al een deel <strong>van</strong> de<br />
zuurstof heeft afgestaan. Het bloed dat de lamellen<br />
verlaat en bijna verzadigd is, komt in contact<br />
met het zuurstofrijkste water. Door dit tegenstroomprincipe<br />
zit er altijd meer zuurstof in het<br />
water dan in het bloed, waardoor over het hele<br />
traject zuurstofopname kan plaatsvin<strong>den</strong> (Fig. 2).<br />
Als het bloed en het water in dezelfde richting<br />
zou<strong>den</strong> stromen, zou er aan<strong>van</strong>kelijk een groot<br />
verschil in zuurstofconcentratie bestaan. Door<br />
het afstaan <strong>van</strong> zuurstof zou het water uiteindelijk<br />
dezelfde concentratie krijgen als het bloed<br />
en zou er geen overdracht <strong>van</strong> zuurstof meer<br />
plaatsvin<strong>den</strong> (Fig. 2).<br />
Als de zuurstofconcentratie in water afneemt<br />
moet de waterstroom over de kieuwen toenemen<br />
om de zuurstofopname gelijk te hou<strong>den</strong>. Als er<br />
bijvoorbeeld 10 keer minder zuurstof in het water<br />
zit dan normaal, moet de waterstroom over<br />
de kieuwen 10 keer zo groot wor<strong>den</strong>. Dit kan<br />
bereikt wor<strong>den</strong> door sneller te pompen met een<br />
groter slagvolume.<br />
Uitscheiding <strong>van</strong> koolzuur, ammoniak en zouten<br />
vindt via de kieuwen plaats. De zoutbalans<br />
(opname en uitscheiding <strong>van</strong> chlori<strong>den</strong>) <strong>van</strong> de<br />
vis wordt geregeld door speciale chloridecellen<br />
aan de basis de secundaire lamellen.<br />
Cichlidae 31-6 december <strong>2005</strong> (© NVC)
De effecten <strong>van</strong> lage zuurstofconcentraties<br />
op vissen<br />
Door verschillende oorzaken kan de zuurstofspanning<br />
in het milieu <strong>van</strong> vissen lager wor<strong>den</strong><br />
dan normaal. We spreken dan <strong>van</strong> hypoxie. Opgemerkt<br />
moet wor<strong>den</strong> dat dit een relatief begrip<br />
is. Voor vissen die normaal leven in een omgeving<br />
met 7 mg zuurstof per liter, is bijvoorbeeld<br />
3 mg/l hypoxisch, terwijl voor vissen die in een<br />
milieu leven met 3 mg/l, een zuurstofconcentratie<br />
<strong>van</strong> 1 mg/l pas hypoxisch kan zijn. Door<br />
toenemende voedselrijkdom (eutrofiëring) en<br />
andere vervuiling <strong>van</strong> water neemt de hoeveelheid<br />
zuurstof wereldwijd in veel meren en andere<br />
wateren af. Bij eutrofiëring ontstaat algenbloei.<br />
Wanneer deze algen dood gaan, kost de<br />
afbraak er<strong>van</strong> veel zuurstof. Dit laatste kan lei<strong>den</strong><br />
tot zelfs volledige zuurstofloosheid (anoxie).<br />
Op korte termijn kunnen vissen op twee manieren<br />
reageren op hypoxie wanneer een aanpassing<br />
door een snellere ademhaling niet meer<br />
mogelijk is. Ze kunnen inactief wor<strong>den</strong>, waardoor<br />
ze energie sparen, of ze kunnen wegvluchten,<br />
wat extra energie kost. Ieder <strong>van</strong> deze<br />
reacties heeft zijn eigen voordeel. In het eerste<br />
geval kan de vis het langer uithou<strong>den</strong> bij de lage<br />
zuurstofspanning, in het tweede geval kan hij<br />
in een gunstigere omgeving terechtkomen. Veel<br />
vissoorten vluchten naar de oppervlakte waar<br />
ze water, vermengd met lucht, ophappen. De<br />
reactie verschilt per vissoort en per situatie.<br />
