

A fast, robust and tunable synthetic gene oscillator - The BioCircuits ...
A fast, robust and tunable synthetic gene oscillator - The BioCircuits ...
A fast, robust and tunable synthetic gene oscillator - The BioCircuits ...

You also want an ePaper? Increase the reach of your titles
YUMPU automatically turns print PDFs into web optimized ePapers that Google loves.
doi: 10.1038/nature07389 SUPPLEMENTARY INFORMATION<br />
Period (min)<br />
25<br />
20<br />
15<br />
10<br />
5<br />
k l = 1000*k l, original (deterministic)<br />
k l = k l, original (deterministic)<br />
k l = 0 (stochastic)<br />
0<br />
0 0.2 0.4 0.6 0.8 1<br />
IPTG (mM)<br />
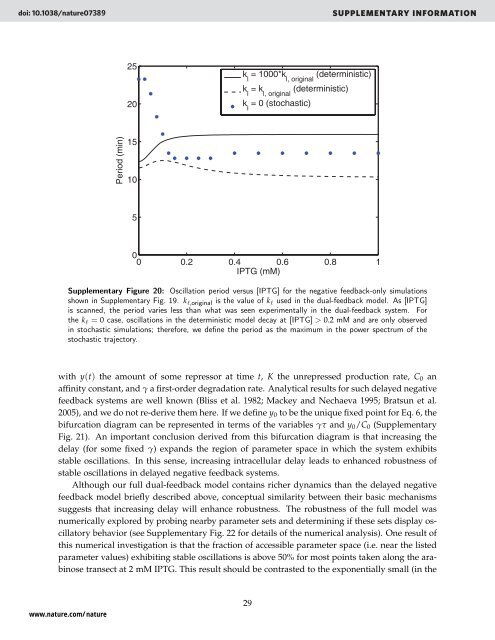
Supplementary Figure 20: Oscillation period versus [IPTG] for the negative feedback-only simulations<br />
shown in Supplementary Fig. 19. k ℓ,original is the value of k ℓ used in the dual-feedback model. As [IPTG]<br />
is scanned, the period varies less than what was seen experimentally in the dual-feedback system. For<br />
the k ℓ = 0 case, oscillations in the deterministic model decay at [IPTG] > 0.2 mM <strong>and</strong> are only observed<br />
in stochastic simulations; therefore, we define the period as the maximum in the power spectrum of the<br />
stochastic trajectory.<br />
with y(t) the amount of some repressor at time t, K the unrepressed production rate, C0 an<br />
affinity constant, <strong>and</strong> γ a first-order degradation rate. Analytical results for such delayed negative<br />
feedback systems are well known (Bliss et al. 1982; Mackey <strong>and</strong> Nechaeva 1995; Bratsun et al.<br />
2005), <strong>and</strong> we do not re-derive them here. If we define y0 to be the unique fixed point for Eq. 6, the<br />
bifurcation diagram can be represented in terms of the variables γτ <strong>and</strong> y0/C0 (Supplementary<br />
Fig. 21). An important conclusion derived from this bifurcation diagram is that increasing the<br />
delay (for some fixed γ) exp<strong>and</strong>s the region of parameter space in which the system exhibits<br />
stable oscillations. In this sense, increasing intracellular delay leads to enhanced <strong>robust</strong>ness of<br />
stable oscillations in delayed negative feedback systems.<br />
Although our full dual-feedback model contains richer dynamics than the delayed negative<br />
feedback model briefly described above, conceptual similarity between their basic mechanisms<br />
suggests that increasing delay will enhance <strong>robust</strong>ness. <strong>The</strong> <strong>robust</strong>ness of the full model was<br />
numerically explored by probing nearby parameter sets <strong>and</strong> determining if these sets display <strong>oscillator</strong>y<br />
behavior (see Supplementary Fig. 22 for details of the numerical analysis). One result of<br />
this numerical investigation is that the fraction of accessible parameter space (i.e. near the listed<br />
parameter values) exhibiting stable oscillations is above 50% for most points taken along the arabinose<br />
transect at 2 mM IPTG. This result should be contrasted to the exponentially small (in the<br />
www.nature.com/nature<br />
29