kleiner bij een anaërobe stofwisseling. Afhankelijk<br />
<strong>van</strong> de soort, kunnen vissen deze situatie enkele<br />
minuten tot enkele dagen overleven. De<br />
Zuid-Amerikaanse pauwoogcichlide, Astronotus<br />
ocellatus, kan langer dan 16 uur overleven bij een<br />
zuurstofconcentratie <strong>van</strong> 0,4 mg/l in water <strong>van</strong><br />
28 ºC en zelfs vier uur bij complete zuurstofloosheid<br />
(Muusze et al., 1998). In het milieu <strong>van</strong><br />
deze soort in het Amazonebekken komen lage<br />
zuurstofconcentraties regelmatig voor.<br />
Tolerantie jegens kortdurende lage<br />
zuurstofconcentraties bij cichli<strong>den</strong><br />
Er zijn verschillen gevon<strong>den</strong> in gedrag, eigenschappen<br />
<strong>van</strong> het bloed, stofwisselingssnelheid<br />
en kieuwvorm tussen cichli<strong>den</strong> die in verschillende<br />
omgevingen leven. Soorten die voorkomen<br />
in moerassen <strong>van</strong> de satellietmeren <strong>van</strong> het<br />
Victoriameer hebben meer rode bloedlichaampjes,<br />
een hoger hemoglobine-gehalte, een groter<br />
kieuwoppervlak, en zijn toleranter jegens<br />
hypoxie dan soorten uit het open water in deze<br />
meren (Chapman et al., 2002). Verder hebben<br />
cichli<strong>den</strong> meerdere typen hemoglobine (Verheyen<br />
et al., 1986) die bij verschillende temperaturen<br />
en zuurstofspanningen functioneel zijn.<br />
De reactie op kortdurende hypoxie werd onderzocht<br />
bij negen cichli<strong>den</strong>soorten uit het Victoriameer<br />
(zeven haplochromine- en twee tilapiine<br />
soorten) en drie soorten uit het Tanganjikameer<br />
(Cyprichromis leptosoma, Tropheus moorii en<br />
Neolamprologus tretocephalus; Chapman et al.,<br />
Het belangrijkste fysiologische effect <strong>van</strong> 1995). Onderzocht werd hoe goed de soorten aan<br />
hypoxie is een zeer sterke<br />
afname <strong>van</strong> het vermogen Aloys Peter tij<strong>den</strong>s het meten <strong>van</strong> de zuurstofconcentratie in de Mwanzagolf.<br />
om zuurstof op te nemen,<br />
waardoor de vis minder<br />
energie beschikbaar heeft<br />
voor allerlei activiteiten.<br />
Het evenwicht tussen energieverbruik<br />
en zuurstofopname<br />
bepaalt de zogeheten<br />
anaërobe drempel.<br />
Dit is de grens tussen de<br />
aërobe (= met zuurstof)<br />
stofwisseling en anaërobe<br />
(= zonder zuurstof) stofwisseling.<br />
Het is een kritisch<br />
punt, want de overlevingskansen<br />
wor<strong>den</strong> snel<br />
Cichlidae 31-6 december <strong>2005</strong> (© NVC) 145
De rotsbewonende Haplochromis (Neochromis) rufocaudalis is minder<br />
tolerant voor lage zuurstofconcentraties dan haplochrominen uit andere<br />
milieus <strong>van</strong> het Victoriameer (zie tekst).<br />
Astatoreochromis alluaudi , een soort die goed tegen lage zuurstofconcentraties<br />
kan. foto: Peter Snelderwaard<br />
Trematocara unimaculatum, een hypoxie-tolerante vis uit het Tanganjikameer.<br />
foto: Ad Konings<br />
de oppervlakte kon<strong>den</strong> ademen tij<strong>den</strong>s hypoxie<br />
en hoe snel ze hun evenwicht verloren (als ge-<br />
146<br />
volg <strong>van</strong> verdoving) als ze<br />
plotseling overgeplaatst<br />
wer<strong>den</strong> <strong>van</strong> een hoge naar<br />
een lage zuurstofconcentratie.<br />
De haplochromine<br />
soorten uit het Victoriameer<br />
bleken toleranter jegens<br />
hypoxie dan ecologisch<br />
vergelijkbare soorten<br />
uit het Tanganjikameer<br />
(Chapman et al., 1995).<br />
Van de Victoria-cichli<strong>den</strong><br />
verloor alleen Haplochromis<br />
(Neochromis) rufocaudalis,<br />
zijn evenwicht binnen 30<br />
minuten bij plotselinge<br />
hypoxie (0,2 mg/l). Deze<br />
algengrazer leeft in zuurstofrijk<br />
water bij de rotsen.<br />
Van de Tanganjika-cichli<strong>den</strong><br />
verloren alle soorten<br />
hun evenwicht bij plotselinge<br />
hypoxie, en dat gebeurde<br />
bij deze soorten<br />
veel sneller dan bij H.<br />
rufocaudalis.<br />
De slakkenkraker Astatorechromis<br />
alluaudi die behalve<br />
in het Victoriameer ook<br />
in satellietmeren, stroompjes<br />
en moerassen voorkomt,<br />
bleek goed aan de<br />
oppervlakte te kunnen<br />
ademen. Wanneer de vissen<br />
niet bij de oppervlakte<br />
kon<strong>den</strong> komen bleven alleen<br />
de tilapiasoorten Oreochromis<br />
esculentus en O.<br />
niloticus overeind bij plotselinge<br />
hypoxie. Deze twee<br />
soorten, waar<strong>van</strong> de laatstgenoemde<br />
geïntroduceerd<br />
werd in het Victoriameer,<br />
kon<strong>den</strong> dus het best tegen<br />
extreem lage zuurstofconcentraties<br />
(minder dan 0,2<br />
mg/l).<br />
Uit het Tanganjikameer<br />
zijn ook soorten bekend die goed tegen hypoxie<br />
kunnen, bijvoorbeeld Trematocara unimaculatum<br />
Cichlidae 31-6 december <strong>2005</strong> (© NVC)
Fig. 3. Diepte waarop Haplochromis (Gaurochromis) hiatus en H. (G.) iris in de Mwanzagolf leef<strong>den</strong>. In<br />
de omgeving <strong>van</strong> H. iris was de zuurstofconcentratie in sommige perio<strong>den</strong> lager dan in de omgeving <strong>van</strong><br />
H. hiatus (zie Fig. 1). De kieuwen <strong>van</strong> H. iris waren groter dan die <strong>van</strong> H. hiatus door onder andere een<br />
groter aantal filamenten (door Martin Brittijn, naar Hoogerhoud et al., 1983).<br />
en Hemibates stenosoma die ge<strong>van</strong>gen wer<strong>den</strong> op<br />
diepten <strong>van</strong> 205 m, waar de zuurstofconcentratie<br />
minder dan 1 mg/l was (Coulter, 1967).<br />
Uit onderzoek <strong>van</strong> Galis en Barel (1980) bleek<br />
dat soorten uit het Victoria-, George-, Malawien<br />
Tanganjikameer met een verschillende levenswijze,<br />
bijvoorbeeld snelle en langzame zwemmers,<br />
verschillen in de vorm en grootte <strong>van</strong> de<br />
kieuwen. De twee nauw verwante insectenetende<br />
soorten Haplochromis (Gaurochromis) hiatus<br />
en H. (G.) iris uit het Victoriameer, die sinds de<br />
Nijlbaarsexplosie niet meer ge<strong>van</strong>gen wer<strong>den</strong>,<br />
vorm<strong>den</strong> een mooi voorbeeld. De laatstgenoemde<br />
soort had een kieuwoppervlak dat 60%<br />
groter was dan dat <strong>van</strong> de eerstgenoemde (Fig. 3;<br />
Hoogerhoud et al.,1983). De verklaring voor dit<br />
verschil is dat H. hiatus leefde in water <strong>van</strong> 3 tot<br />
9 m diep en H. iris in water <strong>van</strong> 8 tot 15 m. In dit<br />
diepere water wer<strong>den</strong> in het regenseizoen regelmatig<br />
lage zuurstofconcentraties (minder dan 3<br />
mg/l) gemeten na het ontstaan <strong>van</strong> een spronglaag<br />
(Fig. 1). Verschillen in kieuwoppervlak zijn<br />
ook gevon<strong>den</strong> tussen populaties <strong>van</strong> dezelfde<br />
soort, bijvoorbeeld Pseudocrenilabrus multicolor<br />
victoriae uit het Kayanjameer met een zuurstofgehalte<br />
<strong>van</strong> 6,1 mg/l en uit de moerassen <strong>van</strong><br />
het Manywameer met een gehalte <strong>van</strong> 0,4 mg/l<br />
(Chapman et al., 2000). Het kieuwoppervlak <strong>van</strong><br />
de laatstgenoemde populatie is 41% groter dan<br />
dat <strong>van</strong> de eerstgenoemde.<br />
Het effect <strong>van</strong> langdurig lage zuurstofconcentraties<br />
in het Victoriameer<br />
Door de toegenomen algenbloei in de afgelopen<br />
30 jaar is de zuurstofconcentratie in de Mwanzagolf<br />
<strong>van</strong> het Victoriameer sterk afgenomen<br />
(<strong>Wanink</strong> et al., 2001). De algenbloei is onder andere<br />
het gevolg <strong>van</strong> verrijking <strong>van</strong> het water met<br />
voedingstoffen door de toenemende ontbossing<br />
en landbouw. Maar ook het verdwijnen <strong>van</strong> de<br />
algenetende Haplochromis-soorten door de geïntroduceerde<br />
Nijlbaars draagt waarschijnlijk bij<br />
aan de algenbloei (Goldschmidt et al., 1993). De<br />
zoöplanktoneter H. (Yssichromis) pyrrhocephalus,<br />
is op het ogenblik een <strong>van</strong> de weinige Haplochromissoorten<br />
die zich hersteld hebben <strong>van</strong> de Nijlbaars<br />
predatie (<strong>Witte</strong> & <strong>Wanink</strong> 2000). Het kieuwoppervlak<br />
<strong>van</strong> deze soort blijkt binnen 20 jaar<br />
Cichlidae 31-6 december <strong>2005</strong> (© NVC) 147
Hypoxieopstelling in het laboratorium in Lei<strong>den</strong>.<br />
Aquarium met vissen bij een zuurstofconcentratie <strong>van</strong> 0,8 mg/l.<br />
meer dan 60% in grootte te zijn toegenomen (M.<br />
Welten, ongepubliceerde gegevens). Deze toename<br />
werd gerealiseerd door verlenging <strong>van</strong> de<br />
filamenten en vergroting <strong>van</strong> het oppervlak <strong>van</strong><br />
de secundaire lamellen.<br />
Om nader te onderzoeken hoe deze veranderingen<br />
tot stand komen, hebben we in het laboratorium<br />
in Lei<strong>den</strong> experimenten gedaan waarbij<br />
cichli<strong>den</strong>soorten uit de Mwanzagolf onder permanente<br />
hypoxie zijn opgekweekt. Jonge vissen<br />
<strong>van</strong> 1,5 tot 2 cm groot wer<strong>den</strong> in aquaria gezet<br />
waar<strong>van</strong> de zuurstofspanning in de loop <strong>van</strong> en-<br />
148<br />
kele weken teruggebracht<br />
werd <strong>van</strong> een normale zuurstofconcentratie<br />
(7 mg/l),<br />
normoxie genoemd, tot extreme<br />
hypoxie (0,8 mg/l).<br />
Alle soorten bleken deze<br />
overgang goed te overleven.<br />
De hypoxie-vissen groei<strong>den</strong><br />
bijna even snel als nestgenoten<br />
die onder normoxie<br />
opgekweekt wer<strong>den</strong>. Enkele<br />
soorten kwamen zelfs tot<br />
broe<strong>den</strong>. Na anderhalf jaar,<br />
de vissen waren inmiddels<br />
volwassen gewor<strong>den</strong>, wer<strong>den</strong><br />
de kieuwen onderzocht.<br />
Net als in het Victoriameer<br />
bleken de hypoxie-vissen<br />
een kieuwoppervlak te hebben<br />
dat 60-80% groter was<br />
dan dat <strong>van</strong> de normoxievissen<br />
(C. <strong>Rutjes</strong>, ongepubliceerde<br />
gegevens). Hieruit<br />
blijkt dat de kieuwen <strong>van</strong><br />
deze Haplochromis-soorten<br />
uitermate plastisch zijn en<br />
dat de vissen zich door “training”<br />
kunnen aanpassen aan<br />
lage zuurstofspanningen. De<br />
uiterlijke kenmerken (het<br />
fenotype) <strong>van</strong> een individu<br />
verander<strong>den</strong> dus onder invloed<br />
<strong>van</strong> hypoxie; de genen<br />
<strong>van</strong> het individu verander<strong>den</strong><br />
echter niet, want die zijn<br />
bij de bevruchting al vastgelegd.<br />
Het verschijnsel dat één<br />
genotype onder invloed <strong>van</strong><br />
het milieu verschillende<br />
fenotypen produceert noemen we fenotypische<br />
plasticiteit. Het is een eigenschap die uitermate<br />
handig is voor planten en dieren die in wisselende<br />
milieus leven. Het vermogen met fenotypische<br />
plasticiteit te reageren op variatie in het<br />
milieu, is een genetisch bepaalde eigenschap die<br />
vooral voorkomt bij organismen die in onstabiele<br />
milieus leven. Als een omgeving tijdelijk verandert,<br />
bijvoorbeeld hypoxisch wordt, dan kunnen<br />
organismen dit overleven dankzij deze fenotypische<br />
plasticiteit. Blijft de hypoxische situatie<br />
generaties lang voortbestaan dan kan het genotype<br />
veranderen door natuurlijke selectie, wat<br />
Cichlidae 31-6 december <strong>2005</strong> (© NVC)
Fig. 4. Een theoretisch voorbeeld <strong>van</strong> de negatieve relatie<br />
tussen de grootte <strong>van</strong> de kieuwen en de omliggende<br />
spieren (rood gekleurd) in de kop <strong>van</strong> een cichlide. sh =<br />
sternohyoideus spier (door Martin Brittijn, naar een figuur<br />
<strong>van</strong> Marnix de Zeeuw).<br />
meestal leidt tot betere aanpassingen dan die<br />
welke alleen door fenotypische plasticiteit verkregen<br />
wor<strong>den</strong>. In hoeverre de veranderingen<br />
in de kieuwen <strong>van</strong> H. pyrrhocephalus uit het<br />
Victoriameer berusten op fenotypische plasticiteit<br />
of het gevolg zijn <strong>van</strong> natuurlijke selectie,<br />
weten we nog niet. We vermoe<strong>den</strong> echter dat het<br />
om een combinatie <strong>van</strong> beide effecten gaat.<br />
Interessant is de vraag, hoe grotere kieuwen in<br />
de kop geplaatst kunnen wor<strong>den</strong>. De structuren<br />
in de kop <strong>van</strong> een cichlide zitten dicht op elkaar<br />
gepakt en er is dus weinig vrije ruimte beschikbaar.<br />
Er zijn ten minste twee mogelijkhe<strong>den</strong> om<br />
dit ruimteprobleem op te lossen. De ene is een<br />
grotere kop, maar dit kan nadelige effecten heb-<br />
ben op de stroomlijn <strong>van</strong> de vis. De andere<br />
mogelijkheid is het verkleinen <strong>van</strong><br />
structuren die aan de kieuwen grenzen<br />
(Fig. 4). Hierdoor kan de functie <strong>van</strong> deze<br />
structuren echter nadelig beïnvloed wor<strong>den</strong>.<br />
Een afname <strong>van</strong> bijvoorbeeld de dikte<br />
<strong>van</strong> de sternohyoideus spier, die voor de<br />
bekopening zorgt bij het opzuigen <strong>van</strong><br />
prooien, zou kunnen betekenen dat de vis<br />
bepaalde prooien minder goed kan <strong>van</strong>gen.<br />
Het verschijnsel waarbij door verbetering<br />
<strong>van</strong> de ene functie een andere wordt<br />
aangetast wordt in het Engels een “tradeoff”<br />
genoemd.<br />
In het verle<strong>den</strong> is diverse malen gesuggereerd<br />
dat hypoxie in het Victoriameer een<br />
belangrijker oorzaak zou zijn voor het verdwijnen<br />
<strong>van</strong> de haplochromine cichli<strong>den</strong><br />
gedurende de afgelopen decennia dan<br />
predatie door de Nijlbaars. Ons onderzoek<br />
wijst er op dat de Victoriacichli<strong>den</strong> die <strong>van</strong><br />
jongs af aan onder hypoxie opgroeien zich<br />
redelijk aan zulke omstandighe<strong>den</strong> kunnen<br />
aanpassen. Wat we nog moeten onderzoeken<br />
is hoe die veranderingen tot stand<br />
komen (fenotypische plasticiteit, natuurlijke<br />
selectie of een combinatie <strong>van</strong> beide)<br />
en wat de effecten <strong>van</strong> de “trade-offs” zijn.<br />
Wat we ook nog niet weten is of de ontwikkeling<br />
<strong>van</strong> embryo’s onder hypoxie<br />
goed verloopt. De enige soort die in onze<br />
proefopstelling bij 0,8 mg/l zuurstof haar<br />
jongen succesvol kon groot brengen was<br />
A. alluaudi. Deze soort komt ook onder natuurlijke<br />
omstandighe<strong>den</strong> in milieus voor<br />
met lage zuurstofconcentraties. Als hypoxie voor<br />
de embryo’s <strong>van</strong> andere soorten dodelijk is, zal<br />
het effect er<strong>van</strong> op de soortenrijkdom veel groter<br />
kunnen zijn dan de bovenbeschreven experimenten<br />
doen vermoe<strong>den</strong>.<br />
In de komende tijd zullen wij ons onderzoek<br />
richten op de bovengestelde vragen. Behalve het<br />
begrijpen <strong>van</strong> de effecten <strong>van</strong> hypoxie op de diversiteit<br />
<strong>van</strong> cichli<strong>den</strong>, hopen we door dit onderzoek<br />
ook inzicht te verkrijgen in mechanismen<br />
die een rol spelen bij de snelle evolutie en<br />
adaptieve radiatie <strong>van</strong> de cichli<strong>den</strong> uit de grote<br />
Afrikaanse meren.<br />
Cichlidae 31-6 december <strong>2005</strong> (© NVC) 149
Literatuur:<br />
Anker, G. Ch., 1989. Neth. J. Zool. 39: 1-40.<br />
Barel, C.D.N., F. <strong>Witte</strong> & M.J.P. <strong>van</strong> Oijen, 1976.<br />
Neth. J. Zool. 26: 163-265.<br />
Barel, C.D.N., M.J.P. <strong>van</strong> Oijen, F. <strong>Witte</strong> & E.L.M.<br />
<strong>Witte</strong>-Maas, 1977. Neth. J. Zool. 27: 333-389.<br />
Bouton, N., N. <strong>van</strong> Os & F. <strong>Witte</strong>, 1998. J. Fish. Biol.<br />
53 (A): 118-127.<br />
Chapman, L.J., L.S. Kaufman, C.A. Chapman & F.E.<br />
McKenzie, 1995. Conserv. Biol. 9: 1274-1288.<br />
Chapman, L. J., C.A. Chapman, F.G. Nordlie & A.E.<br />
Rosenberger, 2002. Comp. Biochem. Physiol. A 133:<br />
421-437.<br />
Coulter, G.W., 1967. Nature 215: 317-318.<br />
Eschmeyer, W.N., C.J. Ferraris & M.D. Hoang, 1998.<br />
Catalogue of Fishes, 3 vols. Cal. Acad. Sci., San<br />
Francisco, 2905 pp.<br />
Fryer, G., 1960. E. Afr. J. Agric. 25: 267-270.<br />
Fryer, G. & T.D. Iles, 1972. The Cichlid Fishes of the<br />
Great Lakes of Africa. Oliver & Boyd, Edinburgh.<br />
Galis, F. & C.D.N. Barel, 1980. Neth. J. Zool. 30:<br />
392-430.<br />
Goldschmidt, T., 1994. Darwins hofvijver. Prometheus<br />
Amsterdam.<br />
Goldschmidt, T., F. <strong>Witte</strong> & J. de Visser, 1990. Oikos<br />
58: 343-355.<br />
Goldschmidt, T., F. <strong>Witte</strong> & J. <strong>Wanink</strong>, 1993. Conserv.<br />
Biol. 7: 686-700.<br />
Goudswaard, P.C., F. <strong>Witte</strong> & E.F.B. Katunzi, 2002.<br />
J. Fish Biol. 60: 838-856.<br />
Greenwood, P.H., 1974. Bull. Br. Mus. nat. Hist.<br />
(Zool.) Suppl. 6.<br />
164<br />
De treiler Mdiria, voorheen Texel 34,<br />
waarmee veel haplochrominen voor<br />
het onderzoek <strong>van</strong> HEST ge<strong>van</strong>gen<br />
wer<strong>den</strong>.<br />
Greenwood, P.H., 1981. The haplochromine fishes<br />
of the East African Lakes. Kraus-Thomson<br />
Organization GmbH, München.<br />
Hoogerhoud, R.J.C., F. <strong>Witte</strong> & C.D.N. Barel, 1983.<br />
Neth. J. Zool. 33: 283-305.<br />
Katunzi, E.F.B., J. Zoutendijk, T. Goldschmidt, J.H.<br />
<strong>Wanink</strong> & F. <strong>Witte</strong>, 2003. Ecol. Freshw. Fish 12: 237-<br />
240.<br />
Kozhova O.M. & L.R. Izmest’eva, 1998. Lake Baikal.<br />
Backhuys Publishers, Lei<strong>den</strong>.<br />
Muusze B., J. Marcon, G. <strong>van</strong> <strong>den</strong> <strong>Thillart</strong> & V.<br />
Almeida-Val, 1998. Comp. Biochem. Physiol. 120A:<br />
151-156.<br />
Oijen, M.J.P. <strong>van</strong>, F. <strong>Witte</strong> & E.L.M. <strong>Witte</strong>-Maas,<br />
1981. Neth. J. Zool. 31: 149-174.<br />
Oijen, M.J.P. <strong>van</strong>, 1991. Zool. Verh. Lei<strong>den</strong> 272: 1-<br />
95.<br />
Oijen, M.J.P. <strong>van</strong>, & F. <strong>Witte</strong>, 1996. Zool. Verh. Lei<strong>den</strong><br />
302: 1-56.<br />
Seehausen, O. & J.J.M. <strong>van</strong> Alphen, 1998. Behav.<br />
Ecol. Sociobiol. 42: 1-8.<br />
Seehausen, O., J.J.M. <strong>van</strong><br />
Alphen & F. <strong>Witte</strong>, 1997. Science<br />
277: 1808-1811.<br />
Seehausen, O., E. Lippitsch, N.<br />
Bouton & H. Zwennes, 1998.<br />
Ichthyol. Expl. Freshw. 9: 129-<br />
228.<br />
Seehausen, O., 1996. Lake<br />
Victoria Rock Cichlids. Verduijn<br />
Cichlids, Zevenhuizen.<br />
Verheyen, E., R. Blust & W.<br />
Decleir, 1986. Comp. Biochem.<br />
Physiol. 84A: 315-318.<br />
<strong>Wanink</strong>, J.H., J.J. Kashindye,<br />
P.C. Goudswaard & F. <strong>Witte</strong>,<br />
2001. Freshw. Biol. 46: 75-85.<br />
<strong>Wanink</strong>, J.H., 1999. Hydrobiologia<br />
407: 183-189.<br />
<strong>Witte</strong>, F., 1981. Neth. J. Zool. 31:<br />
175-202.<br />
<strong>Witte</strong>, F., & W.L.T. <strong>van</strong> Densen<br />
(eds), 1995. Fish Stocks and<br />
Fisheries of Lake Victoria.<br />
Samara Publishing Ltd. Cardigan, Dyfed.<br />
<strong>Witte</strong>, F. & M.J.P. <strong>van</strong> Oijen, 1990. Zool. Verh. Lei<strong>den</strong><br />
262: 1-47.<br />
<strong>Witte</strong>, F. & J.H. <strong>Wanink</strong>, 2000. Cichlidae 26: 77-84.<br />
<strong>Witte</strong>, F., H. <strong>van</strong> der Meer & C.D.N. Barel, 2003.<br />
Cichlidae 29: 104-114.<br />
<strong>Witte</strong>, F., T. Goldschmidt, J. <strong>Wanink</strong>, M. <strong>van</strong> Oijen,<br />
K, Goudswaard, E. <strong>Witte</strong>-Maas & N. Bouton, 1992.<br />
Env. Biol. Fish. 34: 1-28.<br />
Cichlidae 31-6 december <strong>2005</strong> (© NVC)














